Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2015) Volume 3, Issue 2
A reproducible regeneration system based on direct somatic embryogenesis is described for Kenya cassava lines. Cassava plants were regenerated at high frequency by inducing shoot primordial on explants derived from cotyledons of cassava somatic embryos.Various parameters were evaluated on their effects on callus induction, somatic embryogenesis, maturation and germination of somatic embryos as well as recovery of regenerated plantlets. Immature leaf lobes were used as explants for somatic embryogenesis. Three Kenyan cassava genotypes viz; Adhiambo Lera, Kibanda Meno and Serere along with a model cultivar, TMS 60444 were used this system. Remarkable regeneration frequencies were observed in all the evaluated genotypes with Adhiambo Lera showing the best responses. As a result, a highly efficient plant regeneration protocol via germination of somatic embryos was achieved. This system enriches the scope of in vitro regeneration protocols for cassava and is envisaged to be a reliable prerequisite to genetic transformation of African cassava genotypes
Keywords: Somatic embryogenesis; Immature leaf lobes; Kenyan Cassava genotypes; Regeneration
Cassava (Manihot esculenta Crantz, Euphorbiaceae, 2n=36) is one of the most important food crops in the world, especially in the tropics [1]. Its starchy tuberous roots provide a valuable source of cheap calories for about 500 million people in the developing world commonly plagued by chronic food deficiency and malnutrition [2]. In addition, the leaves and tender shoots are eaten in many parts of Africa as a source of vitamins, minerals and protein [3]. It is tolerant to low fertility and drought, which makes it popular among small-scale farmers in places with infertile soils and adverse climates [4]. Cassava is also used in the production of ethanol for fuel, for animal feed, and as a raw material for the starch industry.
Despite these pleasant qualities, cassava has its drawbacks. It contains cyanogenic glycosides which when not effectively removed may cause a variety of health problems in nutritionally-compromised peoples [5]. Due to its low protein content (1-2%) and limited amounts of sulphurcontaining amino acids, additional food sources are required to ensure a diet balanced in protein, vitamins and minerals [6,7]. During the long cultivation period (up to 18 months) of cassava, repeated attacks by various insect pests and virus diseases can cause 20-50% yield losses worldwide, and locally they can lead to total crop failures [8,9]. Further, cassava suffers from postharvest physiological deterioration during transport, storage and marketing [10]. Although the roots can remain in the ground for many months, once they are harvested they deteriorate rapidly and within 48 hours they are unmarketable. In spite of its high importance to food security in third world countries, cassava has long been neglected in plant breeding programmes [11]. The shortcomings associated with cassava production as well as the undesirable traits are potentially amenable to genetic engineering techniques particularly since genetic improvement through traditional breeding has been problematic due to low seed production and long generation time.A critical requirement for the generation of transgenic cassava is an efficient and reproducible plant regeneration system that is compatible with the available transformation methods. Somatic embryos have been induced from cassava young leaf lobes and cotyledons leading to plant regeneration [12-17].The reported cassava regeneration protocols have seldom optimally worked for Kenyan cassava varieties. Apart from a few studies by [17,18], little if any efforts have focused on developing regeneration and transformation systems for African cassava varieties [2]. Further, it is reported that African cassava varieties from Africa respond differently in culture from varieties from other parts of the world. Environmental, biological, and human demands for African varieties may be responsible for genetic divergence of African cassava from their South American progenitors potentially accounting for the different responses in culture (Sayre, personal communication). In this regard, it is imperative to develop and optimize a regeneration protocol that can feasibly work for Kenyan and perhaps African cassava varieties, as a pre-requisite for successful genetic transformation of Kenyan cassava varieties. In this paper, we report successful development of an improved regeneration protocol for Kenyan cassava varieties via somatic embryogenesis.
Plant materials
Four cassava varieties viz; Adhiambo Lera, Kibanga Meno, Serere and TMS60444 were sourced from Kenya Agricultural Research Institute (KARI), Kakamega. The plantlets were grown in vitro on Murashige and Skoog media [19] supplemented with 20 g/L sucrose, MS Vitamins (Duchefa, Germany) and 8 g/L of noble agar. All media used for in vitro propagation of cassava was sterilized through autoclaving. The growth chamber conditions were set at a temperature of 28°C and a 16 hr day/8 night cycle.
