Journal of Thermodynamics & Catalysis
Open Access
ISSN: 2157-7544
ISSN: 2157-7544
Research Article - (2015) Volume 6, Issue 3
The minor groove intercalation loop (leucine-loop) in Uracil-DNA N-glycosylases (UNGs) undergoes significant conformational changes upon substrate interaction. Previous studies have shown that the cold adapted Atlantic cod UNG (cUNG) possesses a tenfold higher catalytic efficiency than human UNG (hUNG). A sequence alignment of the two enzymes revealed amino acid substitutions in and near the leucine-loop of cUNG which may increase the elasticity of the loop and thus explain the observed higher catalytic efficiency of cUNG compared to hUNG. In order to investigate this hypothesis we constructed five single and four multiple mutants of cUNG with cod to human UNG substitutions and characterised them by kinetic experiments at three different temperatures (15, 22 and 37°C) and Differential Scanning Calorimetry (DSC). The results showed that the mutations affected the kcat more than the KM, and mutants with apparent reduced hydrophobic properties (A266T, V267A, A266T/V267A and A266T/ V267A/A274V) showed increased or equivalent kcat to cUNG, while mutants with predicted increased hydrophobicity (A274V, H275Y, L279F, A266T/V267A/A274V/H275Y and A266T/V267A/A274V/H275Y/H250Q) showed reduced kcat compared to cUNG. Thus, mimicking the amino acid composition of the leucine-loop in hUNG reduces the activity of cUNG. The DSC experiments were performed in order to identify a potential increase in the overall stability of the mutants as a result of altered hydrophobicity of the leucine-loop. However, the results showed a slight reduction in the overall stability for mutants with increased kcat and increased stability for mutants with reduced kcat. We suggest these results arise from a need to compensate for a potential increased/reduced elasticity of the leucine-loop which is introduced by the amino acid substitutions. This study confirms the suggested importance of the leucine-loop for efficient catalysis of UNG and shows that it is possible to alter the catalytic capacity of this enzyme by manipulating the amino acid composition of this loop.
Keywords: Cold adaptation of enzymes; DNA repair; Uracil-DNA N-glycosylase; Kinetics; Differential scanning calorimetry
Uracil-DNA N-glycosylase (UNG) is a DNA repair enzyme involved in the base excision repair pathway (BER) for removal of uracil in DNA. Uracil in DNA may result from deamination of cytosine [1] or misincorporation during replication [2]. The crystal structure of the catalytic domain is known from a wide range of organisms including human [3], herpes simplex virus 1 (HSV-1) [4], E. coli [5], Atlantic cod [6], D. radiodurans [7], Vibrio cholerae [8], Vaccinia virus [9], Mycobacterium tuberculosis [10], Bacillus substilis (Banos-Sanz, 2013) and Staphylococcus aureus [11]. All ten enzymes have similar structures and consist of a classic single domain α/β-fold with a central four stranded parallel and twisted β-sheet surrounded by eight to eleven α-helices. The N- and C-terminals are on opposite sides of the central β-sheet and the active site is located within a positively charged groove at the C-terminal end of the parallel β-sheet.
The crystal structures have also been determined of UNG in complex with DNA from human [12-14], Vaccinia virus [15,16] and D. radiodurans [17]. These structures have revealed that the uracil is flipped out of the DNA and into the active site upon detection in a process where the enzyme binds the minor groove, compress the phosphate backbone and kink the DNA [13]. Three Ser-Pro-rich loops are involved in this process (named from the hUNG structure): the 4-Pro loop, the Gly-Ser loop and the minor groove intercalation loop (leucine loop). The four serines in these loops form hydrogen bonds to the phosphates 5’ and 3’ to uracil, and may orient the enzyme correctly for DNA scanning. The initial backbone compression is suggested to be coupled to a minor groove reading head formed by residues in the leucine loop which make water bridged hydrogen bonds to structurally conserved purine N3 sites and widens up the minor groove [13]. When the uracil is detected, it is flipped out of the DNA by further backbone compression, and the resulting DNA stabilised by penetration of a conserved Leucine (Leu272 in hUNG) into DNA minor groove [13]. It is further shown that the leucine plugging of the cavity in DNA increases the lifetime of the extrahelical base [18].
