Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2013) Volume 3, Issue 2
Keywords: Fungus, Aspergillus niger, Heterogeneity, Sporulation,flbA, Regulator of G protein signaling, Transcriptome
Aspergillus species are among the most abundant fungi worldwide. They feed on a large variety of organic substrates, in particular, on plant material [1]. To this end, enzymes are secreted that degrade the organic polymers within the substrate into small molecules that can be taken up as nutrients. The capacity of Aspergillus species to secrete proteins is enormous. For instance, some strains of Aspergillus niger secrete more than 30 grams per liter of glucoamylase [2]. This and the fact that a variety of secreted enzymes of Aspergilli are used in the industry or as pharmaceutical proteinsmakes these fungi important cell factories.
Aspergilli form colonies that consist of hyphae that grow at their tips and that branch subapically [3]. Growth and protein secretion mainly occur at the periphery of the colony when they are grown on an agar medium [4,5]. Hyphae at the periphery of the colonyare exposed to unexplored organic material, whereas hyphae in theintermediate and central zones are confronted with a (partly) utilized substrate. The composition of the substrate explains about 50% of the variation in gene expression between different zones of the colonies [6]. The other half of the variation is caused by differentiation processes in the vegetative mycelium.
Recently, it was shown that zones of the colony that have the capacity to sporulate hardly secrete proteins [7]. This suggested that secretion by vegetative hyphae is repressed by the sporulation process. Indeed, a ΔflbA strain that does not sporulate not only secretes proteins at the colony periphery, but also within its central zones. This is accompanied by a more complex secretome, a reduced width of cell walls, and growth throughout the mycelium [8]. Gene flbA encodes an RGS domain protein that stimulates the intrinsic GTPase activity of the Gα-subunit FadA [9]. This Gα-subunit is part of a heterotrimeric G-protein complex that also consists of the Gβ- and Gγ-subunits SfaD and GpgA, respectively (Figure 1). The activated Gα-subunit and the Gβ-Gγ dimer both stimulate vegetative growth. FlbA converts the active Gα-subunit-GTP into its inactive GDP bound stage, thereby repressing vegetative growth and promoting asexual development [10-12]. Taken together, these data indicate that FlbA stimulates sporulation and at the same time, represses vegetative growth and secretion by the vegetative mycelium. Moreover, it would promote synthesis of cell wall polymers, resulting in an increased width of the cell wall. The underlying molecular mechanisms are not yet known.

Figure 1: Model of the role of FlbA in asexual sporulation of Aspergillus. FlbA is an RGS protein regulating signaling of the Gα-subunit FadA. GTP-bound FadA stimulates vegetative growth. FlbA enhances the intrinsic activity of GTP hydrolysis, converting FadA-GTP to the inactive hetrotrimeric FadA-SfaD-GpgA GDP-bound G-protein complex. FlbA also inhibits fluG and activates brlA. The latter may be a direct effect or the result of inactivation of FadA. Stippled lines represent regulation of FlbA in A. niger based on the transcriptional analysis described this study. Adapted from [3].
Here, the impact of inactivation of flbA on spatial gene expression in the A. niger colony was assessed. A total of 1152 genes had changed their expression ≥ 2 fold when RNA profiles of wild-type and ΔflbA colonies were compared. This set includes genes involved in reproduction and cell wall synthesis, and genes encoding transcriptional regulators and secreted proteins. These genes areof interest to improve A. niger as a cell factory.
Plasmids, fungal strains and growth conditions
The wild-type A. niger strain N402 [13] and its derivative N402ΔflbA [8] were used in this study. These strains were grown as sandwiched colonies between porous polycarbonate membranes (0.1 μm pores, 76 mm diameter; Profiltra, Almere, The Netherlands) placed on top of solidified minimal medium with 25 mM xylose as a carbon source [4-6]. Cultures were inoculated with a 1 mm mycelial plug and grown at 30°C .
RNA preparation and analysis
RNA was isolated from a biological duplicate of concentric zones of 7-day-old colonies. Zone 1 represented the most central zone, zone 3 an intermediate zone and zone 5, the peripheral zone [6]. Mycelium of zones was harvested from three colonies, frozen in liquid nitrogen and ground with a TissueLyser (Qiagen, Venlo, The Netherlands) in a 2 ml Eppendorf tube with two metal balls (4.76 in diameter) for 1 min at 25 Hz. The frozen material was taken up in 1 ml TRIzol reagent (Invitrogen, Bleiswijk, The Netherlands) by vortexing. Samples were incubated for 2 min after mixing with 200 μl chloroform. This was followed by centrifugation at 10000 g for 10 min. RNA was purified using an RNA clean up column (Machery Nagel, Düren, Germany), after addition of 1 volume 70% EtOH to the water phase. After loading the sample on the column, it was centrifuged for 30 sec at 10000 rpm. This was followed by addition of 600 μl RA3 buffer (provided by the RNA-clean up kit). After 2 min centrifugation at 10000 g, 250 μl RNA3 was added, followed by another 2 min centrifugation at 10000 g. RNA was eluted after a 10 min incubation in two steps with 40 μl and 50 μl RNAse free water. The eluted RNA was pooled and 1 μl was checked for concentration and purity using the nanodrop ND- 1000 spectrophotometer (Nanodrop Technology, Wilmington, USA) and the Bioanalyser 2100 (Agilent Technologies, Santa Clara, USA) (Supplemental Figure 1), respectively.
cDNA labeling, microarray hybridization and data analysis
cDNA labeling, microarray hybridization and scanning were performed at ServiceXS (Leiden, The Netherlands), according to Affymetrix protocols. From each RNA sample, 100 ng was used to synthesize biotin-labeled cRNA with the Affymetrix 3’ IVT-Express Labeling Kit. Quality of cRNA was checked with the nano-lab-on-a chip Bioanalyzer 2100 (Agilent Technology). 15 μg cRNA was fragmented and half of it was used for hybridization on Affymetrix A. niger GeneChips [14]. After an automated process of washing and staining, absolute values of expression were calculated from the scanned array using the Affymetrix Command Console v1 software. The Affymetrix probe sets on the chip represent 14,259 annotated ORFs and genetic elements of A. niger [14]; (GEO [15] under accession no. GPL6758). The A. niger array data of this study have been deposited in the GEO database under accession number GSE44391.MAS5.0 absent/present calls showed that on average 50% and 48% of the genes were expressed in colonies of the wild-type and the ΔflbA strain, respectively. Genedata Expressionist and Genedata Analyst were used for normalization and statistical analysis of the arrays (Genedata, Basel, Switzerland). The arrays were condensed with the RMA algorithm and normalized on the quantile. Statistical assessment of differential expression was performed with t-test (p ≤ 0.01, using a false discovery rate [BHQ] of ≤ 0.05) or by ANOVA (p ≤ 0.01, BHQ ≤ 0.05). Differentially expressed genes (change in expression ≥ 2-fold) were clustered using K-Means clustering. A differentially expressed set of genes obtained by ANOVA was used for a maximal paired contrast analysis to find the zone or strain where the highest mean-difference was observed. The Functional Catalogue (FunCat, Munich Information Center for Protein Sequence, Munich, Germany) [16] was used for functional classification of genes. Fisher’s exact test was used to identify over-and under-represented functional classes in the sets of differentially expressed genes (p ≤ 0.001, BHQ ≤ 0.05).
cDNA synthesis and quantitative PCR analysis
cDNA was synthesized from biological duplicates of total RNA of zones 1, 3 and 5 of wild type and ΔflbA colonies using the QuantiTect Reverse Transcription Kit (Qiagen). QPCR was performed on the cDNA using the ABI Prism 7900HT SDS and SYBR Green chemistry (Applied Biosystems, Life Sciences, Foster City, USA). A run without cDNA was used as a negative control for every condition. Cycle threshold (Ct) levels were measured in triplicate for actin (An15g00560), faeA (An09g00120), glaA (An03g06550) and 18S rDNA (An03e03200). Primers were designed according to the recommendations of the PCR master-mix manufacturer (Applied Biosystems, Life Sciences, Foster City, USA), and were described previously [17]. The Ct values of 18S were used to normalize the qPCR data. For qPCR, each cycle Ct-change in qPCR represents a two-fold change in expression (2-Δ(ΔCT), where ΔCt=Cttarget gene-Ct18S gene and Δ(ΔCT)=ΔCTmutant-ΔCTwild type)..
Five concentric zones can be distinguished in 7-day-old sandwiched colonies of A. niger [4,6]. Zone 1, 3 and 5 represent the most inner zone, an intermediate zone, and the most outer zone of the colony, respectively. Gene expression was assessed in these three zones of xylose-grown colonies of the wild-type and the ΔflbA strain of A. niger using Affymetrix microarrays (Supplemental Table 1). QPCR of glaA ,faeA, and actin (Supplemental Table 2) was used to verify micro-array data (Supplemental Table 3). Fold changes of these genes between zone 1 of wild-type and zone 1 of ΔflbA, zone 3 of wild-type and zone 3 of ΔflbA, and zone 5 of wild-type and zone 5 of ΔflbA as obtained with micro-array analysis and QPCR showed a Pearson’s correlation coefficient ≥ 0.97 (Supplemental Table 4).
| wt up intermediate/ center | wt up periphery | ΔflbAup intermediate/ center | ΔflbAup periphery | |
| 01 Metabolism | O | O | O | O |
| 01.01.10 amino acid degradation (catabolism) | O | O | ||
| 01.03.19 nucleotide transport | O | |||
| 01.05.01 C-compound and carbohydrate utilization | O | |||
| 02 Energy | O | O | ||
| 02.11.05 accessory proteins of electron transport and membrane-associated energy conservation | O | |||
| 03 Cell cycle and dna processing | U | |||
| 04 Transcription | U | U | U | |
| 04.05.01 mRNA synthesis | U | U | ||
| 05 Protein synthesis | O | U | ||
| 08 Cellular transport and transport mechanisms | O | |||
| 40 Subcellular localisation | U | |||
| 99 Unclassified proteins | U | U | U | U |
Table 1: Over- (O) and under- (U) representation of functional gene classes in the pool of genes that are ≥ 2 fold up- or down-regulated in the central zone 1 and intermediate zone 3 when compared to the peripheral zone 5of wild-type and ΔflbA colonies.
| zone 1 Up | zone 1 Down | zone 3 Up | zone 3 Down | zone 5 Up | zone 5 Down | |
| 01 Metabolism | O | O | O | O | O | |
| 01.01.07 amino acid transport | O | |||||
| 01.05.01 C-compound and carbohydrate utilization | O | |||||
| 03 Cell cycle and DNA processing | U | |||||
| 06.13.99 other proteolytic degradation | O | |||||
| 08 Cellular transport and transport mechanisms | O | |||||
| 99 Unclassified proteins | U | U | U | U |
Table 2: Over- (O) and under- (U) representation of functional gene classes in the pool of genes that are ≥ 2 fold up- or down-regulated in the ΔflbA strain when compared to the wild-type.
| up-regulated in ΔflbA | Down-regulated in ΔflbA | |
| 01 Metabolism | O | O |
| 01.01.07 amino acid transport | ||
| 01.05.01 C-compound and carbohydrate utilization | O | |
| 02 Energy | U | |
| 03 Cell cycle and DNA Processing | U | |
| 03.03.01 mitotic cell cycle and cell cycle control | U | |
| 04 Transcription | U | |
| 05 Protein synthesis | U | |
| 08 Cellular transport and transport mechanisms | O | |
| 40 Subcellular localisation | U | |
| 99 Unclassified proteins | U | U |
Table 3: Over- (O) and under- (U) representation of functional gene classes in the pool of genes that are ≥ 2 fold up- or down-regulated between the mean expression values of zones 1, 3, and 5 of the ΔflbA strain and those of the wild-type strain.