Callus induction and somatic embryogenesis
Meristematic leaf lobes (2-6 mm long) from in vitro-grown plants were cultured on MS basal medium supplemented with 2% (w/v) sucrose, B5 vitamins, 50 mg/L casein hydrolysate, 0.5 mg/L CuSO4 [20] and 4–16 mg/L 2,4-dichlorophenoxyacetic acid (2,4-D). The same set of meristematic leaf lobes was put in the same media substituted with Picloram. The media pH was adjusted to 5.7 and it was solidified with 0.8% (w/v) noble plant agar. Factors affecting embryogenesis were studied with 0–8% sucrose, 0–1.0 mg/L additional CuSO4 and keeping the embryogenesis cultures in darkness or on a 12 hr day/12 hr night cycle. The cultures were maintained at a temperature of 28°C. The explants were left in the induction medium for 4–6 weeks. The type of calli was observed at each step and the frequency of embryogenic calli formation was recorded four weeks of culture on callus induction medium (CIM).
Maturation of somatic embryos
This entailed the development of globular stage embryos into green cotyledonary embryos with defined shoot and root axes [14]. The globular stage somatic embryos were subcultured on maturation media composed of MS salts [19] supplemented with 2% (w/v) sucrose, 1 mg/L thiamine-HCl, 100 mg/L myo-inositol, 0.01 mg/L 2,4-D, 1.0 mg/L BA, and 0.5 mg/L GA3; MS medium supplemented with 2% (w/v) sucrose, 0.01 mg/L NAA, 0.1 mg/L BA and 0.1 mg/L GA3; or MS basal medium supplemented with 2% (w/v) sucrose. The media pH was solidified with 0.8% (w/v) Difco-Bacto agar. The embryos were maintained in the maturation medium in the dark for 4 weeks.
Germination and plant recovery
Mature somatic embryos were transferred to basal MS medium supplemented with 2% (w/v) sucrose; MS medium supplemented with 2% (w/v) sucrose, 1 mg/L thiamine-HCl, 100 mg/L myo-inositol, 0.01 mg/L 2,4-D, 1.0 mg/L BA and 0.5 mg/L GA3; MS basal salt supplemented with 2% (w/v) sucrose and 0.8% (w/v) activated charcoal. All the media were solidified with 0.8% (w/v) noble agar for germination and plant recovery. Germination and conversion rates were recorded after four weeks in culture. The cultures were exposed to a daily photoperiod of 12 hours. All cultures were kept at 28°C. Some regenerated plantlets were maintained in growth room ready for transformation experiments while others were hardened in the glass house.
Data analysis
All data were expressed as mean ± SEM (n=3). The collected was subjected to one-way ANOVA. Statistically significant means were analysed by Tukey’s Pairwise comparisons at 95% level of significance.
Effects of 2,4-D and Picloram on callus induction and somatic embryogenesis
It was possible to induce calli in all the cassava varieties at all the concentrations of 2,4-D and picloram. Four concentrations of 2, 4-D and picloram (4, 8, 12 and 16 mg/L) were tested for their ability to induce calli and somatic embryogenesis. The different auxin treatments were used to determine which concentration was best for calli and somatic embryo induction of the cassava cultivars. All the treatments produced calli and somatic embryos. For both callus induction and somatic embryogenesis, the best auxin concentration was 8 mg/L of both 2,4-D and picloram, although 2,4-D had better response than picloram (Tables 1 and 2). 12 mg/L of both 2,4-D and picloram also induced calli and embryogenesis appreciably compared to 4mg/L and 16mg/L, which induced the least responses. Adhiambo Lera and Serere were the best responding varieties with regards to callus induction and somatic embryogenesis (Tables 1 and 2). There was no significant difference in the frequency of callus induction and somatic embryogenesis at auxin concentrations of 4mg/L and 16mg/L (P>0.05). Formation of embryogenic calli was consistent with the frequency of callus induction in all the cassava varieties. Embryogenesis began as swollen regions at the cut ends and mid-veins on the adaxial surface of the immature leaf lobes. The swollen segments then gave rise to two kinds of calli viz; a loose non-embryogenic friable white callus and a translucent gelatinous embryogenic callus that formed globular stage embryos (Figure 1).