Cold adapted enzymes are characterised by low thermal stability and high catalytic efficiency compared to their mesophilic counterparts. It has been suggested that a consequence of the reduced stability is an increased flexibility which is necessary in order for the enzymes to perform efficient catalysis at low temperatures [19]. No general structural feature seems to account for the increased efficiency and reduced stability of the cold adapted enzymes [20]. However, for some enzymes correlations between cold adaptation and reduced interactions between structural domains or subunits are found. This is observed for salmon trypsin [21], subtilisin from the Antarctic Bacillus TA39 [22], lactate dehydrogenase from Antarctic notothenoid fishes [23], citrate synthase from an Antarctic bacteria [24], phosphoglycerate kinase from Antarctic Pseudomonas sp. TACII18 [25], anionic trypsin from Chum salmon (Oncorhynvhus keta) [26] and alkaline phosphatase from Atlantic cod (Gadus morhua) [27]. These findings suggest local flexibility as a strategy for cold adaptation. Local flexibility is also suggested to be a strategy for cold adaptation of malate dehydrogenase from Aquaspirillium arcticum [28], chitobiase from Arthrobacter sp. TAD20 [29], DNA-ligase from Pseudoalteromonas haloplanktis [30] and alkaline proteinase from an Antarctic Pseudomonas species [31,32] argues that in order to increase the reaction rate of cold adapted enzymes, the best compromise would be to maximise the flexibility in the regions involved in catalysis and maintain or increase the rigidity in regions not involved in catalysis.
Cold adapted UNG has been isolated from the liver of Atlantic cod (Gadus morhua) and characterisation experiments revealed that it possesses cold adapted features, like increased catalytic efficiency and reduced temperature optimum for activity, compared to human UNG (hUNG) [33]. The catalytic domain of cod UNG (cUNG) has been produced (in E. coli) and purified, and characterisation experiments shows that the activity of cUNG is more thermolabile and has a 10- fold increase in catalytic efficiency at optimum conditions (pH 7.0 and 50 mM NaCl) compared to hUNG [34]. The crystal structure of the catalytic domain of cUNG has been determined at 1.9 Å resolution and a comparative analysis of the structures of cUNG and hUNG suggested that a less stable C-terminal part and an increased electrostatic surface potential near the active site might be responsible for the cold adapted behaviour of the cod enzyme [6]. The importance of an optimised positive electrostatic surface potential for the cold adapted features of cUNG has been confirmed experimentally by mutational and structural analysis [35], and theoretical calculation studies have suggested that cUNG in part uses local flexibility of the minor groove intercalation loop (leucine-loop) as a strategy for cold adaptation [6,36]. Recently the first thermal stability analysis of cUNG and hUNG, performed by DSC, was published and shows that the thermal melting temperature (Tm) of cUNG is 9°C lower than for hUNG [37]. Thus the overall stability of cUNG is lower than for hUNG and suggests that it is a more elastic molecule which is able to perform rapid conformational changes during catalysis at low temperatures. Lately the crystal structure and biophysical analysis of the complex of cUNG-Ugi (uracil DNA glycosylase inhibitor) further confirmed the importance of the optimised electrostatic surface potential for the high catalytic efficiency of cUNG compared to hUNG [38].
This study was initiated in order to investigate the hypothesis that a more dynamic leucine-loop of cUNG might contribute to the high catalytic efficiency of cUNG compared to hUNG in addition to the previously observed optimised electrostatic surface potential. Thus, nine mutants of cUNG were constructed, with cod to human UNG amino acid substitutions, and analysed by enzyme kinetics and DSC. In summary our results support the hypothesis that the high catalytic efficiency of the cold adapted UNG can in part be explained by a more dynamic leucine loop compared hUNG. However, the DSC experiments showed an unexpected reduced overall stability of the mutants with increased kcat and increased overall stability for the mutants with reduced kcat. We believe that this effect originates from a need to compensate for the increased/reduced rigidity which is introduced in the leucine-loop by the substitutions.
Materials
E. coli TOP10 [F - mcrA (mrr-hsdRMS-mcrBC) 80lacΖ M15 lacX74 deoR recA1 araD139 (ara-leu) 7697 galU galK rpsL (Str R) endA1 nupG] was purchased from Invitrogen (Carlsbad, CA). E. coli NR8052 [Δ(pro-lac, thi, ara, trpE9777, ung1)], purified recombinant human UNG (UNGΔ84) and the gene encoding the catalytic domain of human UNG was kindly provided by Dr. Hans E. Krokan, Institute for Cancer Research and Molecular Biology, Norwegian University of Science and Technology.