| Annotation | Gene name* | Description | wild-type zone 1 | wild-type zone 3 | wild-type zone 5 | ΔflbA zone 1 | ΔflbA zone 3 | ΔflbA zone 5 | Regulation | sigP |
|---|---|---|---|---|---|---|---|---|---|---|
| Top 50 down-regulated genes inthe ΔflbA strain compared to wildtype | ||||||||||
| An05g01730 | uncharacterized | hypothetical protein | 11442 ± 3316 | 14891 ± 315 | 4511 ± 2021 | 42 ± 7 | 37 ± 0 | 27 ± 1 | Down | y |
| An09g00840 | uncharacterized | weak similarity to antigenic cell wall galactomannoprotein MP1 - Aspergillus fumigatus | 3153 ± 2599 | 10725 ± 146 | 3462 ± 103 | 29 ± 11 | 35 ± 22 | 14 ± 2 | Down | y |
| An18g05480 | uncharacterized | strong similarity to alcohol oxidase AOX1 - Pichia pastoris | 7168 ± 839 | 7322 ± 2676 | 3105 ± 1353 | 46 ± 2 | 44 ± 3 | 42 ± 5 | Down | n |
| An16g05920 | uncharacterized | weak similarity to surface recognition protein PTH11 - Magnaporthe grisea | 4282 ± 1762 | 6936 ± 1040 | 5405 ± 665 | 48 ± 1 | 47 ± 11 | 44 ± 1 | Down | n |
| An07g06460 | uncharacterized | similarity to C-7 hydroxycephem methyltransferase coupling protein of patent WO9529253-A1 - Streptomyces lactamdurans | 1807 ± 56 | 3006 ± 541 | 4779 ± 1295 | 29 ± 5 | 24 ± 5 | 28 ± 0 | Down | n |
| An16g05930 | uncharacterized | strong similarity to predicted protein An08g06890 - Aspergillus niger | 5034 ± 1765 | 7101 ± 440 | 6203 ± 1646 | 63 ± 18 | 58 ± 11 | 58 ± 9 | Down | n |
| An01g06890 | uncharacterized | similarity to hypothetical peptide synthetase pesA - Metarhizium anisopliae | 1497 ± 1410 | 2785 ± 1060 | 5197 ± 1630 | 88 ± 10 | 29 ± 2 | 18 ± 0 | Down | n |
| An16g05910 | uncharacterized | similarity to cholesterol 7alpha-hydroxylase CYP7 - Sus scrofa | 902 ± 636 | 1646 ± 590 | 2001 ± 725 | 17 ± 3 | 25 ± 5 | 18 ± 2 | Down | y |
| An03g00690 | uncharacterized | hypothetical protein | 1261 ± 1329 | 2284 ± 1815 | 2321 ± 1481 | 21 ± 7 | 28 ± 1 | 26 ± 2 | Down | n |
| An08g02330 | uncharacterized | strong similarity to multidrug resistance protein MLP-2 - Rattus norvegicus | 2009 ± 37 | 2116 ± 179 | 1500 ± 529 | 45 ± 2 | 29 ± 3 | 22 ± 4 | Down | n |
| An07g06480 | uncharacterized | similarity to cytochrome 4F8 cyp4F8 - Homo sapiens | 420 ± 130 | 586 ± 213 | 753 ± 142 | 9 ± 1 | 10 ± 2 | 9 ± 1 | Down | y |
| An05g01710 | uncharacterized | strong similarity to hypothetical protein An16g07720 - Aspergillus niger | 1738 ± 1486 | 2310 ± 357 | 387 ± 170 | 19 ± 2 | 18 ± 2 | 19 ± 3 | Down | n |
| An01g00530 | pepB | proteinase aspergillopepsin II - Aspergillus niger | 682 ± 649 | 3677 ± 2802 | 6268 ± 3005 | 44 ± 5 | 35 ± 1 | 34 ± 0 | Down | y |
| An01g06860 | uncharacterized | strong similarity to hypothetical Fum9p protein - Gibberella moniliformis | 1135 ± 1295 | 2702 ± 926 | 5766 ± 2392 | 67 ± 10 | 36 ± 3 | 31 ± 0 | Down | n |
| An01g06870 | uncharacterized | strong similarity to hypothetical protein Fum8p - Gibberella moniliformis | 581 ± 599 | 1632 ± 396 | 3427 ± 1234 | 50 ± 2 | 26 ± 1 | 21 ± 6 | Down | n |
| An02g08300 | uncharacterized | strong similarity to the hypothetical protein encoded by An11g06450 - Aspergillus niger | 842 ± 71 | 710 ± 230 | 2085 ± 860 | 30 ± 1 | 21 ± 2 | 24 ± 8 | Down | n |
| An15g07700 | protD | strong similarity to aspergillopepsin II precursor (acid proteinase A) - Aspergillus niger | 649 ± 452 | 5044 ± 2370 | 9630 ± 101 | 75 ± 5 | 76 ± 5 | 65 ± 2 | Down | y |
| An09g00670 | gelD | strong similarity to beta (1-3) glucanosyltransferase Gel3p - Aspergillus fumigatus | 1596 ± 565 | 2244 ± 607 | 2063 ± 821 | 45 ± 1 | 39 ± 6 | 59 ± 10 | Down | y |
| An16g06570 | uncharacterized | hypothetical protein | 7069 ± 619 | 3576 ± 3514 | 1162 ± 1292 | 62 ± 4 | 52 ± 8 | 62 ± 4 | Down | y |
| An02g00090 | uncharacterized | strong similarity to prolidase - Aureobacterium esteraromaticum | 6755 ± 2149 | 4420 ± 2149 | 707 ± 467 | 134 ± 13 | 58 ± 3 | 36 ± 11 | Down | n |
| An05g02450 | uncharacterized | similarity to halogenase bhaA from patent DE19926770-A1 - Amycolatopsis mediterranei | 663 ± 62 | 1035 ± 59 | 469 ± 33 | 25 ± 6 | 18 ± 2 | 18 ± 1 | Down | y |
| An07g08950 | eglC | endoglucanase B eglB - Aspergillus niger | 2511 ± 570 | 11775 ± 532 | 7793 ± 1370 | 52 ± 29 | 366 ± 244 | 422 ± 77 | Down | y |
| An08g02300 | uncharacterized | weak similarity to enniatin synthetase - Fusarium scirpi[truncated ORF] | 1123 ± 198 | 1542 ± 488 | 1754 ± 1024 | 79 ± 8 | 53 ± 1 | 22 ± 5 | Down | n |
| An01g06900 | sm to amyR- A. ory | weak similarity to transcription activator amyR - Aspergillus oryzae | 758 ± 96 | 1056 ± 406 | 654 ± 233 | 34 ± 8 | 25 ± 1 | 21 ± 1 | Down | n |
| An12g01320 | ppoD | strong similarity to linoleate diol synthase - Gaeumannomyces graminis | 1123 ± 612 | 1043 ± 345 | 132 ± 72 | 18 ± 2 | 20 ± 3 | 14 ± 2 | Down | y |
| An01g06880 | uncharacterized | similarity to dihydroflavonol 4-reductase BAA12723.1 - Rosa hybrid cultivar | 625 ± 534 | 1769 ± 746 | 3403 ± 1745 | 110 ± 3 | 36 ± 3 | 27 ± 6 | Down | n |
| An02g08330 | uncharacterized | strong similarity to sequence 253 from Patent WO0100804 - Corynebacterium glutamicum | 627 ± 234 | 624 ± 177 | 877 ± 309 | 27 ± 2 | 25 ± 1 | 24 ± 6 | Down | y |
| An01g06840 | uncharacterized | strong similarity to acid-CoA ligase Fat2p - Saccharomyces cerevisiae | 419 ± 441 | 1295 ± 731 | 3234 ± 1795 | 52 ± 0 | 34 ± 1 | 29 ± 4 | Down | n |
| An03g06670 | uncharacterized | weak similarity to myosin-like protein MLP1 - Saccharomyces cerevisiae | 5998 ± 1898 | 6281 ± 1029 | 1761 ± 437 | 171 ± 1 | 171 ± 7 | 118 ± 9 | Down | n |
| An01g06830 | uncharacterized | similarity to ketosphinganine reductase Tsc10p - Saccharomyces cerevisiae | 418 ± 405 | 1031 ± 265 | 2067 ± 700 | 46 ± 1 | 33 ± 6 | 24 ± 3 | Down | n |
| An01g06850 | uncharacterized | similarity to 4-hydroxybutyrate dehydrogenase - Alcaligenes eutrophus | 967 ± 1081 | 1917 ± 437 | 4224 ± 1181 | 112 ± 1 | 54 ± 2 | 44 ± 4 | Down | n |
| An03g00640 | uncharacterized | similarity to neutral amino acid permease mtr - Neurospora crassa[truncated ORF] | 2950 ± 1037 | 2645 ± 295 | 2479 ± 1469 | 149 ± 28 | 122 ± 9 | 57 ± 2 | Down | n |
| An01g11670 | eglA | strong similarity to endo-beta-1,4-glucanase A eglA - Emericella nidulans | 638 ± 408 | 2367 ± 12 | 589 ± 232 | 21 ± 2 | 42 ± 24 | 58 ± 1 | Down | y |
| An07g08940 | uncharacterized | similarity to acetyl-esterase I of patent WO9502689-A - Aspergillus aculeatus | 242 ± 13 | 1268 ± 407 | 914 ± 553 | 20 ± 1 | 31 ± 7 | 26 ± 0 | Down | y |
| An11g07020 | uncharacterized | strong similarity to the hypothetical protein encoded by An07g00200 - Aspergillus niger | 3237 ± 1203 | 2067 ± 1234 | 1620 ± 142 | 125 ± 37 | 97 ± 1 | 54 ± 11 | Down | n |
| An07g09330 | cbhA | cellulose 1,4-beta-cellobiosidase cbhA from patent WO9906574-A1- Aspergillus niger | 1157 ± 1121 | 7883 ± 2929 | 3034 ± 1406 | 67 ± 16 | 130 ± 62 | 170 ± 76 | Down | y |
| An09g06200 | uncharacterized | strong similarity to PTH11 transmembrane protein - Magnaporthe grisea strain 4091-5-8 | 5976 ± 1928 | 5351 ± 2683 | 3632 ± 1622 | 442 ± 58 | 211 ± 45 | 82 ± 12 | Down | y |
| An04g09990 | uncharacterized | strong similarity to 2,5-dichloro-2,5-cyclohexadiene-1,4-diol dehydrogenase linC - Sphingomonas paucimobilis | 749 ± 308 | 422 ± 138 | 106 ± 4 | 36 ± 6 | 34 ± 6 | 34 ± 5 | Down | y |
| An04g03840 | uncharacterized | similarity to microtubule binding protein D-CLIP-190 - Drosophila melanogaster | 365 ± 35 | 373 ± 52 | 257 ± 17 | 16 ± 0 | 14 ± 0 | 12 ± 3 | Down | n |
| An05g01720 | uncharacterized | strong similarity to hypothetical protein An06g00950 - Aspergillus niger | 1552 ± 1286 | 2068 ± 211 | 368 ± 231 | 47 ± 7 | 42 ± 7 | 41 ± 1 | Down | n |
| An09g02990 | uncharacterized | similarity to probable Sua5 protein APE2397 - Aeropyrum pernix | 618 ± 224 | 781 ± 58 | 344 ± 112 | 25 ± 6 | 26 ± 0 | 24 ± 2 | Down | n |
| An04g06750 | uncharacterized | similarity to hypothetical transmembrane protein - Candida albicans | 491 ± 201 | 386 ± 153 | 489 ± 169 | 25 ± 4 | 20 ± 2 | 19 ± 2 | Down | n |
| An07g06490 | uncharacterized | strong similarity to insulin-degrading enzyme IDE - Rattus norvegicus | 213 ± 37 | 361 ± 124 | 474 ± 112 | 17 ± 0 | 17 ± 0 | 16 ± 2 | Down | n |
| An12g05390 | uncharacterized | weak similarity to integral membrane protein PTH11 - Magnaporthe grisea | 2079 ± 127 | 1355 ± 14 | 84 ± 32 | 38 ± 0 | 31 ± 2 | 30 ± 3 | Down | y |
| An16g02910 | uncharacterized | strong similarity to hypothetical protein CC0533 - Caulobacter crescentus | 865 ± 51 | 1033 ± 82 | 613 ± 238 | 52 ± 1 | 40 ± 8 | 42 ± 10 | Down | y |
| An08g11680 | uncharacterized | strong similarity to 2,5-dicloro-2,5-cyclohexadiene-1,4-diol dehydrogenase linC - Pseudomonas paucimobilis | 320 ± 4 | 214 ± 41 | 125 ± 4 | 56 ± 0 | 68 ± 11 | 61 ± 1 | Down | y |
| An01g06930 | uncharacterized | strong similarity to polyketide synthase FUM5 - Gibberella moniliformis | 714 ± 704 | 1442 ± 246 | 2506 ± 866 | 110 ± 1 | 60 ± 2 | 46 ± 1 | Down | n |
| An14g03130 | uncharacterized | hypothetical protein | 1109 ± 560 | 752 ± 164 | 262 ± 94 | 49 ± 11 | 29 ± 4 | 25 ± 2 | Down | y |
| An08g03760 | uncharacterized | similarity to hypothetical protein Rv3472 - Mycobacterium tuberculosis | 191 ± 86 | 197 ± 37 | 843 ± 343 | 16 ± 1 | 16 ± 1 | 23 ± 5 | Down | n |
| An08g05230 | uncharacterized | strong similarity to putative endoglucanase IV - Trichoderma reesei | 1152 ± 1281 | 2411 ± 2729 | 648 ± 116 | 55 ± 5 | 58 ± 11 | 46 ± 5 | Down | y |
| Top 50 up-regulated genesin the ΔflbA strain compared to wildtype | ||||||||||
| An08g08490 | uncharacterized | similarity to the calcium-independent phospholipase A2 2 - Homo sapiens | 143 ± 8 | 118 ± 7 | 85 ± 3 | 728 ± 0 | 847 ± 91 | 775 ± 76 | Up | n |
| An07g06240 | uncharacterized | strong similarity to ferrioxamine B permease sit1 - Saccharomyces cerevisiae | 88 ± 24 | 87 ± 8 | 94 ± 8 | 323 ± 28 | 397 ± 32 | 1915 ± 383 | Up | n |
| An03g03620 | uncharacterized | strong similarity to multidrug resistance protein atrD - Aspergillus nidulans | 18 ± 1 | 22 ± 2 | 26 ± 10 | 83 ± 5 | 110 ± 1 | 378 ± 123 | Up | y |
| An08g10830 | uncharacterized | strong similarity to geranylgeranyl pyrophosphate synthase ggpps - Gibberella fujikuroi | 65 ± 5 | 58 ± 0 | 68 ± 18 | 412 ± 9 | 388 ± 31 | 555 ± 30 | Up | n |