| Average number of calli per explant | ||||||||
|---|---|---|---|---|---|---|---|---|
| 4 | 8 | 12 | 16 | |||||
| 2,4-D | Picloram | 2,4-D | Picloram | 2,4-D | Picloram | 2,4-D | Picloram | |
| TMS 6044 | 64.3 ± 0.9b | 60.3 ± 0.9b | 85.0 ± 0.6a | 81.0 ± 0.6c | 74.3 ± 0.9b | 70.0 ± 0.6b | 68.2 ± 2.4bc | 58.0 ± 1.2b |
| AdhiamboLera | 75.0 ± 1.5a | 71.7 ± 1.2a | 93.0 ± 1.8c | 90.0 ± 1.2a | 82.7 ± 0.9a | 78.7 ± 1.3a | 68.3 ± 0.9a | 65.0 ± 2.3a |
| KibandaMeno | 58.3 ± 1.2c | 55.3 ± 0.9c | 74.7 ± 2.4b | 71.0 ± 0.6b | 67.7 ± 1.5b | 63.0 ± 1.5b | 57.0 ± 1.7c | 54.3 ± 2.3b |
| Serere | 66.3 ± 0.9b | 61.3 ± 0.9b | 78.7 ± 0.9b | 75.3 ± 1.8b | 68.7 ± 1.9bc | 65.7 ± 2.9b | 1.3 ± 0.9b | 57.1 ± 2.1b |
Table 1: Effects of the different concentrations 2,4-D and Picloram (mg/L) on callus induction.
| Average number of embryonic calli per explant | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 4 | 8 | 12 | 16 | ||||||
| 2,4-D | Picloram | 2,4-D | Picloram | 2,4-D | Picloram | 2,4-D | Picloram | ||
| TMS 6044 | 20.7 ± 1.2a | 20.5 ± 3.4a | 33.3 ± 2.0b | 29.7 ± 3.0a | 26.7 ± 0.9a | 27.7 ± 1.5b | 21.0 ± 0.6a | 19.3 ± 1.8b | |
| AdhiamboLera | 27.0 ± 2.7a | 26.3 ± 2.3b | 40.7 ± 0.9a | 39.4 ± 3.2b | 37.3 ± 1.2b | 33.8 ± 1.9a | 26.7 ± 0.9b | 27.7 ± 1.5a | |
| KibandaMeno | 22.0 ± 2.3a | 23.7 ± 3.6a | 34.7 ± 0.9ab | 33.6 ± 2.6c | 33.0 ± 0.6c | 29.8 ± 2.5ab | 22.7 ± 0.3a | 18.7 ± 1.9b | |
| Serere | 27.0 ± 0.6a | 22.2 ± 2.4a | 37.7 ± 1.5ab | 42.4 ± 3.1b | 33.3 ± 1.7b | 30.5 ± 2.3ab | 31.3 ± 0.9c | 19.8 ± 1.6b | |
Table 2: Effects of the different concentrations 2,4-D and Picloram (mg/L) on somatic embryogenesis.
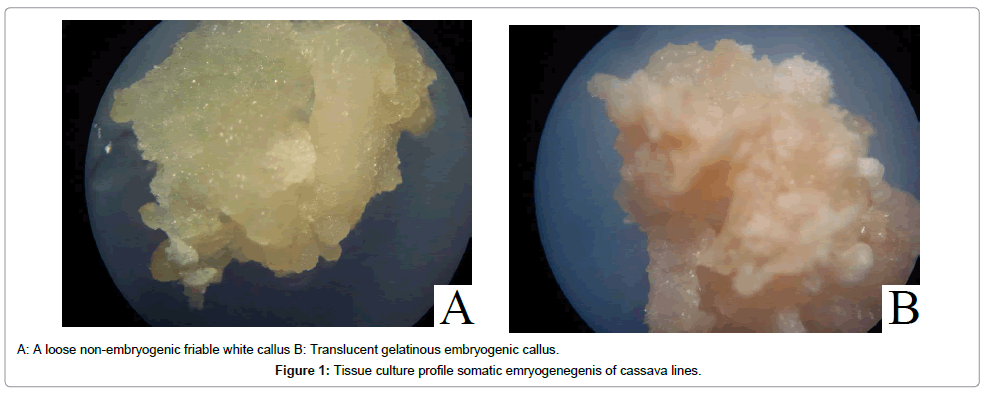
Figure 1: Tissue culture profile somatic emryogenegenis of cassava lines.