| An01g12200 | uncharacterized | similarity to hypothetical protein F10B6.29 - Arabidopsis thaliana | 112 ± 53 | 68 ± 23 | 94 ± 12 | 741 ± 26 | 726 ± 82 | 429 ± 36 | Up | n |
| An09g00270 | aglC | alpha-galactosidase C aglC - Aspergillus niger[truncated ORF] | 89 ± 18 | 85 ± 22 | 760 ± 428 | 879 ± 73 | 1158 ± 119 | 1805 ± 251 | Up | nc |
| An07g04900 | uncharacterized | strong similarity to mRNA sequence of cDNA clone 2589 - Aspergillus niger | 1065 ± 70 | 929 ± 155 | 364 ± 239 | 5638 ± 245 | 5796 ± 687 | 3483 ± 369 | Up | y |
| An11g09170 | uncharacterized | similarity to the secreted aspartic proteinase SAP8 - Candida albicans | 48 ± 5 | 55 ± 11 | 42 ± 1 | 316 ± 18 | 441 ± 6 | 290 ± 41 | Up | y |
| An14g01840 | uncharacterized | similarity to hypothetical temperature-shock induced protein TIR3 - Saccharomyces cerevisiae | 227 ± 122 | 258 ± 66 | 426 ± 36 | 1679 ± 739 | 2311 ± 995 | 2553 ± 877 | Up | y |
| An11g02600 | uncharacterized | strong similarity to PT2/PHT4 Phosphate transporter - Arabidopsis thaliana | 50 ± 15 | 60 ± 10 | 104 ± 14 | 483 ± 47 | 653 ± 132 | 394 ± 127 | Up | n |
| An16g07040 | sm to btgE -A. nid | similarity to beta-1,3-glucanosyltransferase BGT1 - Aspergillus fumigatus[truncated ORF] | 502 ± 224 | 676 ± 353 | 2375 ± 144 | 5982 ± 202 | 6554 ± 1346 | 7358 ± 380 | Up | y |
| An17g00120 | uncharacterized | strong similarity to major facilitator superfamily transporter protein mfs1 - Botrytis cinerea | 30 ± 6 | 26 ± 0 | 24 ± 2 | 237 ± 3 | 221 ± 13 | 140 ± 13 | Up | n |
| An16g08360 | uncharacterized | weak similarity to cytochrome c1 of ubiquinol--cytochrome-c reductase - Paracoccus denitrificans[truncated ORF] | 64 ± 10 | 75 ± 12 | 158 ± 35 | 610 ± 68 | 591 ± 130 | 852 ± 12 | Up | n |
| An11g01810 | uncharacterized | weak similarity to probable membrane protein YBR005w - Saccharomyces cerevisiae | 55 ± 6 | 62 ± 5 | 96 ± 20 | 438 ± 9 | 541 ± 62 | 564 ± 34 | Up | n |
| An02g07930 | ppoC | strong similarity to linoleate diol synthase precursor - Gaeumannomyces graminis | 180 ± 34 | 126 ± 52 | 38 ± 2 | 983 ± 279 | 747 ± 19 | 464 ± 32 | Up | n |
| An09g01240 | uncharacterized | strong similarity to phospholipase B - Penicillium notatum | 78 ± 7 | 63 ± 9 | 104 ± 3 | 499 ± 142 | 727 ± 111 | 627 ± 178 | Up | y |
| An01g06280 | uncharacterized | strong similarity to IgE-binding protein - Aspergillus fumigatus | 267 ± 123 | 341 ± 167 | 136 ± 4 | 2010 ± 28 | 2634 ± 280 | 1002 ± 145 | Up | y |
| An13g01250 | uncharacterized | strong similarity to the yeast siderophore-iron transporter for enterobactin Enb1 - Saccharomyces cerevisiae. | 37 ± 2 | 35 ± 3 | 36 ± 3 | 183 ± 64 | 331 ± 79 | 415 ± 58 | Up | n |
| An14g07130 | uncharacterized | strong similarity to neutral amino acid permease mtr - Neurospora crassa | 95 ± 8 | 70 ± 4 | 93 ± 8 | 481 ± 32 | 602 ± 135 | 1245 ± 123 | Up | n |
| An11g04810 | aox1 | alternative oxidase Aox1 - Aspergillus niger | 43 ± 8 | 54 ± 32 | 47 ± 20 | 279 ± 254 | 470 ± 353 | 890 ± 194 | Up | n |
| An15g03940 | uncharacterized | strong similarity to monosaccharide transporter Mst-1 - Amanita muscaria | 244 ± 17 | 202 ± 47 | 2589 ± 838 | 3196 ± 21 | 3689 ± 943 | 8423 ± 210 | Up | y |
| An13g00510 | uncharacterized | strong similarity to hexokinase 1 hxk1 - Schizosaccharomyces pombe | 199 ± 43 | 202 ± 28 | 67 ± 10 | 3335 ± 329 | 2920 ± 299 | 223 ± 64 | Up | n |
| An18g01290 | uncharacterized | strong similarity to predicted protein An13g01340 - Aspergillus niger | 26 ± 2 | 24 ± 2 | 38 ± 10 | 233 ± 53 | 218 ± 65 | 405 ± 13 | Up | y |
| An01g07000 | uncharacterized | strong similarity to C-14 sterol reductase ERG24 - Saccharomyces cerevisiae | 89 ± 14 | 77 ± 2 | 270 ± 166 | 498 ± 24 | 1009 ± 531 | 3138 ± 50 | Up | y |
| An10g00680 | uncharacterized | strong similarity to H+-ATPase V0 domain 17 KD subunit, vacuolar, CUP5 - Saccharomyces cerevisiae | 443 ± 342 | 220 ± 116 | 119 ± 50 | 2783 ± 380 | 2761 ± 26 | 1102 ± 42 | Up | y |
| An16g09040 | uncharacterized | strong similarity to N-acetylglucosamine-6-phosphate deacetylase CaNAG2 - Candida albicans | 629 ± 51 | 516 ± 71 | 81 ± 7 | 6612 ± 950 | 6653 ± 1009 | 637 ± 136 | Up | n |
| An01g00390 | uncharacterized | hypothetical protein | 45 ± 2 | 47 ± 2 | 319 ± 99 | 542 ± 59 | 685 ± 35 | 1891 ± 74 | Up | n |
| An16g01850 | uncharacterized | similarity to blastomyces yeast phase-specific protein 1 bys1 - Ajellomyces dermatitidis | 2984 ± 2759 | 1197 ± 780 | 166 ± 54 | 12215 ± 313 | 12035 ± 1137 | 3132 ± 648 | Up | y |
| An16g01880 | Lipanl | strong similarity to lysophospholipase - Aspergillus foetidus | 648 ± 245 | 795 ± 248 | 454 ± 202 | 10644 ± 663 | 10585 ± 2214 | 2676 ± 160 | Up | y |
| An14g02940 | uncharacterized | strong similarity to L-sorbose dehydrogenase, FAD dependent - Gluconobacter oxydans | 212 ± 101 | 153 ± 71 | 139 ± 92 | 2065 ± 372 | 1565 ± 349 | 1690 ± 279 | Up | n |
| An07g03570 | uncharacterized | strong similarity to sorbitol utilization protein sou2 - Candida albicans | 83 ± 7 | 85 ± 17 | 1534 ± 228 | 1757 ± 42 | 1613 ± 167 | 6032 ± 312 | Up | n |
| An18g03360 | uncharacterized | similarity to the proteophosphoglycan ppg1 - Leishmania major | 32 ± 19 | 46 ± 26 | 27 ± 5 | 473 ± 19 | 503 ± 4 | 242 ± 2 | Up | y |
| An14g06980 | uncharacterized | strong similarity to delta-12 fatty acid desaturase - Mortierella alpina | 33 ± 3 | 36 ± 2 | 301 ± 112 | 465 ± 89 | 750 ± 251 | 1939 ± 15 | Up | n |
| An05g00790 | uncharacterized | strong similarity to nodulin GmNOD53b - Glycine max[truncated ORF] | 25 ± 15 | 20 ± 4 | 12 ± 1 | 213 ± 57 | 341 ± 30 | 161 ± 9 | Up | n |
| An03g06660 | uncharacterized | strong similarity to peptide transporter ptr2 - Arabidopsis thaliana | 2166 ± 1123 | 1709 ± 1392 | 80 ± 15 | 12416 ± 598 | 10787 ± 865 | 3842 ± 47 | Up | n |
| An18g01320 | uncharacterized | strong similarity to extracellular protease precursor BAR1 - Saccharomyces cerevisiae | 148 ± 76 | 129 ± 56 | 328 ± 31 | 2146 ± 67 | 2897 ± 61 | 2501 ± 161 | Up | y |
| An05g00800 | uncharacterized | similarity to nodulin GmNOD53b - Glycine max[truncated ORF] | 47 ± 21 | 31 ± 11 | 22 ± 1 | 395 ± 130 | 667 ± 69 | 333 ± 6 | Up | n |
| An12g09870 | uncharacterized | hypothetical protein | 50 ± 0 | 57 ± 1 | 56 ± 0 | 571 ± 232 | 432 ± 82 | 2224 ± 250 | Up | n |
| An11g00100 | uncharacterized | strong similarity to triacylglycerol lipase LIP5 - Candida rugosa | 71 ± 4 | 64 ± 3 | 59 ± 8 | 800 ± 192 | 866 ± 397 | 2265 ± 77 | Up | y |
| An01g01630 | uncharacterized | strong similarity to hypothetical protein An09g00510 - Aspergillus niger | 69 ± 71 | 36 ± 26 | 96 ± 40 | 1151 ± 99 | 830 ± 115 | 802 ± 17 | Up | y |
| An01g07730 | uncharacterized | weak similarity to TcSL-2 protein precursor - Toxocara cani | 118 ± 74 | 64 ± 3 | 63 ± 5 | 1697 ± 86 | 1478 ± 312 | 1257 ± 12 | Up | y |
| An15g03550 | uncharacterized | weak similarity to protopectinase patent WO9806832-A1 - Bacillus subtilis | 57 ± 44 | 57 ± 45 | 20 ± 0 | 954 ± 34 | 1066 ± 21 | 330 ± 22 | Up | y |
| An16g06950 | uncharacterized | strong similarity to carboxylic acid transport protein JEN1 - Saccharomyces cerevisiae | 31 ± 8 | 26 ± 0 | 23 ± 5 | 837 ± 53 | 725 ± 9 | 357 ± 20 | Up | n |
| An03g01770 | uncharacterized | strong similarity to the EST an_3645 - Aspergillus niger | 758 ± 657 | 336 ± 176 | 55 ± 12 | 10029 ± 207 | 8451 ± 0 | 1933 ± 247 | Up | y |
| An03g05360 | uncharacterized | strong similarity to neutral amino acid permease mtr - Neurospora crassa | 39 ± 0 | 36 ± 2 | 31 ± 2 | 1673 ± 148 | 1362 ± 88 | 771 ± 2 | Up | y |
| An14g04210 | uncharacterized | similarity to hypothetical protein An07g05660 - Aspergillus niger | 30 ± 0 | 35 ± 7 | 31 ± 0 | 931 ± 174 | 1386 ± 104 | 2511 ± 5 | Up | y |
| An03g06220 | sm to gelD- A. nid | strong similarity to beta (1-3) glucanosyltransferase GEL3 - Aspergillus fumigatus | 42 ± 3 | 48 ± 11 | 58 ± 3 | 1406 ± 256 | 1874 ± 54 | 5043 ± 547 | Up | y |
| An02g08560 | uncharacterized | similarity to probable dioxygenase SCOEDB - Streptomyces coelicolor | 22 ± 3 | 23 ± 3 | 19 ± 0 | 797 ± 22 | 1048 ± 83 | 1682 ± 135 | Up | y |
| An02g08130 | uncharacterized | similarity to hypothetical protein 2SCG18.24 - Streptomyces coelicolor | 46 ± 3 | 48 ± 9 | 41 ± 5 | 2045 ± 64 | 2532 ± 185 | 3306 ± 224 | Up | y |
| An09g05050 | uncharacterized | questionable ORF | 25 ± 1 | 24 ± 4 | 22 ± 3 | 3522 ± 1024 | 2872 ± 506 | 8559 ± 471 | Up | y |
Table 4: Hybridization values in colony zones 1, 3 and 5 of top 50 up- and down-regulated genes of the ΔflbA strain when compared to wild-type.*Gene name or its closest homologue. A. nid: A. nidulans; A. ory: A. oryzae.
Principal component analysis (PCA) showed that the transcriptomes of the wild-type zones and those of the ΔflbA strain cluster in the first component of the analysis. This component accounts for 54% of the variation in the datasets (Figure 2). The second component of the analysis accounted for 27% of the variation and separates the transcriptomes from zones 1 and 3 of the wild-type and the ΔflbA strain from zone 5 of these strains (Figure 2). Expression in the central zone 1 and the intermediate zone 3 of the wild-type correlated to a high extent (Pearson’sr2 ≥ 0.98), when compared to the duplicates of these zones (r2 ≥ 0.97 and ≥ 0.98, respectively). Wild-type zone 5 was more distinct from zone 3 and 1 (r2=0.91 and 0.93). The expression profile of zone 5 was also most different within the ΔflbA colony (Figure 2). The fact that zone 5 has the most distinct expression profile was also illustrated by the number of genes, with a fold change in expression ≥ 2 between the zones of wild-type and ΔflbA colonies (Figure 3). The number of differentially expressed genes between zone 1 and zone 3 of the wildtype colony was 5, between zone 3 and zone 5, 119, and between zone 1 and zone 5, 325. These numbers were 0, 115 and 595 for the ΔflbA strain, respectively (Figure 3). Of these differentially expressed genes, only 138 were found in both thewild-type and the ΔflbA strain.
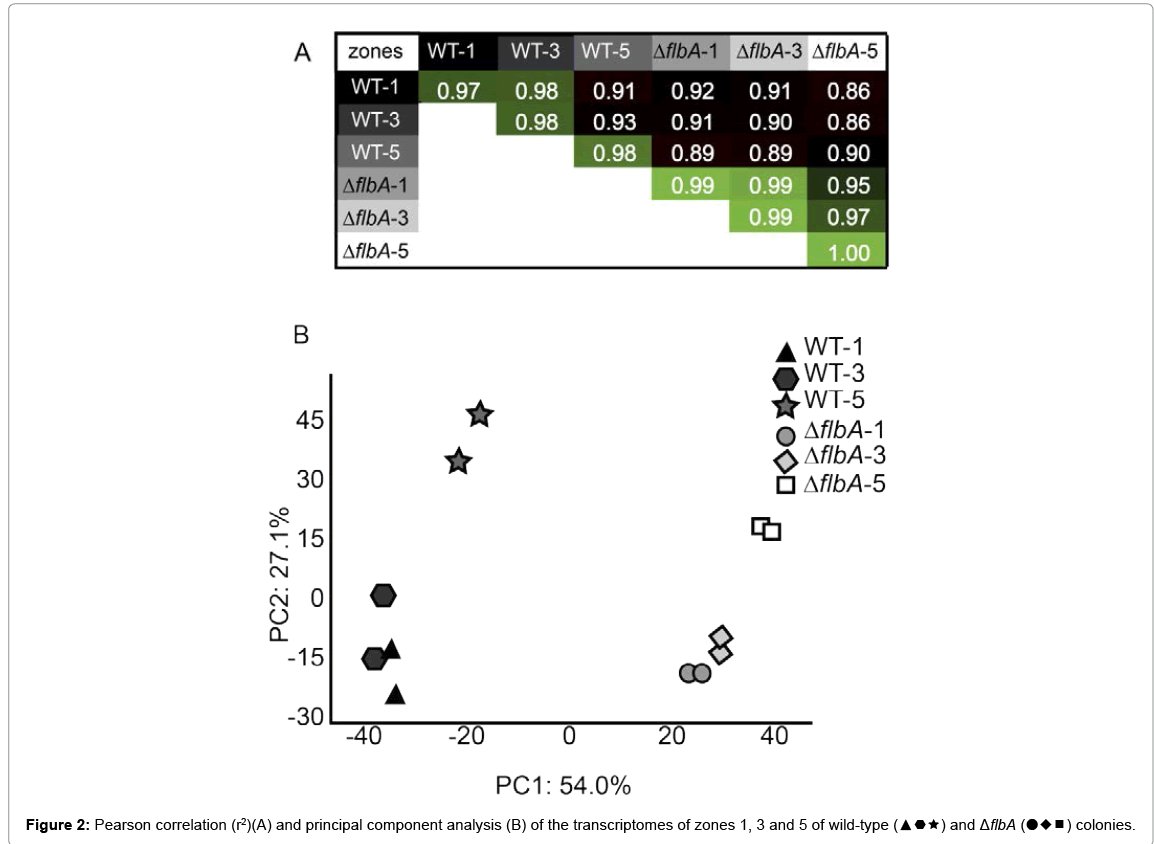
Figure 2: Pearson correlation (r2)(A) and principal component analysis (B) of the transcriptomes of zones 1, 3 and 5 of wild-type (▲▅★) and ΔflbA (●♦■) colonies.
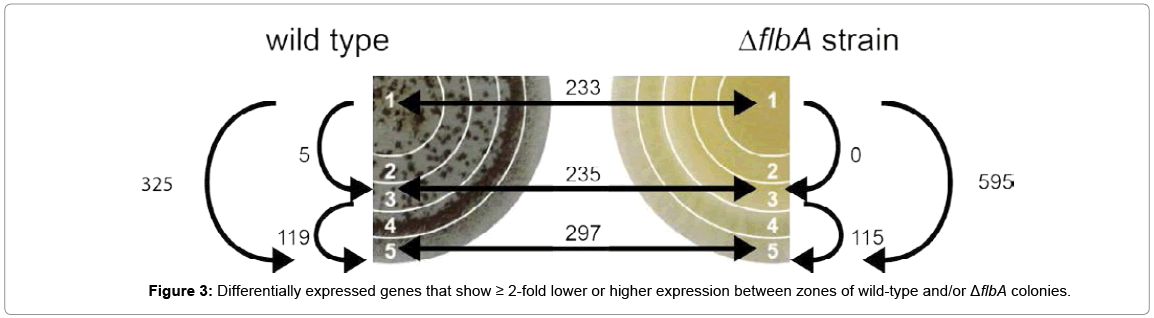
Figure 3: Differentially expressed genes that show ≥ 2-fold lower or higher expression between zones of wild-type and/or ΔflbA colonies.
In the next analysis, the number of genes was determined that are differentially expressed when zones of the wild-type and the ΔflbA strain were compared (Figure 3). The number of genes differentially expressed in zone 1 of the wild-type and the ΔflbA strain was 233. Of these, 104 and 129 were up- and down-regulated in the ΔflbA strain, respectively. A total of 235 genes were found to be differentially expressed inzone 3. Of these, 89 were up-regulated in ΔflbA, whereas 149 were up-regulated in the wild-type. In zone 5, 297 genes were differentially expressed, of which 126 and 153 were up- and downregulated in the ΔflbA strain, respectively.
Expression analysis of functional gene classes
Fisher’s exact test was used to determine whether functional gene classes [16] were over-or underrepresented in the set of genes that are differentially expressed in zones 1 and 3, compared to zone 5 of the wildtype and the ΔflbA strain (Table 1). The functional classes metabolism and amino acid degradation were overrepresented in the up-regulated genes of zones 1 and 3 of the wild-type colony (Table 1), whereas the functional classes metabolism, energy, accessory proteins of electron transport and membrane associated energy conservation and protein synthesis were overrepresented in the peripheral wild-type zone 5 (Table 1). The functional classes transcription, mRNA synthesis and unclassified proteins were under-represented in this cluster of genes, while only the latter functional group was underrepresented in zone 1 and 3 of the wild-type. In the up-regulated genes in the central and intermediate zone of the ΔflbA strain, the functional class metabolism was over-represented, whereas cell cycle and DNA processing, transcription, protein synthesis and unclassified proteins were underrepresented (Table 1). Metabolism, amino acid degradation, nucleotide transport, C-compound and carbohydrate utilization, energy and cellular transport and transport mechanisms were over-represented in the up-regulated genes in the peripheral zone of ΔflbA colonies (Table 1). On the other hand, transcription, mRNA synthesis, subcellular localization and unclassified proteins were under-represented.
Funcat analysis was performed on the differentially expressed genes between the zones of the wild-type and the ΔflbA colonies (Table 2). Up-regulated genes in zone 1 of ΔflbA compared to wild-type zone 1 were overrepresented in metabolism and amino acid transport, whereas unclassified proteins were underrepresented. Metabolism and unclassified proteins were also over- and underrepresented, respectively, in the down-regulated genes in zone 1 of ΔflbA compared to wild-type zone 1. The same was observed in the down-regulated genes in zone 3, whereas only the gene class metabolism was overrepresented in the up-regulated genes of this zone. Cellular transport and transport mechanisms were overrepresented in the up-regulated genes of the peripheral zone 5 of ΔflbA colonies. In contrast, the downregulated genes in this zone showed an over-representation of genes of the functional classes metabolism, C-compound and carbohydrate utilization and other proteolytic degradation, whereas cell cycle and DNA processing and unclassified proteins were underrepresented (Table 2).
Expression analysis of specific functional gene classes
The expression profiles of zones 1, 3 and 5 of the wild-type were pooled and compared with the pooled profiles of zones 1, 3 and 5 of the ΔflbA strain. In other words, for each gene the mean expression within the wild-type colony was compared to the mean expression within the ΔflbA strain. This was done to increase the statistical power of the comparison (comparing 6 instead of 2 arrays in each case). The number of genes up-regulated in ΔflbA colonies was 520, while 632 genes were down-regulated. Funcat analysis showed that the up-regulated genes are overrepresented in the functional categories metabolism andcellular transport and transport mechanisms, whereas transcription and unclassified proteins were underrepresented (Table 3). Down-regulated genes were overrepresented in metabolism and C-compound and carbohydrate utilization, and underrepresented in energy, cell cycle and DNA processing, mitotic cell cycle and cell cycle control, protein synthesis, subcellular localisation, and unclassified proteins (Table 3).
Twenty-six out of the 50 genes with the highest up-regulation (≥ 2-fold) in ΔflbA compared to wild-type are predicted to encode proteins with a signal sequence for secretion (sigP) (Table 4). This number was 22 in the top 50 of genes with the highest down-regulation in ΔflbA colonies. Among the top 50 up-regulated and down-regulated genes, 47and 43 genes are not yet characterized, respectively, of which 18 and 23 belong to the family of unclassified proteins. These gene sets also contain genes involved in development and cell wall biosynthesis/ remodelling and genes encoding transcriptional regulators. Up- and down-regulated (≥ 2-fold) genes of these classes [18] were analyzed in more detail and are described below.
Transcriptional changes associated to asexual and sexual development: Of the 68 genes implicated in asexual and/or sexual reproduction in A. niger, 13 are found to be differentially expressed between the wild-type and ΔflbA strain (Figure 4A, Table 5; Supplemental Table 1). Of these genes, 6 are down-regulated in the ΔflbA strain and 7 are up-regulated. Of the genes indicated in Figure 1, flbD and sfaD are ≥ 2-fold higher expressed in the ΔflbA strain.

Figure 4: Genes involved inasexual development (A), and cell wall synthesis (B), and genes encoding extracellular carbohydrases (C) and transcription factors (D) that show ≥ 2-fold lower or higher expression between wild-type and ΔflbA colonies.
| Annotation | Gene Name* | Description | wild-type zone 1 | wild-type zone 3 | wild-type zone 5 | ΔflbA zone 1 | ΔflbA zone 3 | ΔflbA zone 5 | Regulation |
| An02g03160 | flbA | strong similarity to developmental regulator flbA - Emericella nidulans | 147 ± 11 | 166 ± 25 | 77 ± 24 | 21 ± 2 | 24 ± 4 | 25 ± 3 | Down |
| An04g05880 | ppoA | strong similarity to linoleate diol synthase - Gaeumannomyces graminis | 1658 ± 245 | 1167 ± 359 | 282 ± 22 | 301 ± 2 | 298 ± 6 | 188 ± 11 | Down |
| An04g06620 | (RAM1/STE16) | similarity to farnesyl-protein transferase beta chain - Homo sapiens | 1125 ± 2 | 1045 ± 31 | 831 ± 197 | 527 ± 31 | 499 ± 42 | 378 ± 4 | Down |
| An04g07400 | sm rosA- A. fum | strong similarity to C6 zinc finger transcription factor PRO1 - Sordaria macrospora[putative sequencing error] | 120 ± 28 | 195 ± 76 | 562 ± 77 | 86 ± 3 | 83 ± 9 | 43 ± 6 | Down |
| An05g00480 | stuA /phd1 | strong similarity to transcription factor involved in differentiation stuA - Aspergillus nidulans | 3804 ± 1205 | 4031 ± 108 | 1721 ± 59 | 1672 ± 63 | 1531 ± 47 | 978 ± 7 | Down |
| An12g01320 | ppoD | strong similarity to linoleate diol synthase - Gaeumannomyces graminis | 1123 ± 612 | 1043 ± 345 | 132 ± 72 | 18 ± 2 | 20 ± 3 | 14 ± 2 | Down |
| An01g03750 | abaA | strong similarity to protein abaA - Aspergillus nidulans | 61 ± 9 | 71 ± 12 | 67 ± 17 | 113 ± 28 | 129 ± 3 | 182 ± 22 | Up |
| An01g04830 | flbD | strong similarity to myb-like DNA binding protein flbD - Aspergillus nidulans | 89 ± 18 | 91 ± 8 | 81 ± 20 | 238 ± 35 | 220 ± 32 | 107 ± 5 | Up |
| An02g07930 | ppoC | strong similarity to linoleate diol synthase precursor - Gaeumannomyces graminis | 180 ± 34 | 126 ± 52 | 38 ± 2 | 983 ± 279 | 747 ± 19 | 464 ± 32 | Up |
| An14g01820 | phiA /binB | strong similarity to hypothetical cell wall protein binB - Aspergillus nidulans | 2018 ± 1048 | 3064 ± 430 | 843 ± 70 | 8800 ± 3676 | 14346 ± 1791 | 4066 ± 653 | Up |
| An15g02740 | apsA | strong similarity to the anucleate primary sterigmata gene apsA - Aspergillus nidulans | 199 ± 47 | 206 ± 53 | 252 ± 49 | 453 ± 29 | 444 ± 68 | 560 ± 22 | Up |
| An16g01860 | (STE23) | strong similarity to protease involved in a-factor processing STE23 - Saccharomyces cerevisiae | 99 ± 20 | 95 ± 10 | 118 ± 8 | 192 ± 8 | 215 ± 7 | 279 ± 28 | Up |
| An18g02090 | sfaD (STE 4) | strong similarity to G-protein beta subunit sfaD - Aspergillus nidulans | 41 ± 10 | 43 ± 8 | 104 ± 4 | 109 ± 5 | 113 ± 4 | 165 ± 13 | Up |
Table 5: Hybridization values of 13 genes that areimplicated in asexual and/or sexual reproduction and that are differentially expressed in wild-type when compared to the ΔflbA strain ofA. niger. The table is based on Pel et al. [18], with the addition of flbE (An08g07210), flbB (An15g03710) and flbC (An12g08230). *Gene name or its closest homologue. Sm: Similar; A. fum: A. fumigatus.
Transcriptional changes associated to cell wall synthesis: A total of 102 genes are predicted to be involved in cell wall synthesis [18]. Of these genes, 10 and 2 were found to be up- and down-regulated in the ΔflbA strain when compared to the wild-type, respectively (Figure 4B, Table 6, Supplemental Table 1). Among the up-regulated genes are three glucanosyl transferase genes, two chitin synthase genes, the chitinase gene chiB, two glucanase genes, and two glycosylphosphatidylinositolanchored endo-mannanase genes. One of these glucanosyl transferases, a gelD homologue of A. nidulans was 48-fold up-regulated in ΔflbA compared to wild-type. In contrast, the annotated gelD gene of A. niger was found to be 40-fold down-regulated in ΔflbA. The other downregulated gene in the ΔflbA strain is theglucan beta-1,3 exoglucanase gene exsG.
| Annotation | Gene Name* | Description | wild-type zone 1 | wild-type zone 3 | wild-type zone 5 | ΔflbA zone 1 | ΔflbA zone 3 | ΔflbA zone 5 | Regulation |
| An01g12450 | exsG | strong similarity to putative glucan beta-1,3 exoglucanase - Trichoderma harzianum | 3091 ± 197 | 3667 ± 1573 | 1008 ± 250 | 966 ± 4 | 1053 ± 14 | 689 ± 27 | Down |
| An09g00670 | gelD | strong similarity to beta (1-3) glucanosyltransferase Gel3p - Aspergillus fumigatus | 1596 ± 565 | 2244 ± 607 | 2063 ± 821 | 45 ± 1 | 39 ± 6 | 59 ± 10 | Down |
| An02g02660 | dfgG | strong similarity to the protein required for filamentous growth, cell polarity, and cellular elongation Dfg5 - Saccharomyces cerevisiae | 87 ± 5 | 110 ± 17 | 102 ± 21 | 487 ± 30 | 648 ± 129 | 532 ± 98 | Up |
| An03g06220 | sm to gelD- A. nid | strong similarity to beta (1-3) glucanosyltransferase GEL3 - Aspergillus fumigatus | 42 ± 3 | 48 ± 11 | 58 ± 3 | 1406 ± 256 | 1874 ± 54 | 5043 ± 547 | Up |
| An04g04670 | sm to chiB- A. nid | strong similarity to chitinase cts1 - Coccidioides immitis | 106 ± 36 | 104 ± 26 | 111 ± 1 | 171 ± 65 | 308 ± 102 | 331 ± 185 | Up |
| An07g04650 | sm to btgC- A. nid | similarity to exo-beta-1,3-glucanase BGL2 - Saccharomyces cerevisiae. | 203 ± 14 | 275 ± 109 | 184 ± 41 | 699 ± 119 | 797 ± 4 | 319 ± 12 | Up |
| An07g07530 | chrB | strong similarity to cell wall protein UTR2 - Saccharomyces cerevisiae | 176 ± 39 | 254 ± 48 | 1452 ± 40 | 1812 ± 115 | 1764 ± 453 | 2719 ± 364 | Up |
| An08g07350 | sm to gelB- A. nid | strong similarity to glycophospholipid-anchored surface glycoprotein GAS1 precursor - Saccharomyces cerevisiae | 226 ± 26 | 257 ± 8 | 363 ± 114 | 674 ± 9 | 839 ± 71 | 1001 ± 69 | Up |
| An09g02290 | sm to chsD- A.nid | strong similarity to chitin synthase chsE - Aspergillus nidulans | 308 ± 12 | 314 ± 57 | 255 ± 23 | 779 ± 15 | 723 ± 100 | 479 ± 18 | Up |
| An12g10380 | chsF | strong similarity to chitin synthase C chsC - Aspergillus fumigatus | 118 ± 6 | 137 ± 13 | 478 ± 239 | 468 ± 12 | 527 ± 142 | 1407 ± 9 | Up |
| An16g07040 | sm to btgE -A. nid | similarity to beta-1,3-glucanosyltransferase BGT1 - Aspergillus fumigatus[truncated ORF] | 502 ± 224 | 676 ± 353 | 2375 ± 144 | 5982 ± 202 | 6554 ± 1346 | 7358 ± 380 | Up |
| An16g08090 | dfgE | strong similarity to hypothetical protein B2J23.120 - Neurospora crassa | 114 ± 8 | 125 ± 5 | 174 ± 10 | 229 ± 21 | 280 ± 11 | 438 ± 28 | Up |
Table 6: Hybridization values of 12 genes that are implicated in cell wall biosynthesis and that are differentially expressed in wild-type when compared to the ΔflbA strain of A. niger. The table was adapted from Pel et al. [18]. *Description of gene name or its closest homologue. Sm: Similar; A. nid: A. nidulans.