Effect of light on somatic embryogenesis
The effect of light on somatic embryogenesis was done by exposing the somatic embryo cultures to 16 hours/day photoperiod versus 8 hours/day dark incubation. This was observed to affect the process of somatic embryogenesis. Following four weeks in culture, the light-exposed explants formed more non-embryogenic calli than those kept in the dark (Table 3). Generally, a few embryos were produced when the explants were given 12 hours of light followed by 12 hours of darkness. With dark incubation, Adhiambo Lera and Serere showed the best responses with 36.6% and 33.8% of lightexposed explants developing somatic embryos respectively compared to 87.7% and 80.1% of dark incubated explants respectively. The two cassava varieties had the highest mean numbers of embryogenic calli per explants compared to TMS 60444 and Kibanda Meno (Table 3). TMS 60444 and Kibanda Meno had 32.5% and 31.5% of light-exposed explants producing somatic embryos respectively compared to 75.4% and 74.1% of dark incubated explants respectively. Nevertheless, light is required for germination of embryos [21]. Adhiambo Lera had the best response with regards to callus induction of explants exposed to darkness compared to other genotypes (P<0.05) (Table 3). However, there was no significant difference in the number of embryonic calli produced per explants between Adhiambo Lera and Serere genotypes.
| Total number of explants | No. ofexplants with developed callus | Mean no. of embryonic calli /explants | ||||
|---|---|---|---|---|---|---|
| 12 | 0 | 12 | 0 | 12 | 0 | |
| TMS 60444 | 124.0 ± 1.7 | 125.0 ± 1.7 | 40.3 ± 1.2a (32.5%) |
94.3 ± 2.3a (75.4%) |
12.7 ± 1.8a | 127.3 ± 2.3a |
| AdhiamboLera | 125.7 ± 2.9 | 121.7 ± 4.9 | 46.0 ± 2.3b (36.6%) |
106.7 ± 1.8b (87.7%) |
17.7 ± 2.0ab | 161.3 ± 5.8b |
| KibandaMeno | 115.3 ± 2.9 | 123.7 ± 2.7 | 36.3 ± 1.8ab (31.5%) |
91.7 ± 1.2a (74.1%) |
13.0 ± 1.7a | 130.3 ± 0.9a |
| Serere | 125.0 ± 2.5 | 122.0 ± 1.2 | 42.3 ± 2.7ab (33.8%) | 97.7 ± 2.6a (80.1%) |
25.7 ± 2.2b | 159.3 ± 1.2b |
Table 3: Effect of light on somatic embryogenesis.
Effects of copper sulphate
The effect of additional Copper in the form of CuSO4 was studied with all the cultivars by the inclusion of 0.25, 0.5, 0.75 and 1.0 mg/L of additional CuSO4 in the induction medium. The results were cultivardependent and consistent (Table 4). For cultures devoid of additional copper and those with 0.25 mg/L, formation of embryogenic calli was comparable in all varieties (P>0.05). For all varieties except TMS 60444, formation of embryogenic calli with inclusion of 0.5, 0.75 and 1 mg/L of additional Copper was also comparable. However, 0.5 mg/L CuSO4 had the best response with regards to the number of embryonic calli per explants (Table 4).
| Average number of embryonic calli per explant | ||||||
|---|---|---|---|---|---|---|
| 0.0 | 0.25 | 0.5 | 0.75 | 1 | ||
| TMS 6044 | 23.7 ± 2.0a | 25.3 ± 1.2a | 43.7 ± 1.9a | 34.0 ± 0.6a | 27.3 ± 3.1a | |
| AdhiamboLera | 31.0 ± 1.2a | 33.0 ± 1.7a | 62.7 ± 1.2b | 53.0 ± 1.7b | 44.3 ± 2.5b | |
| KibangaMeno | 25.7 ± 2.2a | 32.0 ± 3.2a | 52.3 ± 1.9b | 50.0 ± 0.6b | 39.0 ± 0.6b | |
| Serere | 29.0 ± 1.2a | 32.0 ± 1.2a | 59.0 ± 0.6b | 1.7 ± 5.2ab | 40.0 ± 1.2b | |
Table 4: Effect of different CuSO4 (mg/L) concentrations on somatic embryogenesis of cassava.
Effect of sucrose on embryogenesis
Callus formation and embryogenesis hardly took place with the exclusion of sucrose from the callus induction medium (Table 5). The explants died after four weeks in culture. The number of embryos produced in 2-6 % sucrose media was cultivar dependent. 2% sucrose concentration promoted formation of healthier embryos than the others. Explants grown on 6% sucrose were all cream-colored and most turned brown after 4 weeks in culture. This is a sign of cell death caused by formation of phenolic compounds in the cultures [2]. Adhiambo Lera and Serere genotypes produced the highest number of embryonic calli with all the sucrose concentrations (Table 5). However, the response of Adhiambo Lera significantly different from the response of Serere at 4% and 6% sucrose concentrations (P<0.05). Kibanda Meno and the model cultivar TMS 60444 had comparable responses at 2% and 4% sucrose concentrations (Table 5).
| Sucrose concentration (percent w/v) | ||||||
|---|---|---|---|---|---|---|
| Cultivar | 0 | 2 | 4 | 6 | ||
| No. of embryonic calli | ||||||
| TMS 6044 | 0.0 ± 0.0 | 84.0 ± 2.3a | 77.3 ± 1.7a | 39.3 ± 1.2a | ||
| AdhiamboLera | 0.0 ± 0.0 | 110.7 ± 5.8c | 108.0 ± 2.3b | 52.0 ± 1.2c | ||
| KibangaMeno | 0.0 ± 0.0 | 89.3 ± 2.9ab | 79.0 ± 1.5a | 32.0 ± 1.2b | ||
| Serere | 0.0 ± 0.0 | 106.3 ± 3.6bc | 97.0 ± 1.5c | 37.3 ± 1.8ab | ||
Table 5: Effect of different sucrose concentrations on somatic embryogenesis.
Maturation of somatic embryos
The early stage somatic embryos were transferred to the maturation medium after 4 weeks of culture with adjoining callus. All the embryos grew to cotyledonary embryos with distinct root and shoot axes in all media compositions albeit with differences in frequencies. Three different maturation media differing in cytokinins content were used in this experiment. Embryos maintained on MS medium supplemented with 2% sucrose, 1 mg/L thiamine-HCl, 100 mg/L myo-inositol, 0.01 mg/L 2,4-D, 1.0 mg/L BA and 0.5 mg/L GA3 was superior to MS medium supplemented with 2% sucrose, 0.01 mg/L NAA, 0.1 mg/L BA and 0.1 mg/L GA3 and MS medium supplemented with 2% sucrose. As Figure 2 shows, more than 50% percent of embryonic calli produced cotyledonary embryos in the medium containing MS salts supplemented with 2% (w/v) sucrose, 1 mg/L thiamine-HCl, 100 mg/L myo-inositol, 0.01 mg/L 2,4-D, 1.0 mg/L BAP and 0.5 mg/L GA3. In the medium supplemented with 0.01 mg/L NAA, 0.1 mg/L BA and 0.1 mg/L GA3, more than 70% of embryonic calli hardly produced cotyledonary embryos. The medium supplemented with 2% sucrose promoted the least frequency of maturation (Figure 2). Overall the cultivar Adhiambo Lera had the highest frequency of maturation of somatic embryos followed by Serere and TMS 60444. Kibanda Meno had the least response in all the media compositions. As figure 3 shows, the embryos first of all turned green and then progressively formed distinct shoot and root apices. However, there was no significant difference in the frequency of maturation of somatic embryos of Adhiambo Lera and Serere in the media containing MS salts supplemented with 2% (w/v) sucrose, 1 mg/L thiamine-HCl, 100 mg/L myo-inositol, 0.01 mg/L 2,4-D, 1.0 mg/L BAP and 0.5 mg/L GA3 (P>0.05) (Table 6). In the same media, the responses of TMS 60444 and Serere genotypes were comparable.