Transcriptional changes associated to proteins with a signal sequence for secretion: As mentioned above, wild-type and ΔflbA were grown on minimal medium supplemented with xylose. This carbon source activates XlnR, a transcriptional regulator that controls 16 genes encoding xylanolytic enzymes [19-25]. Of these, none were higher expressed in the ΔflbA strain, whereas 6 (xlnB, xlnC, axhA, aglB, eglB, cbhA, chbB) were found to be ≥ 2-fold lower expressed when compared to the wild-type (Figure 4C, Table 7, Supplemental Table 1). In fact, xlnC, xlnB, and axhA are among the highest expressed genes at the periphery of wild-type colonies, while they are hardly expressed in the ΔflbA strain (Table 4). Thirty-two other carbohydrase genes are down-regulated in the ΔflbA strain (e.g. the glucoamylase gene glaA), while 8 of these genes are up-regulated in this strain. Among these genes are the α-galactosidase gene aglC, the α-glucan synthase agsE, the pectin lysase pelB and the pectin esterase pmeA (Figure 4C, Table 7).
| Annotation | Gene Name* | Description | wild-type zone 1 | wild-type zone 3 | wild-type zone 5 | ΔflbA zone 1 | ΔflbA zone 3 | ΔflbA zone 5 | Regulation |
| An01g00780 | xlnB | xylanase xynB of patent WO9713853-A2 - Aspergillus niger | 4528 ± 1905 | 16891 ± 683 | 19085 ± 0 | 953 ± 977 | 4014 ± 2052 | 1371 ± 235 | Down |
| An01g01870 | eglC | strong similarity to hypothetical Avicelase III aviIII - Aspergillus aculeatus | 77 ± 47 | 501 ± 267 | 400 ± 240 | 22 ± 0 | 23 ± 1 | 23 ± 2 | Down |
| An01g03340 | sm to celA- A.ory | strong similarity to xyloglucan-specific endo-beta-1,4-glucanase - Aspergillus aculeatus | 552 ± 60 | 4966 ± 831 | 3065 ± 537 | 151 ± 48 | 272 ± 34 | 191 ± 34 | Down |
| An01g06120 | gdbA | strong similarity to 4-alpha-glucanotransferase / amylo-1,6-glucosidase GDB1 - Saccharomyces cerevisiae | 1138 ± 28 | 1366 ± 280 | 915 ± 411 | 446 ± 27 | 385 ± 41 | 290 ± 12 | Down |
| An01g11660 | cbhB | 1,4-beta-D-glucan cellobiohydrolase B precursor cbhB of patent WO9906574-A1 - Aspergillus niger | 1289 ± 335 | 8224 ± 845 | 6037 ± 1558 | 48 ± 13 | 430 ± 386 | 1098 ± 313 | Down |
| An01g11670 | eglA | strong similarity to endo-beta-1,4-glucanase A eglA - Emericella nidulans | 638 ± 408 | 2367 ± 12 | 589 ± 232 | 21 ± 2 | 42 ± 24 | 58 ± 1 | Down |
| An01g14600 | uncharacterized | strong similarity to the endo-1,4-beta-Xylanase B XynB, patent WO9414965 - Aspergillus tubingensis | 256 ± 239 | 364 ± 349 | 195 ± 92 | 30 ± 1 | 34 ± 6 | 37 ± 4 | Down |
| An02g11150 | aglB | alpha-galactosidase aglB - Aspergillus niger | 390 ± 96 | 2373 ± 668 | 3155 ± 569 | 111 ± 5 | 160 ± 21 | 126 ± 14 | Down |
| An02g13240 | agdC | strong similarity to alpha-1-6-glucosidase glcA - Aspergillus parasiticus | 806 ± 223 | 877 ± 435 | 313 ± 147 | 224 ± 5 | 144 ± 26 | 77 ± 7 | Down |
| An03g00940 | xlnC | endo-1,4-beta-xylanase C precursor xlnC - Aspergillus niger | 2605 ± 1941 | 15761 ± 4701 | 15538 ± 1230 | 337 ± 232 | 1058 ± 553 | 1067 ± 324 | Down |
| An03g00960 | axhA | 1,4-beta-D-arabinoxylan arabinofuranohydrolase axhA - Aspergillus niger | 4089 ± 1838 | 15097 ± 1853 | 17374 ± 0 | 456 ± 313 | 1314 ± 776 | 1602 ± 429 | Down |
| An03g01050 | Uncharacterized | similarity to endo-beta-1,4-glucanase - Bacillus polymyxa | 137 ± 32 | 345 ± 202 | 1027 ± 608 | 31 ± 8 | 48 ± 17 | 89 ± 12 | Down |
| An03g03740 | bgl4 | strong similarity to beta-glucosidase bgl4 - Humicola grisea var. thermoidea | 656 ± 278 | 958 ± 22 | 587 ± 133 | 359 ± 34 | 367 ± 72 | 312 ± 13 | Down |
| An03g06550 | glaA | glucan 1,4-alpha-glucosidase glaA - Aspergillus niger | 370 ± 102 | 1497 ± 1144 | 5690 ± 1251 | 182 ± 15 | 171 ± 29 | 166 ± 8 | Down |
| An04g06920 | agdA | extracellular alpha-glucosidase aglU - Aspergillus niger | 236 ± 29 | 316 ± 16 | 651 ± 135 | 176 ± 8 | 145 ± 5 | 176 ± 10 | Down |
| An04g09360 | uncharacterized | strong similarity to hypothetical protein CC0812 - Caulobacter crescentus | 136 ± 33 | 153 ± 76 | 90 ± 13 | 68 ± 0 | 64 ± 1 | 38 ± 8 | Down |
| An04g09690 | sm to pmeA- A. nid | strong similarity to pectin methylesterase PME1 - Aspergillus aculeatus | 60 ± 19 | 190 ± 41 | 174 ± 59 | 38 ± 3 | 48 ± 0 | 47 ± 9 | Down |
| An04g09700 | uncharacterized | strong similarity to endo-xylogalacturonan hydrolase xghA - Aspergillus tubingensis | 60 ± 41 | 435 ± 272 | 143 ± 33 | 28 ± 4 | 28 ± 0 | 39 ± 3 | Down |
| An05g02410 | uncharacterized | strong similarity to beta-glucuronidase GUSB - Canis familiaris | 287 ± 18 | 343 ± 48 | 338 ± 149 | 201 ± 2 | 132 ± 4 | 104 ± 8 | Down |
| An06g00170 | aglA | alpha-galactosidase aglA - Aspergillus niger | 600 ± 56 | 678 ± 85 | 124 ± 23 | 59 ± 7 | 32 ± 6 | 18 ± 3 | Down |
| An07g08950 | eglC | endoglucanase B eglB - Aspergillus niger | 2511 ± 570 | 11775 ± 532 | 7793 ± 1370 | 52 ± 29 | 366 ± 244 | 422 ± 77 | Down |
| An07g09330 | cbhA | cellulose 1,4-beta-cellobiosidase cbhA from patent WO9906574-A1- Aspergillus niger | 1157 ± 1121 | 7883 ± 2929 | 3034 ± 1406 | 67 ± 16 | 130 ± 62 | 170 ± 76 | Down |
| An08g05230 | uncharacterized | strong similarity to putative endoglucanase IV - Trichoderma reesei | 1152 ± 1281 | 2411 ± 2729 | 648 ± 116 | 55 ± 5 | 58 ± 11 | 46 ± 5 | Down |
| An08g10780 | uncharacterized | strong similarity to hypothetical protein T16K5.230 of A. thaliana | 72 ± 40 | 140 ± 70 | 1109 ± 145 | 30 ± 8 | 26 ± 2 | 43 ± 0 | Down |
| An09g02160 | rgaeA | rhamnogalacturonan acetyl esterase rgaeA - Aspergillus niger | 318 ± 9 | 829 ± 168 | 409 ± 176 | 165 ± 41 | 149 ± 17 | 58 ± 2 | Down |
| An11g02100 | sm to bglR- A. nid | strong similarity to furostanol glycoside 26-O-beta-glucosidase CSF26G1 - Costus speciosus | 360 ± 21 | 1634 ± 301 | 808 ± 253 | 105 ± 38 | 140 ± 12 | 114 ± 5 | Down |
| An12g04610 | uncharacterized | similarity to endoglucanase IV egl4 - Trichoderma reesei | 966 ± 1072 | 5636 ± 3072 | 243 ± 11 | 76 ± 9 | 108 ± 31 | 58 ± 5 | Down |
| An13g03710 | agdD | strong similarity to alpha-glucosidase AGLU - Bacillus sp. SAM1606 | 103 ± 18 | 130 ± 57 | 77 ± 28 | 44 ± 4 | 42 ± 4 | 47 ± 3 | Down |
| An14g01800 | aglD | alpha-galactosidase aglD - Aspergillus niger | 54 ± 4 | 109 ± 6 | 136 ± 18 | 27 ± 0 | 33 ± 3 | 34 ± 2 | Down |
| An14g02670 | uncharacterized | strong similarity to endoglucanase IV egl4 - Trichoderma reesei | 347 ± 273 | 857 ± 754 | 737 ± 10 | 37 ± 3 | 45 ± 3 | 37 ± 1 | Down |
| An14g04190 | gbeA | strong similarity to 1,4-alpha-glucan branching enzyme glc3 - Saccharomyces cerevisiae | 1445 ± 19 | 1453 ± 195 | 915 ± 221 | 769 ± 3 | 603 ± 58 | 451 ± 20 | Down |
| An14g04200 | rhgB | rhamnogalacturonase rhgB - Aspergillus niger | 83 ± 57 | 177 ± 184 | 105 ± 34 | 33 ± 7 | 36 ± 5 | 34 ± 0 | Down |
| An14g05820 | uncharacterized | strong similarity to beta-galactosidase lacA - Aspergillus niger | 126 ± 37 | 290 ± 21 | 275 ± 6 | 60 ± 13 | 62 ± 12 | 72 ± 12 | Down |
| An15g04550 | xynA | strong similarity to xylanase A xynA of patent WO200068396-A2 - Aspergillus niger | 55 ± 0 | 384 ± 304 | 1176 ± 75 | 35 ± 4 | 41 ± 21 | 34 ± 1 | Down |
| An15g05370 | pgaII | polygalacturonase pgaII of patent EP421919-A - Aspergillus niger | 224 ± 219 | 767 ± 877 | 327 ± 240 | 29 ± 3 | 25 ± 5 | 28 ± 5 | Down |
| An16g00540 | uncharacterized | similarity to putative large secreted protein - Streptomyces coelicolor[truncated ORF] | 32 ± 1 | 89 ± 3 | 159 ± 14 | 23 ± 1 | 25 ± 1 | 26 ± 6 | Down |
| An16g06800 | eglB | strong similarity to endoglucanase eglB - Aspergillus niger | 154 ± 25 | 226 ± 100 | 423 ± 232 | 87 ± 5 | 92 ± 5 | 77 ± 5 | Down |
| An17g00300 | xarB | strong similarity to bifunctiona xylosidase-arabinosidase xarB - Thermoanaerobacter ethanolicus | 156 ± 10 | 534 ± 22 | 396 ± 19 | 88 ± 6 | 124 ± 19 | 144 ± 17 | Down |
| An01g01540 | uncharacterized | strong similarity to alpha,alpha-trehalase treA - Aspergillus nidulans | 154 ± 10 | 161 ± 8 | 149 ± 13 | 568 ± 30 | 607 ± 43 | 366 ± 5 | Up |
| An01g10350 | sm to lacB- A. niger | strong similarity to secreted beta-galactosidase lacA - Aspergillus niger | 113 ± 5 | 109 ± 20 | 206 ± 35 | 469 ± 36 | 394 ± 38 | 499 ± 69 | Up |
| An03g00190 | pelB | the pectin lyase pelB - Aspergillus niger | 56 ± 4 | 48 ± 4 | 61 ± 9 | 338 ± 67 | 536 ± 67 | 248 ± 3 | Up |
| An03g06310 | pmeA | pectinesterase pmeA- Aspergillus niger | 33 ± 6 | 36 ± 6 | 43 ± 5 | 65 ± 4 | 75 ± 14 | 179 ± 41 | Up |
| An09g00260 | aglC | alpha-galactosidase C aglC - Aspergillus niger[truncated ORF] | 31 ± 6 | 34 ± 4 | 143 ± 60 | 208 ± 5 | 216 ± 38 | 370 ± 31 | Up |
| An09g00270 | aglC | alpha-galactosidase C aglC - Aspergillus niger[truncated ORF] | 89 ± 18 | 85 ± 22 | 760 ± 428 | 879 ± 73 | 1158 ± 119 | 1805 ± 251 | Up |
| An09g03070 | agsE | strong similarity to alpha-glucan synthase mok1 - Schizosaccharomyces pombe | 174 ± 2 | 142 ± 40 | 163 ± 50 | 544 ± 44 | 598 ± 19 | 458 ± 41 | Up |
| An15g03550 | uncharacterized | weak similarity to protopectinase patent WO9806832-A1 - Bacillus subtilis | 57 ± 44 | 57 ± 45 | 20 ± 0 | 954 ± 34 | 1066 ± 21 | 330 ± 22 | Up |
Table 7: Hybridization values of 46 genes that are implicated in carbohydrate degradationand that are differentially expressed in wild-type when compared to the ΔflbA strain of A. niger. The table was adapted from Pel et al. [18]. *Description of gene name or its closest homologue. Sm: Similar; A. nid: A. nidulans; A. ory: A. oryzae.
Of the total number of 2612 genes with a predicted signal sequence for secretion, 156 and 189 were up- and down-regulated, respectively (Supplemental Figure 2, Supplemental Table 1and 5). The up-regulated genes are mainly uncharacterized proteins, but include carbohydrases (see above), a putative cytochrome P450 reductase and two putative proteases. Among the down-regulated are several carbohydrate degrading enzymes (Supplemental Table 1 and 5), a phytase (phyB) and six (putative) proteases.