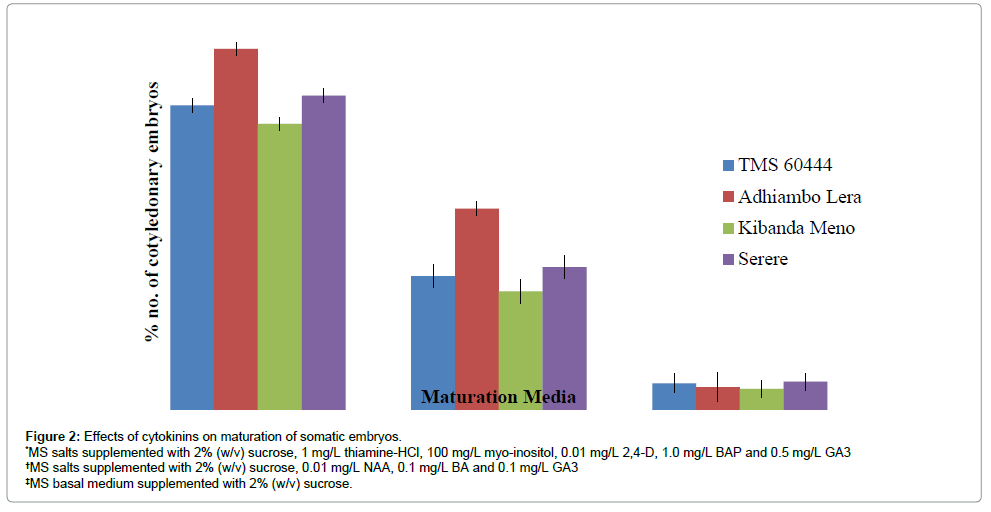
Figure 2: Effects of cytokinins on maturation of somatic embryos.
*MS salts supplemented with 2% (w/v) sucrose, 1 mg/L thiamine-HCl, 100 mg/L myo-inositol, 0.01 mg/L 2,4-D, 1.0 mg/L BAP and 0.5 mg/L GA3
†MS salts supplemented with 2% (w/v) sucrose, 0.01 mg/L NAA, 0.1 mg/L BA and 0.1 mg/L GA3
‡MS basal medium supplemented with 2% (w/v) sucrose.
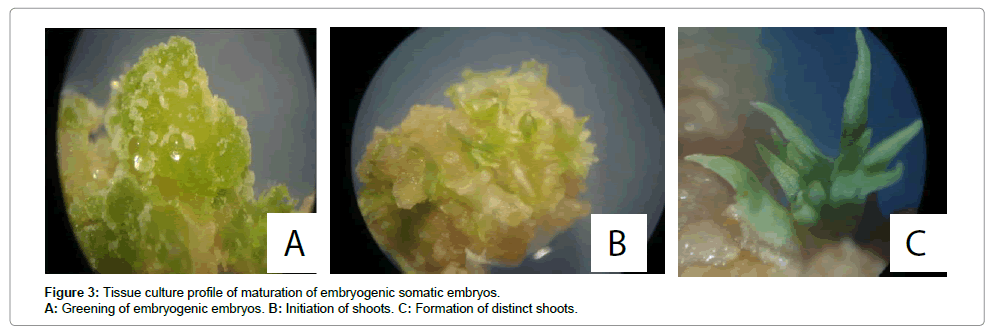
Figure 3: Tissue culture profile of maturation of embryogenic somatic embryos. A: Greening of embryogenic embryos. B: Initiation of shoots. C: Formation of distinct shoots.