Previously, we identified 138 proteins with a signal sequence for secretion in the secretome of ΔflbA colonies when grown on fresh xylose medium [8]. Of these, 70 were not found in the wild-type secretome (i.e. even after partial degradation of the cell wall by the addition of cycloheximide) [8]. Differential expression was observed in the case of 23 out of 70 encoding genes (Table 8). One of these genes was down-regulated, whereas 22 were up-regulated in the ΔflbA colonies. Most of these genes are uncharacterized, but encode proteins that are putative carbohydrate degrading enzymes (3), proteases (2), cell wall/ morphogenesis enzymes (2), lipases (2), phospholipases (2), oxidases and superoxide-dismutases (5) and other/unknown proteins (6) (Table 8). Futhermore, 2 and 5 genes were up- and downregulated, respectively, of the genes encoding 30 proteins of the ΔflbA secretome [8] that were absent in the wild-type secretome [7], and that are predicted to be secreted via non-classical secretion (15) (SecP), or have no prediction for secretion (15) [8] (Table 8). The secretome of the cycloheximide treated wild-type strain contained 55 proteins [7] that were absent in the ΔflbA secretome [8], of which 35 encoding genes were differentially expressed. The two up-regulated genes in the ΔflbA strain encode uncharacterized proteins. The 33 down-regulated genes encode carbohydrate degrading enzymes (17), phytases (2), proteases (5), oxidase (1) and other/unknown proteins (5) (Table 8). Of the 37 proteins that were identified in both the wild-type and ΔflbA secretome, 6 and 14 encoding genes were found to be 2-fold higher and lower expressed in the ΔflbA strain, respectively (Table 8). The up-regulated genes consist of putative cell wall/ morphogenesis enzymes (3), carbohydrate degrading enzymes (1), a protease (1), and a superoxide dismutase (1), while the down-regulated genes encode putative carbohydrate degrading enzymes (11), a cell wall/ morphogenesis enzyme (1), a phospholipase (1) and a lipase (1).
| Annotation | Gene Name* | Description | wild-type zone 1 | wild-type zone 3 | wild-type zone 5 | ΔflbA zone 1 | ΔflbA zone 3 | ΔflbA zone 5 | Regulation |
| proteinproducts identified in the secretome of ΔflbAthat are predicted to be secreted | |||||||||
| An11g06480 | uncharacterized | weak similarity to antigenic protein f86.aa. of patent WO9859071 - Borrelia burgdorferi | 165 ± 5 | 154 ± 2 | 119 ± 6 | 43 ± 9 | 38 ± 1 | 43 ± 4 | Down |
| An09g00260 | aglC | alpha-galactosidase C aglC - Aspergillus niger[truncated ORF] | 31 ± 6 | 34 ± 4 | 143 ± 60 | 208 ± 5 | 216 ± 38 | 370 ± 31 | Up |
| An03g06310 | pmeA | pectinesterase pmeA- Aspergillus niger | 33 ± 6 | 36 ± 6 | 43 ± 5 | 65 ± 4 | 75 ± 14 | 179 ± 41 | Up |
| An15g03550 | uncharacterized | weak similarity to protopectinase patent WO9806832-A1 - Bacillus subtilis | 57 ± 44 | 57 ± 45 | 20 ± 0 | 954 ± 34 | 1066 ± 21 | 330 ± 22 | Up |
| An11g00100 | uncharacterized | strong similarity to triacylglycerol lipase LIP5 - Candida rugosa | 71 ± 4 | 64 ± 3 | 59 ± 8 | 800 ± 192 | 866 ± 397 | 2265 ± 77 | Up |
| An16g08870 | uncharacterized | strong similarity to the triacylglycerol lipase I precursor lipI - Geotrichum candidum | 139 ± 16 | 125 ± 28 | 84 ± 2 | 395 ± 28 | 267 ± 13 | 181 ± 12 | Up |
| An09g01240 | uncharacterized | strong similarity to phospholipase B - Penicillium notatum | 78 ± 7 | 63 ± 9 | 104 ± 3 | 499 ± 142 | 727 ± 111 | 627 ± 178 | Up |
| An16g01880 | lipanl | strong similarity to lysophospholipase - Aspergillus foetidus | 648 ± 245 | 795 ± 248 | 454 ± 202 | 10644 ± 663 | 10585 ± 2214 | 2676 ± 160 | Up |
| An02g00740 | uncharacterized | similarity to 6-Hydroxy-D-nicotine oxidase 6-HDNO - Arthrobacter oxidans | 31 ± 1 | 30 ± 2 | 35 ± 3 | 61 ± 5 | 69 ± 11 | 130 ± 14 | Up |
| An02g08560 | uncharacterized | similarity to probable dioxygenase SCOEDB - Streptomyces coelicolor | 22 ± 3 | 23 ± 3 | 19 ± 0 | 797 ± 22 | 1048 ± 83 | 1682 ± 135 | Up |
| An03g00460 | uncharacterized | strong similarity to the 6-hydroxy-D-nicotine oxidase 6-HDNO - Arthrobacter oxidans | 129 ± 69 | 90 ± 27 | 102 ± 7 | 913 ± 19 | 751 ± 8 | 198 ± 2 | Up |
| An03g05210 | uncharacterized | strong similarity to reticuline oxidase bbe1 - Eschscholzia californica | 146 ± 47 | 180 ± 82 | 308 ± 23 | 423 ± 2 | 453 ± 23 | 767 ± 34 | Up |
| An06g00720 | uncharacterized | similarity to chloroperoxidase CPO - Caldariomyces fumago | 59 ± 24 | 59 ± 16 | 40 ± 14 | 109 ± 12 | 134 ± 32 | 109 ± 2 | Up |
| An03g06220 | sm to gelD- A. nid | strong similarity to beta (1-3) glucanosyltransferase GEL3 - Aspergillus fumigatus | 42 ± 3 | 48 ± 11 | 58 ± 3 | 1406 ± 256 | 1874 ± 54 | 5043 ± 547 | Up |
| An16g07040 | sm to btgE -A. nid | similarity to beta-1,3-glucanosyltransferase BGT1 - Aspergillus fumigatus[truncated ORF] | 502 ± 224 | 676 ± 353 | 2375 ± 144 | 5982 ± 202 | 6554 ± 1346 | 7358 ± 380 | Up |
| An05g02170 | uncharacterized | strong similarity to serine-type carboxypeptidase F CPD-II - Aspergillus niger | 60 ± 17 | 57 ± 6 | 70 ± 11 | 109 ± 12 | 121 ± 19 | 240 ± 17 | |
| An18g01320 | uncharacterized | strong similarity to extracellular protease precursor BAR1 - Saccharomyces cerevisiae | 148 ± 76 | 129 ± 56 | 328 ± 31 | 2146 ± 67 | 2897 ± 61 | 2501 ± 161 | Up |
| An01g06280 | uncharacterized | strong similarity to IgE-binding protein - Aspergillus fumigatus | 267 ± 123 | 341 ± 167 | 136 ± 4 | 2010 ± 28 | 2634 ± 280 | 1002 ± 144 | Up |
| An03g00770 | uncharacterized | strong similarity to allergic bronchopulmonary aspergillosis allergen rAsp f 4 of patent WO9828624-A1 - Aspergillus fumigatus | 93 ± 3 | 98 ± 34 | 454 ± 19 | 473 ± 118 | 437 ± 60 | 2858 ± 7 | Up |
| An04g07160 | uncharacterized | similarity to hypothetical protein MLD14.3 - Arabidopsis thaliana | 202 ± 34 | 215 ± 74 | 246 ± 38 | 548 ± 65 | 656 ± 82 | 666 ± 81 | Up |
| An06g00160 | uncharacterized | weak similarity to hypothetical cell wall protein binB - Aspergillus nidulans | 319 ± 29 | 330 ± 70 | 128 ± 21 | 3226 ± 732 | 2940 ± 1230 | 453 ± 184 | Up |
| An07g02730 | uncharacterized | strong similarity to SUN family protein Psu1 - Schizosaccharomyces pombe | 261 ± 96 | 317 ± 133 | 1534 ± 12 | 2045 ± 210 | 2424 ± 353 | 3717 ± 85 | Up |
| An09g03650 | uncharacterized | weak similarity to hypothetical protein Ta0309 - Thermoplasma acidophilum | 84 ± 40 | 52 ± 1 | 133 ± 16 | 375 ± 55 | 488 ± 99 | 438 ± 1 | Up |
| protein products identified in the ΔflbA secreteome that are predicted to be released by non-classical secretion | |||||||||
| An09g02830 | uncharacterized | strong similarity to acylaminoacyl-peptidase DPP V - Aspergillus fumigatus | 385 ± 95 | 420 ± 131 | 622 ± 25 | 251 ± 45 | 224 ± 14 | 116 ± 2 | |
| An09g00270 | aglC | alpha-galactosidase C aglC - Aspergillus niger[truncated ORF] | 89 ± 18 | 85 ± 22 | 760 ± 428 | 879 ± 73 | 1158 ± 119 | 1805 ± 251 | Up |
| protein products identified in the ΔflbA secretome without signal sequence for secretion | |||||||||
| An01g06970 | ara1 | strong similarity to D-arabinose dehydrogenase ARA1 - Saccharomyces cerevisiae | 2370 ± 516 | 3061 ± 357 | 2476 ± 456 | 643 ± 50 | 627 ± 28 | 1838 ± 51 | |
| An02g11970 | uncharacterized | strong similarity to 4-nitrophenylphosphatase pho2 - Schizosaccharomyces pombe | 85 ± 12 | 93 ± 21 | 123 ± 9 | 164 ± 6 | 183 ± 36 | 416 ± 16 | Up |
| An08g10830 | uncharacterized | strong similarity to geranylgeranyl pyrophosphate synthase ggpps - Gibberella fujikuroi | 65 ± 5 | 58 ± 0 | 68 ± 18 | 412 ± 9 | 388 ± 31 | 555 ± 30 | Up |
| An09g06250 | uncharacterized | strong similarity to vacuolar aminopeptidase yscI - Saccharomyces cerevisiae | 184 ± 42 | 187 ± 30 | 405 ± 70 | 526 ± 16 | 555 ± 41 | 430 ± 39 | Up |
| An12g08610 | glkA | glucokinase GlkA - Aspergillus niger | 302 ± 29 | 324 ± 114 | 306 ± 96 | 652 ± 16 | 622 ± 75 | 660 ± 65 | |
| protein products identified in wild-type and ΔflbA secretome that are predicted to be secreted | |||||||||
| An02g11150 | aglB | alpha-galactosidase aglB - Aspergillus niger | 390 ± 96 | 2373 ± 668 | 3155 ± 569 | 111 ± 5 | 160 ± 21 | 126 ± 14 | Down |
| An01g00780 | xlnB | endo-1,4-beta-xylanase B precursor xlnB - Aspergillus niger | 4528 ± 1905 | 16891 ± 683 | 19085 ± 0 | 953 ± 977 | 4014 ± 2052 | 1371 ± 235 | Down |
| An03g00940 | xlnC | endo-1,4-beta-xylanase C precursor xlnC - Aspergillus niger | 2605 ± 1941 | 15761 ± 4701 | 15538 ± 1230 | 337 ± 232 | 1058 ± 553 | 1067 ± 324 | Down |
| An03g00960 | axhA | 1,4-beta-D-arabinoxylan arabinofuranohydrolase axhA - Aspergillus niger | 4089 ± 1838 | 15097 ± 1853 | 17374 ± 0 | 456 ± 313 | 1314 ± 776 | 1602 ± 429 | Down |
| An14g01800 | aglD | alpha-galactosidase aglD - Aspergillus niger | 54 ± 4 | 109 ± 6 | 136 ± 18 | 27 ± 0 | 33 ± 3 | 34 ± 2 | Down |
| An17g00300 | xarB | strong similarity to bifunctiona xylosidase-arabinosidase xarB - Thermoanaerobacter ethanolicus | 156 ± 10 | 534 ± 22 | 396 ± 19 | 88 ± 6 | 124 ± 19 | 144 ± 17 | Down |
| An01g11660 | cbhB | 1,4-beta-D-glucan cellobiohydrolase B precursor cbhB of patent WO9906574-A1 - Aspergillus niger | 1289 ± 335 | 8224 ± 845 | 6037 ± 1558 | 48 ± 13 | 430 ± 386 | 1098 ± 313 | Down |
| An07g09330 | cbhA | cellulose 1,4-beta-cellobiosidase cbhA from patent WO9906574-A1- Aspergillus niger | 1157 ± 1121 | 7883 ± 2929 | 3034 ± 1406 | 67 ± 16 | 130 ± 62 | 170 ± 76 | Down |
| An16g06800 | eglB | strong similarity to endoglucanase eglB - Aspergillus niger | 154 ± 25 | 226 ± 100 | 423 ± 232 | 87 ± 5 | 92 ± 5 | 77 ± 5 | Down |
| An08g05230 | uncharacterized | strong similarity to putative endoglucanase IV - Trichoderma reesei | 1152 ± 1281 | 2411 ± 2729 | 648 ± 116 | 55 ± 5 | 58 ± 11 | 46 ± 5 | Down |