| Mean no. of cotyledonary embryos | |||||
|---|---|---|---|---|---|
| Cultivar | Initial no. of embryonic calli | Maturation media 1* | Maturation media 2† | Maturation media 3‡ | |
| TMS 60444 | 98.3 ± 1.5 | 53.3 ± 0.9ac | 23.9 ± 0.9b | 4.7 ± 1.2a | |
| AdhiamboLera | 91.3 ± 0.9 | 58.3 ± 1.2a | 33.0 ± 1.2a | 6.3 ± 1.8a | |
| KibangaMeno | 90.7 ± 1.2 | 46.3 ± 1.5b | 19.3 ± 2.0b | 3.3 ± 1.2a | |
| Serere | 90.7 ± 1.8 | 50.7 ± 1.2bc | 23.0 ± 1.7b | 4.3 ± 0.9a | |
Table 6: Effects of different cytokinins on maturation of somatic embryos
Germination of somatic embryos and plant recovery
The germination medium comprising MS salts, B5 vitamins, 2% sucrose, and 0.8% activated charcoal was superior to basal MS medium plus 2% sucrose, and MS supplemented with 2% sucrose, 1 mg/L thiamine-HCl, 100 mg/L myo-inositol, 0.01 mg/L 2,4-D, 1.0 mg/L BA and 0.5 mg/L GA3 in terms of germination time and frequency (Table 7). Single cotyledonary embryos did not always produce single plantlets. In some cases up to five plantlets arose from one embryo. The recovery of plantlets from embryos was genotype-dependent and it was consistent. Overall, Adhiambo Lera was the best genotype in terms of both germination time and frequency. As table 7 shows, there was significant difference between Adhiambo Lera and other genotypes in all media compositions in terms of germination frequency (P<0.05). On the other hand, the other genotypes; TMS 60444, Kibanda Meno and Serere had comparable germination frequencies in the basal MS medium supplemented with B5 vitamins and 2 % (w/v) sucrose and in MS medium supplemented with 2% (w/v) sucrose, 1 mg/L thiamine- HCl, 100 mg/L my-oinositol, 0.01 mg/L 2,4-D, 1.0 mg/L BAP and 0.5 mg/L GA3 (P>0.05). Upon germination, the recovered plants were grown in the growth room for two weeks following which they were hardened in the glass house. Growth of the regenerated plantlets is summarized in Figure 4.
| Mean no. of cotyledonary embryos | |||||
|---|---|---|---|---|---|
| Cultivar | Initial no. of embryonic calli | Maturation media 1* | Maturation media 2† | Maturation media 3‡ | |
| TMS 60444 | 98.3 ± 1.5 | 53.3 ± 0.9ac | 23.9 ± 0.9b | 4.7 ± 1.2a | |
| AdhiamboLera | 91.3 ± 0.9 | 58.3 ± 1.2a | 33.0 ± 1.2a | 6.3 ± 1.8a | |
| KibangaMeno | 90.7 ± 1.2 | 46.3 ± 1.5b | 19.3 ± 2.0b | 3.3 ± 1.2a | |
| Serere | 90.7 ± 1.8 | 50.7 ± 1.2bc | 23.0 ± 1.7b | 4.3 ± 0.9a | |
Table 6: Effects of different cytokinins on maturation of somatic embryos
.

Figure 4: Recovery of regenerated cassava plantlets.
A: Young cassava plantlets immediately transferred to soil in glass house from growth room plantlets. B: Mature cassava plants in glass house. C: Mature cassava plantlets in the field outside glasshouse.
In this study, a protocol for somatic embryogenesis and plant recovery for Kenyan cassava varieties has been optimized. Studies on African cassava varieties by [2] established that 12 mg/L 2, 4-D was better than 12 mg/L picloram for embryogenesis induction for most of the cultivars. Conversely, in this study, it was established that 8mg/L 2,4-D and Picloram produced the best responses in terms of callus induction and somatic embryogenesis in all cultivars. Moreover, 2,4-D (8 mg/L) was better with regards to frequency of leaf lobe embryogenesis than Picloram (8 mg/L) in all cultivars (Table 1 and 2). Nevertheless, embryogenesis was induced at all auxin concentrations in all cultivars. Successful somatic embryogenesis in Glycine max on medium supplemented with 40 mg/L 2, 4-D has been accomplished in the past [22]. This is in spite of the studies by [17], who reported an inhibition of embryogenesis in cassava by the supplementation of the induction-medium with more than 12 mg/L 2, 4-D. The observed disparities in the results of this study can be blamed on use of different explants. This study used young leaf lobes, while Konan’s group used the cotyledons of zygotic embryos.