| An03g06550 | glaA | glucan 1,4-alpha-glucosidase glaA - Aspergillus niger | 370 ± 102 | 1497 ± 1144 | 5690 ± 1251 | 182 ± 15 | 171 ± 29 | 166 ± 8 | Down |
| An01g12450 | exsG | strong similarity to putative glucan beta-1,3 exoglucanase - Trichoderma harzianum | 3091 ± 197 | 3667 ± 1573 | 1008 ± 250 | 966 ± 4 | 1053 ± 14 | 689 ± 27 | Down |
| An02g09690 | uncharacterized | strong similarity to lipase I precursor TFL I - Geotrichum fermentans[putative sequencing error] | 652 ± 18 | 1719 ± 762 | 529 ± 399 | 195 ± 22 | 294 ± 108 | 356 ± 50 | Down |
| An01g14940 | uncharacterized | similarity to nonhemolytic phospholipase C PC-PLC - Burkholderia pseudomallei | 299 ± 58 | 486 ± 126 | 203 ± 35 | 122 ± 2 | 138 ± 34 | 57 ± 2 | Down |
| An01g01540 | uncharacterized | strong similarity to alpha,alpha-trehalase treA - Aspergillus nidulans | 154 ± 10 | 161 ± 8 | 149 ± 13 | 568 ± 30 | 607 ± 43 | 366 ± 5 | Up |
| An07g07530 | crhB | strong similarity to cell wall protein UTR2 - Saccharomyces cerevisiae | 176 ± 39 | 254 ± 48 | 1452 ± 40 | 1812 ± 115 | 1764 ± 453 | 2719 ± 364 | Up |
| An14g01820 | phiA /binB | strong similarity to hypothetical cell wall protein binB - Aspergillus nidulans | 2018 ± 1048 | 3064 ± 430 | 843 ± 70 | 8800 ± 3676 | 14346 ± 1791 | 4066 ± 653 | Up |
| An08g07350 | sm to gelB- A. nid | strong similarity to glycophospholipid-anchored surface glycoprotein GAS1 precursor - Saccharomyces cerevisiae | 226 ± 26 | 257 ± 8 | 363 ± 114 | 674 ± 9 | 839 ± 71 | 1001 ± 69 | Up |
| An08g00490 | uncharacterized | strong similarity to lactonohydrolase - Fusarium oxysporum | 160 ± 14 | 237 ± 17 | 346 ± 177 | 484 ± 34 | 516 ± 99 | 806 ± 14 | Up |
| An14g02470 | uncharacterized | strong similarity to the protein PRO304 of patent WO200104311-A1 - Homo sapiens | 326 ± 177 | 470 ± 61 | 384 ± 107 | 2772 ± 181 | 3044 ± 235 | 770 ± 49 | Up |
| Protein products identified in wild-type secretome that are predicted to be secreted | |||||||||
| An01g14600 | uncharacterized | strong similarity to the endo-1,4-beta-Xylanase B XynB, patent WO9414965 - Aspergillus tubingensis | 256 ± 239 | 364 ± 349 | 195 ± 92 | 30 ± 1 | 34 ± 6 | 37 ± 4 | Down |
| An06g00170 | aglA | alpha-galactosidase aglA - Aspergillus niger | 600 ± 56 | 678 ± 85 | 124 ± 23 | 59 ± 7 | 32 ± 6 | 18 ± 3 | Down |
| An04g06920 | agdA | extracellular alpha-glucosidase aglU - Aspergillus niger | 236 ± 29 | 316 ± 16 | 651 ± 135 | 176 ± 8 | 145 ± 5 | 176 ± 10 | Down |
| An04g09690 | sm to pmeA- A. nid | strong similarity to pectin methylesterase PME1 - Aspergillus aculeatus | 60 ± 19 | 190 ± 41 | 174 ± 59 | 38 ± 3 | 48 ± 0 | 47 ± 9 | Down |
| An07g08940 | uncharacterized | similarity to acetyl-esterase I of patent WO9502689-A - Aspergillus aculeatus | 242 ± 13 | 1268 ± 407 | 914 ± 553 | 20 ± 1 | 31 ± 7 | 26 ± 0 | Down |
| An01g11670 | eglA | strong similarity to endo-beta-1,4-glucanase A eglA - Emericella nidulans | 638 ± 408 | 2367 ± 12 | 589 ± 232 | 21 ± 2 | 42 ± 24 | 58 ± 1 | Down |
| An07g08950 | eglC | endoglucanase B eglB - Aspergillus niger | 2511 ± 570 | 11775 ± 532 | 7793 ± 1370 | 52 ± 29 | 366 ± 244 | 422 ± 77 | Down |
| An01g01870 | eglC | strong similarity to hypothetical Avicelase III aviIII - Aspergillus aculeatus | 77 ± 47 | 501 ± 267 | 400 ± 240 | 22 ± 0 | 23 ± 1 | 23 ± 2 | Down |
| An03g01050 | uncharacterized | similarity to endo-beta-1,4-glucanase - Bacillus polymyxa | 137 ± 32 | 345 ± 202 | 1027 ± 608 | 31 ± 8 | 48 ± 17 | 89 ± 12 | Down |
| An08g10780 | uncharacterized | strong similarity to hypothetical protein T16K5.230 of A. thaliana | 72 ± 40 | 140 ± 70 | 1109 ± 145 | 30 ± 8 | 26 ± 2 | 43 ± 0 | Down |
| An14g02670 | uncharacterized | strong similarity to endoglucanase IV egl4 - Trichoderma reesei | 347 ± 273 | 857 ± 754 | 737 ± 10 | 37 ± 3 | 45 ± 3 | 37 ± 1 | Down |
| An08g08370 | uncharacterized | similarity to the alpha-1,2-mannosidase aman2 - Bacillus sp. M-90 | 1771 ± 843 | 2086 ± 211 | 376 ± 19 | 179 ± 33 | 78 ± 14 | 38 ± 4 | Down |
| An16g00540 | uncharacterized | similarity to putative large secreted protein - Streptomyces coelicolor[truncated ORF] | 32 ± 1 | 89 ± 3 | 159 ± 14 | 23 ± 1 | 25 ± 1 | 26 ± 6 | Down |
| An02g02540 | uncharacterized | strong similarity to acetyl-esterase I from patent R63066 - Aspergillus aculeatus | 296 ± 66 | 375 ± 174 | 350 ± 139 | 83 ± 16 | 94 ± 1 | 74 ± 12 | Down |
| An04g09700 | uncharacterized | strong similarity to endo-xylogalacturonan hydrolase xghA - Aspergillus tubingensis | 60 ± 41 | 435 ± 272 | 143 ± 33 | 28 ± 4 | 28 ± 0 | 39 ± 3 | Down |
| An14g04200 | rhgB | rhamnogalacturonase rhgB - Aspergillus niger | 83 ± 57 | 177 ± 184 | 105 ± 34 | 33 ± 7 | 36 ± 5 | 34 ± 0 | Down |
| An15g05370 | pgaII | polygalacturonase pgaII of patent EP421919-A - Aspergillus niger | 224 ± 219 | 767 ± 877 | 327 ± 240 | 29 ± 3 | 25 ± 5 | 28 ± 5 | Down |
| An07g02360 | uncharacterized | similarity to 6-hydroxy-D-nicotine oxidase 6-HDNO - Arthrobacter oxidans | 60 ± 24 | 49 ± 23 | 104 ± 15 | 32 ± 3 | 31 ± 2 | 25 ± 4 | Down |
| An08g09850 | uncharacterized | strong similarity to phosphate-repressible acid phosphatase precursor phoA - Penicillium chrysogenum | 424 ± 125 | 551 ± 364 | 636 ± 431 | 245 ± 15 | 256 ± 8 | 132 ± 7 | Down |
| An08g11030 | phyB | acid phosphatase aph, 3-phytase phyB - Aspergillus niger | 436 ± 438 | 975 ± 1113 | 973 ± 889 | 54 ± 4 | 59 ± 5 | 44 ± 0 | Down |
| An08g04640 | protB | strong similarity to putative lysosomal pepstatin insensitive protease CLN2 - Canis familiaris | 98 ± 32 | 218 ± 127 | 827 ± 628 | 62 ± 13 | 58 ± 5 | 59 ± 2 | Down |
| An16g09010 | protH | strong similarity to carboxypeptidase I protein of patent WO9814599-A1 - Aspergillus oryza | 1018 ± 249 | 1107 ± 340 | 353 ± 147 | 312 ± 2 | 298 ± 3 | 161 ± 9 | Down |
| An06g00190 | uncharacterized | strong similarity to lysosomal pepstatin insensitive protease CLN2 - Homo sapiens | 305 ± 23 | 733 ± 87 | 1175 ± 28 | 144 ± 2 | 114 ± 5 | 58 ± 5 | Down |
| An12g05960 | uncharacterized | strong similarity to dipeptidyl peptidase II DPPII - Rattus norvegicus | 105 ± 22 | 126 ± 40 | 207 ± 38 | 81 ± 22 | 81 ± 4 | 47 ± 7 | Down |
| An14g02150 | uncharacterized | strong similarity to serine-type carboxypeptidase precursor cpdS - Aspergillus phoenicis | 49 ± 10 | 57 ± 12 | 38 ± 7 | 23 ± 2 | 21 ± 1 | 23 ± 4 | Down |
| An09g00670 | geld | strong similarity to beta (1-3) glucanosyltransferase Gel3p - Aspergillus fumigatus | 1596 ± 565 | 2244 ± 607 | 2063 ± 821 | 45 ± 1 | 39 ± 6 | 59 ± 10 | Down |
| An09g00840 | uncharacterized | weak similarity to antigenic cell wall galactomannoprotein MP1 - Aspergillus fumigatus | 3153 ± 2599 | 10725 ± 146 | 3462 ± 103 | 29 ± 11 | 35 ± 22 | 14 ± 2 | Down |
| An01g15200 | uncharacterized | strong similarity to mature penicillin V amidohydrolase PVA of patent US5516679-A - Fusarium oxysporum[truncated ORF] | 1551 ± 24 | 1626 ± 657 | 351 ± 58 | 146 ± 9 | 92 ± 0 | 108 ± 31 | Down |
| An13g01880 | uncharacterized | strong similarity to cephalosporin esterase - Rhodosporidium toruloides | 407 ± 104 | 3000 ± 374 | 3350 ± 147 | 96 ± 37 | 194 ± 69 | 155 ± 2 | Down |
| An01g00210 | uncharacterized | hypothetical protein | 105 ± 13 | 174 ± 37 | 91 ± 31 | 36 ± 2 | 37 ± 0 | 50 ± 8 | Down |
| An02g11890 | uncharacterized | strong similarity to hypothetical protein An14g01330 - Aspergillus niger | 61 ± 14 | 161 ± 33 | 124 ± 11 | 24 ± 1 | 30 ± 1 | 29 ± 2 | Down |
| An08g04630 | uncharacterized | hypothetical protein | 361 ± 349 | 674 ± 706 | 305 ± 86 | 51 ± 1 | 48 ± 13 | 36 ± 1 | Down |
| An15g02250 | uncharacterized | hypothetical protein | 1220 ± 1202 | 1162 ± 905 | 78 ± 37 | 47 ± 2 | 49 ± 6 | 50 ± 9 | Down |
| An06g01000 | uncharacterized | strong similarity to hypothetical protein AN5357.2 - Aspergillus nidulans | 201 ± 49 | 252 ± 68 | 397 ± 160 | 601 ± 188 | 760 ± 179 | 892 ± 307 | Up |
| An11g00040 | uncharacterized | weak similarity to cDNA for 59-kDa readthrough protein RT - Sorghum chlorotic spot virus | 780 ± 377 | 869 ± 597 | 1222 ± 357 | 3463 ± 134 | 3264 ± 0 | 2983 ± 97 | Up |
| Protein products identified in wild-type without signal sequence for secretion | |||||||||
| An14g01790 | uncharacterized | hypothetical protein | 93 ± 2 | 199 ± 15 | 220 ± 21 | 37 ± 4 | 54 ± 8 | 47 ± 16 | Down |
| An01g00610 | uncharacterized | weak similarity to hypothetical protein yukJ - Bacillus subtilis | 455 ± 238 | 550 ± 223 | 1226 ± 644 | 47 ± 3 | 40 ± 6 | 73 ± 6 | Down |
Table 8: Hybridization values of 87 differentially regulated genes whose protein products were identified in the secretome of xylose-grown sandwiched colonies of the ΔflbA
[8] strain and/or the wild-type [7] of A. niger. *Description of gene name or its closest homologue. Sm: Similar; A. nid: A. nidulans.
Transcriptional changes associated to transcription factors: Pel et al. [18] described 623 A. niger transcription factors, of which 20 and 18 are found to be up- and down-regulated in the ΔflbA strain, respectively (Figure 4D, Table 9, Supplemental Table 1). Of these genes, 28 encode transcription factors with an unknown function, while 3 are homologous to transcription factor genes of A. oryzae and 1 to A. fumigatus. These orthologues encode atfB (An14g06250), xlnR (An11g06290), amyR (An01g06900) and rosA (An04g07400), respectively. Moreover, the regulator of extracellular protease prtT was found to be down-regulated in the ΔflbA strain. In contrast, pacC that is activated upon exposure to alkaline pH and acuB that activates acetate metabolism are ≥ 2-fold higher expressed in the ΔflbA strain. Similarly, abaA and flbD, both involved in reproductive development (see above), are upregulated in ΔflbA.
| Annotation | Gene Name* | Description | wild-type | wild-type | wild-type | ΔflbA | ΔflbA | ΔflbA | Regulation |
| zone 1 | zone 3 | zone 5 | zone 1 | zone 3 | zone 5 | ||||
| An04g08620 | uncharacterized | similarity to heme activator protein HAP1 - Saccharomyces cerevisiae | 1079 ± 14 | 1118 ± 11 | 942 ± 47 | 642 ± 42 | 512 ± 28 | 191 ± 21 | Down |
| An12g01870 | uncharacterized | similarity to positive regulator of the lactose-galactose regulon LAC9 - Kluyveromyces lactis | 495 ± 135 | 1003 ± 137 | 720 ± 305 | 254 ± 24 | 271 ± 9 | 167 ± 16 | Down |
| An16g08800 | uncharacterized | similarity to the zinc finger transcription factor ACEI - Trichoderma reesei | 543 ± 121 | 642 ± 119 | 310 ± 34 | 77 ± 17 | 78 ± 5 | 77 ± 1 | Down |
| An15g00120 | uncharacterized | similarity to Wilms tumor susceptibility protein WT1 - Homo sapiens | 476 ± 307 | 442 ± 230 | 580 ± 259 | 112 ± 11 | 111 ± 1 | 65 ± 3 | Down |
| An04g05060 | uncharacterized | similarity to XFIN protein - Xenopus laevis | 3584 ± 911 | 3946 ± 117 | 1943 ± 688 | 1248 ± 6 | 1042 ± 73 | 810 ± 91 | Down |
| An04g07400 | sm rosA- A. fum | strong similarity to C6 zinc finger transcription factor PRO1 - Sordaria macrospora[putative sequencing error] | 120 ± 28 | 195 ± 76 | 562 ± 77 | 86 ± 3 | 83 ± 9 | 43 ± 6 | Down |
| An13g01370 | uncharacterized | strong similarity to the hypothetical protein An01g14540 - Aspergillus niger | 568 ± 230 | 603 ± 155 | 359 ± 47 | 207 ± 8 | 157 ± 8 | 192 ± 26 | Down |
| An11g07610 | uncharacterized | strong similarity to the hypothetical protein encoded by An15g04740 - Aspergillus niger | 226 ± 129 | 212 ± 48 | 104 ± 38 | 54 ± 1 | 43 ± 2 | 33 ± 0 | Down |
| An11g06290 | sm to xlnR -A. ory | strong similarity to transcriptional activator xlnR - Aspergillus niger | 145 ± 30 | 141 ± 35 | 84 ± 17 | 54 ± 2 | 40 ± 4 | 30 ± 1 | Down |
| An01g13790 | uncharacterized | weak similarity to homeodomain protein Prep-1 - Homo sapiens | 792 ± 126 | 685 ± 218 | 368 ± 78 | 220 ± 10 | 203 ± 29 | 317 ± 8 | Down |
| An12g02880 | uncharacterized | weak similarity to hypothetical protein SPBC530.05 - Schizosaccharomyces pombe | 80 ± 51 | 80 ± 48 | 39 ± 1 | 24 ± 1 | 27 ± 0 | 31 ± 0 | Down |
| An06g00830 | uncharacterized | weak similarity to hypothetical transcription regulatory protein SPBC530.08 - Schizosaccharomyces pombe | 121 ± 26 | 146 ± 14 | 221 ± 35 | 55 ± 6 | 67 ± 22 | 49 ± 8 | Down |
| An08g03770 | uncharacterized | weak similarity to mucin MUC5AC - Homo sapiens | 85 ± 33 | 77 ± 21 | 132 ± 63 | 23 ± 2 | 19 ± 3 | 18 ± 2 | Down |
| An07g07370 | uncharacterized | weak similarity to PC-MYB2 - Arabidopsis thaliana | 2629 ± 9 | 2410 ± 318 | 1104 ± 502 | 724 ± 44 | 649 ± 44 | 253 ± 2 | Down |
| An04g01430 | uncharacterized | weak similarity to the chitinase ChiA - Emericella nidulans | 14247 ± 1226 | 12839 ± 0 | 6031 ± 975 | 4922 ± 660 | 3743 ± 121 | 3693 ± 284 | Down |
| An01g06900 | sm to amyR- A. ory | weak similarity to transcription activator amyR - Aspergillus oryzae | 758 ± 96 | 1056 ± 406 | 654 ± 233 | 34 ± 8 | 25 ± 1 | 21 ± 1 | Down |
| An14g06250 | sm to atfB-A. ory | weak similarity to transcription factor atf1+ - Schizosaccharomyces pombe | 269 ± 19 | 523 ± 113 | 434 ± 242 | 67 ± 0 | 67 ± 5 | 89 ± 8 | Down |
| An04g06940 | prtT | similarity to hypothetical transcriptional regulator SPAC1399_1 - Schizosaccharomyces pombe | 2945 ± 971 | 3996 ± 1429 | 2983 ± 738 | 1702 ± 175 | 1499 ± 130 | 608 ± 77 | Down |
| An08g06580 | acuB / FacB | DNA binding protein facB - Aspergillus niger | 364 ± 29 | 340 ± 18 | 400 ± 148 | 899 ± 47 | 929 ± 83 | 656 ± 14 | Up |
| An07g05960 | uncharacterized | similarity to finger protein msn2 - Saccharomyces cerevisiae | 143 ± 21 | 220 ± 133 | 537 ± 288 | 759 ± 389 | 1226 ± 285 | 1125 ± 434 | Up |
| An04g06950 | uncharacterized | similarity to homeobox transcription factor hth - Drosophila melanogaster | 102 ± 1 | 124 ± 6 | 67 ± 16 | 288 ± 24 | 302 ± 50 | 287 ± 54 | Up |
| An08g06850 | uncharacterized | similarity to hypothetical C2H2 zinc-finger protein SPBC1105.14 - Schizosaccharomyces pombe | 66 ± 3 | 75 ± 1 | 129 ± 30 | 583 ± 676 | 787 ± 517 | 791 ± 668 | Up |
| An06g02570 | uncharacterized | similarity to nitrogen assimilation regulatory protein nit-4 - Neurospora crassa | 56 ± 3 | 56 ± 10 | 114 ± 11 | 133 ± 0 | 144 ± 7 | 235 ± 27 | Up |
| An03g05170 | uncharacterized | similarity to sterol regulatory element binding protein-1 srebp-1 - Homo sapiens | 352 ± 68 | 378 ± 126 | 574 ± 9 | 654 ± 59 | 806 ± 134 | 1485 ± 37 | Up |
| An04g06430 | uncharacterized | similarity to zinc finger protein BMZF3 of patent WO9962951-A1 - Homo sapiens | 43 ± 3 | 45 ± 6 | 63 ± 23 | 103 ± 1 | 143 ± 1 | 147 ± 0 | Up |
| An01g05630 | uncharacterized | strong similarity to cytoplasmic aspartate--tRNA ligase APS - Saccharomyces cerevisiae[possible sequencing error] | 70 ± 1 | 75 ± 5 | 63 ± 0 | 189 ± 5 | 214 ± 24 | 164 ± 7 | Up |
| An14g05810 | uncharacterized | strong similarity to histone acetyltransferase GCN5 - Saccharomyces cerevisiae | 63 ± 15 | 58 ± 2 | 46 ± 1 | 148 ± 17 | 140 ± 22 | 85 ± 2 | Up |
| An01g04830 | flbD | strong similarity to myb-like DNA binding protein flbD - Aspergillus nidulans | 89 ± 18 | 91 ± 8 | 81 ± 20 | 238 ± 35 | 220 ± 32 | 107 ± 5 | Up |
| An03g06760 | uncharacterized | strong similarity to probable transcription activator SPAC139.03 - Schizosaccharomyces pombe | 106 ± 0 | 112 ± 14 | 57 ± 13 | 158 ± 8 | 209 ± 42 | 193 ± 22 | Up |
| An01g03750 | abaA | strong similarity to protein abaA - Aspergillus nidulans | 61 ± 9 | 71 ± 12 | 67 ± 17 | 113 ± 28 | 129 ± 3 | 182 ± 22 | Up |
| An02g07890 | pacC | transcription factor pacC - Aspergillus niger | 263 ± 12 | 282 ± 35 | 184 ± 19 | 552 ± 69 | 552 ± 118 | 562 ± 23 | Up |
| An07g09460 | tan1 | transposase Tan1 of patent WO9808960-A1 - Aspergillus niger | 17 ± 4 | 16 ± 3 | 20 ± 2 | 38 ± 3 | 67 ± 10 | 51 ± 6 | Up |
| An08g06030 | uncharacterized | weak similarity to putative zinc finger transcription factor stzA - Aspergillus nidulans | 232 ± 9 | 305 ± 46 | 394 ± 61 | 668 ± 211 | 801 ± 153 | 741 ± 116 | Up |
| An04g00480 | uncharacterized | weak similarity to the vitellogenin II precursor - Gallus gallus | 469 ± 62 | 309 ± 76 | 266 ± 36 | 1222 ± 145 | 1525 ± 171 | 947 ± 325 | Up |
| An11g10870 | uncharacterized | weak similarity to transcription factor CHA4 - Saccharomyces cerevisiae | 193 ± 83 | 180 ± 27 | 273 ± 6 | 626 ± 226 | 622 ± 163 | 560 ± 43 | Up |
| An14g07040 | uncharacterized | weak similarity to transcriptional activator of lysine pathway genes LYS14 - Saccharomyces cerevisiae | 76 ± 5 | 71 ± 10 | 70 ± 1 | 178 ± 10 | 217 ± 56 | 141 ± 8 | Up |
| An17g00800 | uncharacterized | weak similarity to transcriptional activator prtT of patent WO200020596-A1 - Aspergillus niger | 119 ± 48 | 89 ± 35 | 69 ± 5 | 201 ± 38 | 195 ± 22 | 142 ± 5 | Up |
| An02g06000 | uncharacterized | weak similarity to ZMS1 - Saccharomyces cerevisiae | 471 ± 242 | 481 ± 196 | 683 ± 362 | 980 ± 41 | 1338 ± 83 | 1688 ± 256 | Up |
Table 9: Hybridization values of 38 transcription factor genes [18] that are differentially expressed in wild-type when compared to the ΔflbA strain. Names of transcription factors are indicated that show homology toor that are encoded by A. niger genes. *Description of gene or its closest homologue. Sm: Similar; A. fum: A. fumigatus; A. ory: A. oryzae.
Colonies of an A. niger strain in which flbA is inactivated do not sporulate. Instead, they grow throughout the colony and show secretion in a large part of the mycelium [8]. Moreover, cell walls of the ΔflbA strain are thinner than those of the wild-type [8]. The molecular mechanisms underlying these phenomena are not known. Here, the impact of inactivation of flbA on spatial gene expression in a sandwiched grown A. niger colony was assessed with emphasis on genes encoding secreted proteins, genes involved in cell wall synthesis, genes involved in asexual and sexual development and genes encoding transcriptional regulators. The latter genes may control spatial growth and secretion, cell wall synthesis or may have a role in reproduction.
Genes that could account for a sexual pathway have been identified in A. niger [18]. However, so far only asexual reproduction has been shown to occur in this fungus. Asexual development involves the formation of conidia forming conidiophores. This process has been studied in most detail in A. nidulans [3,26]. Based on its genome, A. niger is expected to have similar mechanisms underlying conidiophore development. Indeed, the central regulator of conidiophore formation brlA, as well as flbA have a similar role in asexual development in A. niger and A. nidulans [8,27]. Inactivation of flbA of A. niger does not have a major impact on the expression of genes involved in asexual development, at least in colonies grown as a sandwiched culture as used in this study. Of the 68 genes implicated in reproduction, only 13 were differentially expressed when compared to the wild-type. Genes flbD and sfaD (Figure 1) were among the 6 genes that were up-regulated ≥ 2-fold in the ΔflbA strain. Notably, flbD and sfaD expression was not increased in the ΔflbA strains of A. nidulans and A. oryzae [28,29]. Transcripts of brlA were absent in wild type A. niger colonies, while some brlA expression was observed in the center of ΔflbA colonies. In contrast, FlbA directly or indirectly stimulates expression of brlA in A. nidulans [9], A. oryzae [29] and A. fumigates [30]. It can thus be concluded that the sporulation pathway of A. niger is similar but not identical to that of A. nidulans, A. oryzae and A. fumigatus. Our data also indicate that FlbA represses asexual reproduction in zones of A. niger colonies that do no longer have the potential to sporulate. It might be that this also occurs in other Aspergilli. This should be studied by extracting RNA of zones rather than the whole mycelium, as is routinely done.
A total of 102 genes are predicted to be involved in cell wall synthesis [18]. Of these genes, only 10 and 2 were found to be up- and downregulated in the ΔflbA strain, respectively. It is tempting to speculate that these genes are involved in the changed spatial distribution of growing hyphae in the colony. In this respect, the up-regulation of two chitin synthase genes is of particular interest. The chsD homologue of A. nidulans is expressed throughout the A. niger colony. This gene is upregulated in all zones of ΔflbA colonies. The chsF chitin synthase gene is even more interesting. This gene is periphery specific in wild-type colonies, while it is expressed throughout ΔflbA colonies. Zonal expression of chsF thus correlates with growth in colonies of wildtype and ΔflbA. The differentially expressed genes involved in cell wall synthesis may also impact cell wall thickness in the ΔflbA strain. This may involve synthesizing activity, cell wall processing activity (i.e. crosslinking), as well as degradation activity.
The secretome of the wild-type and the ΔflbA strain was determined by transferring 7-day-old colonies that had been grown on agar medium to a ring plate containing fresh medium. Proteins were released in the wells of the ring plate that had been formed during the 24 h incubation time. In addition, proteins are released that had been formed during the 7 days of growth on the agar plate and that slowly diffuse through the cell wall [4]. FlbA was shown to have a more complex secretome when compared to the wild-type [8]. Out of 138 secreted proteins of the ΔflbA strain [8], 101 had not been identified in the secretome of wild-type colonies [8]. Cycloheximide treatment releases proteins into the culture medium that had been trapped in the cell walls of the wild-type strain [8]. Still, 70 secreted proteins were found in the ΔflbA secretome that were absent in the medium of cycloheximide treated wild-type colonies. Of these proteins, 23 were differentially expressed, of which 22 were up-regulated in the ΔflbA strain. This shows that part of the differences in the secretome are caused by transcriptional control during the 7 days of growth on the agar medium. Another part of the differences may be explained by induction of genes after transfer to fresh medium and by (post)-translational regulation. Moreover, we can not exclude that part of the proteome is still trapped in or associated with the wild type cell wall after cycloheximide treatment. 55 proteins were identified in the wild-type secretome that were absent in the culture medium of the ΔflbA strain [8]. Of the genes encoding these proteins, 35 were differentially expressed, of which 33 were downregulated in the ΔflbA strain. This shows that transcriptional control during growth on the agar medium has a major impact on genes encoding proteins that are released by the wild-type strain but not by the ΔflbA strain. Finally, 20 out of 37 genes encoding proteins that were released both in wild-type and the ΔflbA secretome were differentially expressed. Quantitative proteomics should reveal whether this differential expression also results in quantitative differences in levels of these proteins in the medium. In this study, A. niger was grown on xylose. XlnR is the xylanolytic regulator that controls expression of at least 16 genes [19-25]. All encoded proteins were identified in the wild-type secretome after cycloheximide treatment, whereas 15 out of 16 proteins were identified in the ΔflbA secretome. 6 out of 16 XlnR regulated genes were down-regulated in the ΔflbA strain. This is not in conflict with the proteomics data per se, since quantitative proteomics was not performed on wild-type versus ΔflbA. Moreover, proteomics was performed on colonies that had been transferred to fresh medium (see above).
Spatial growth and secretion and cell wall synthesising, modifying and degrading activities are most probably regulated by transcriptional regulators. In total, 38 transcriptional regulators were found to be differentially regulated in the ΔflbA strain. Most of these regulators have not been characterised. However, the list includes 9 genes with a known function or that are homologous to a transcription factor that has been characterized in other Aspergillus species. Genes that have homology to rosA of A. fumigatus and atfB of A. oryzae (both downregulated in ΔflbA), and to flbD and abaA of A. nidulans (both upregulated in ΔflbA) are involved in development (rosA, flbD, and abaA) and stress tolerance of conidia (attB) [3,31-33]. Genes homologous to xlnR and amyR of A. oryzae, prtT [34] and acuB [35,36] (all downregulated in ΔflbA) are predicted to be involved in degradation of polysaccharides, proteins and acetate. Gene prtT has been shown to be the transcriptional activator of protease genes in A. niger [34]. Its down-regulation in ΔflbA colonies is in agreement with the finding that only 6 protease geneswere up-regulated in ΔflbA, while 13 were down-regulated. The transcriptional regulatory gene pacC [36] was found to be up-regulated at the periphery of ΔflbA colonies. It is not known how pacC expression is exactly regulated in Aspergillus. The pH does not seem to regulate its expression or that of the six pH-sensing pal components [37]. However, PacC protein is abundant in alkaline growth conditions, where it is activated by proteolytical cleavage [37-39]. At alkaline pH, alkaline-associated genes of A. nidulans are activated by PacC (palD, prtA, xlnA, acvA, ipnA, pacC, sidA, mirA, mirB) [40], while acid-associated genes (pacA, xlnB, abfB, gabA, and stcU) are repressed [41]. These genes are not fully annotated in A. niger, but homologues of mirA (An02g14190), mirB (An03g03560) and sidA (An05g00220) are up-regulated in ΔflbA, whereas xlnB, putative gabA (An12g10000) and xlnA are down-regulated when compared to wildtype. This might be explained by the overexpression of PacC since the pH of the agar-medium underneath wild-type sandwiched colonies was 3, while it was 6 in the case of the ΔflbA strain. Genes An11g06290 and An01g06900 are homologous to the xylanolytic and amylolytic regulatory genes xlnR and amyR of A. oryzae, respectively. However, they do not encode the XlnR and AmyR regulators of A. niger since these functions have been attributed to An15g05810 [20,24,25] and An04g06910, respectively [42]. The xlnR gene of A. niger was downregulated 1.5 fold, which may explain the high number (i.e. 9 out of 16) of xylanolytic genes that are down-regulated compared to wild-type. Apart from the 9 transcriptional regulatory genes with a predicted function, there are 29 of such genes with an unknown role in growth and development of A. niger [43]. One or more of these genes may be involved in spatial growth, spatial secretion, cell wall formation and asexual development. As such, they are of interest to improve A. niger as a cell factory.