Many plant species regenerated in vitro through callus are bedeviled by somaclonal variation [2]. This is postulated to be due to the high rate of cell division during callus formation induced by high auxin levels in the induction medium. This drawback is reduced in plants recovered through somatic embryogenesis [17]. Observed that the incidence of somaclonal variation in cassava regenerated by somatic embryogenesis with 4-16 mg/L 2,4-D is negligible. As a convention, embryogenesis in most plants starts with callus induction, followed by transfer to another medium for embryo emergence. In cassava, the explant on the induction medium proliferates to globular stage embryos in about two weeks of culture, hence reducing the length of time the regenerative tissues stay on high auxin (8-12 mg/L) medium [15]. Perhaps this ameliorates the incidence of somaclonal variation further in cassava plants recovered from somatic embryos. Recently, there have been more reports of somaclonal variation in cassava regeneration systems [16,21]. The drive to improve on the efficiency of cassava transformation has led to the development of additional techniques for plant regeneration. A classical example of such efforts is the use of friable embryogenic callus to regenerate somatic embryos. This technique requires the callus to be on the high auxin medium (50 mg/L) for six or more months resulting in high rates of somaclonal variation [21,23]. Copper is thought to enhance growth of embryos during somatic embryogenesis. Somatic embryogenesis experiments performed with media lacking additional copper produced embryos difficult to distinguish from the non-embryogenic callus because they were too small [2]. However, the inclusion of additional copper made the embryos distinct at the early stage of embryogenesis. This was also the case in this study. Presumably, it is due to the fact that Ihemere worked on African cassava varieties.
Callus formation is often considered to inhibit plant regeneration and in cassava plant regeneration from callus has never been possible, neither via organogenesis nor via embryogenesis. It is therefore assumed that the increased capacity of shoot organogenesis is at least partially related to the inhibition of callus formation [24]. Cassava regeneration via organogenesis involves the induction of shoots from young leaf lobes and embryo cotyledons on MS medium supplemented with cytokinins (23 μM zeatin and 44 μM benzylaminopurine) [25,26]. This method results in the regeneration of multiple shoots but the origin of each shoot is doubtful making it suspect for plant transformation purposes [2]. Embryo development was achieved in medium supplemented with 2% sucrose, 1 mg/L thiamine-HCl, 100 mg/L my-oinositol, 0.01 mg/L 2,4-D, 1.0 mg/L BA and 0.5 mg/L GA3 was superior to MS medium supplemented with 2% sucrose, 0.01 mg/L NAA, 0.1 mg/L BA and 0.1 mg/L GA3 and MS medium supplemented with 2% sucrose but devoid of phytohormones. This agrees with [15], who cultured globular stage embryos of cassava on medium without plant growth regulators and achieved the highest rate of regeneration yet in cassava. It can therefore be inferred that phytohormones may only be critical in the induction medium for embryogenesis to occur.
Culturing explants in light at the beginning of the induction process led to reduction in the number of embryos formed. This indicates that light-intensity affects embryogenesis. Embryogenesis in cassava is enhanced by lowering light intensity [21]. Studies on the effect of light on callus growth and somatic embryogenesis from Lavandula vera and Teucrium chamaedrys showed that the relative length of the incubation period under illumination significantly affected callus growth and somatic embryo induction and proliferation, although in a species-specific fashion. Lavender callus growth was improved under increased incubation in darkness while somatic embryogenesis was remarkably reduced under the same conditions [27]. Further, somatic embryogenesis from some species, such as cucumber, squash, melon and gardenia is promoted under initial culture incubation in darkness [27-29]. It is, thus, thought that lowering the light intensity of the cultures could have accounted for the high rates of embryogenesis in all the cultivars studied in this research.
In conclusion, this study has optimized a reproducible in vitro regeneration pathway of Kenyan genotypes using three Kenyan genotypes viz; Adhiambo Lera, Kibanda Meno and Serere along with an exotic model cultivar TMS 60444 using immature leaf lobes as explants for somatic embryogenesis. It avails an opportunity for further studies on cassava with varying research interests to mitigate the various genetic and phenotypic drawbacks associated with cassava. This also arrests the view that African cassava genotypes are recalcitrant to in vitro manipulations.
This work is part of a continuing project on genetic engineering of Kenyan cassava genotypes funded by the Danish International Development Agency (DANIDA). The cassava cultivars used in this study were kindly provided by the Kenya Agricultural Research Institute (KARI). Duncan Ogweda is hereby acknowledged for maintenance of explants sources and regenerated plant