Construction of mutants
The following mutants were constructed: A266T, V267A, Double (A266T/V267A), A274V, Triple1 (A266T/V267A/A274V), H275Y, Leuloop (A266T/V267A/A274V/H275Y), L279F and Triple2 (A274V, H275Y, L279F) The template for construction of the mutants was the gene encoding the catalytic domain of recombinant cod UNG with three additional codons (MEF) and two altered Arg codons (from AGA to CGT) in the N-terminal part of the sequence (cUNGΔ81) [34]. The Triple1, Leuloop and H275Y mutants were constructed as previously described [35]. The A266T, V267A, Double, A274V, L279F and Triple2 were constructed using the QuickChange® Site-Directed Mutagenesis Kit according to the manual from the manufacturer (Stratagene, CA). The primers used for the mutagenesis were as indicated in Table 1. The downstream primers are complementary and reverse of the primers listed in Table 1. The DNA sequencing was performed using BigDye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems).
| Mutant | 5’-primer |
|---|---|
| A266T | CATGTCTTGCAAACCGTTCATCCATCTC |
| V267A | CATGTCTTGCAAGCTGCGCATCCATCTCCTTTG |
| Double (A266T/V267A) | CATGTCTTGCAAACCGCGCATCCATCTCCTTTG |
| A274V | CCTTTGTCTGTGCATCGTGGGTTCCTTGGTTG |
| Triple1 (A266T/V267A/A274V) | GCAAACCGCGCATCCATCTCCTTTGTCTGTGCATCGTG |
| H275Y | GTCTGCTTATCGTGGGTTC |
| Leuloop (A266T/V267A/A274V/H275Y) |
CTTGCAAACCGCGCATCCATCTCCTTTGTCTGTGTATCGTG |
| L279F | CCTTTGTCTGCTCATCGTGGGTTCTTTGGTTG |
| Triple2 (A274V/H275Y/L279F) | CCTTTGTCTGTGTATCGTGGGTTCTTTGGTTG |
Table 1: Primers used for construction of mutants.
Expression
Large-scale expression was performed in 2 L baffled Erlenmeyer flasks using a Multitron Incubator shaker (Infors HT, Bottmingen, Switzerland). Fifty ml pre-cultures of E. coli NR8052 transformed with the pTrc99A plasmid containing genes encoding the mutants were used for inoculation of 0.5 L of LB-medium supplemented with 0.1% glucose and 100 μg/ml of ampicillin. The cells were grown at 37°C until OD600 was approximately 2.0. The expression was induced by adding IPTG to 1 mM concentration 30 minutes after lowering the temperature to 30°C or 20°C and the cells were harvested 5 hours or 16 hours after induction, respectively, by centrifugation.
Protein purification
The protein purification was performed using the Äkta Explorer Purification System (Amersham Biosciences, Uppsala, Sweden). Harvested cells from 4 L culture were re-suspended in 150 ml extraction buffer [25 mM Tris/HCl, 10 mM NaCl, 1 mM EDTA, 1% (v/v) glycerol, pH 8.0] and 4 Complete Mini EDTA-free Protease Inhibitor Cocktail tablets from Roche Applied Science (Indianapolis, USA) dissolved in 4 ml distilled water were added. The cells were disrupted on ice by ultrasonication using Sonics VC Ultrasonic Processor (Sonics & Materials, Inc., Newtown, USA) for one hour (9.9 seconds on, 9.9 seconds off) at 40% amplitude. The cell extract was centrifuged and the supernatant was collected. Thereafter, 2% (w/v) protamine sulfate was added to the final concentration of 0.25% (v/v) and incubated on ice for 15 min with stirring. The solution was centrifuged at 20,000 rpm for 1 hour at 4°C, and the supernatant was collected. The pH of the cell extract was adjusted to 7.5 followed by filtration using a 0.2 μm syringe filter (Millipore, Billerica, USA). The filtrate was then applied onto a Q-Sepharose FF column coupled with an SP Sepharose FF column, both equilibrated in 25 mM Tris/HCl, 10 mM NaCl, 1 mM EDTA, 1% (v/v) glycerol, pH 7.5]. The rest of the purification steps were performed as described above. Fractions containing UNG were pooled and concentrated using Amicon Centrifugal Filter Units (molecular weight cut off, 10 kDa) from Millipore.
Activity measurements
Nick translated calf thymus DNA (Sigma-Aldrich, Steinheim, Germany) with deoxy[5-3H]uridine 5’-triphosphate (Amersham Biosciences, Uppsala, Sweden) was used as substrate. Preparation of substrate and measurement of UNG activity was performed as previously described [33]. The standard assay procedure is in short: The enzyme was diluted in cold dilution buffer (5 mM Tris-HCl, 10 mM NaCl, 1% glycerol, pH 8.0). The activity was measured in a total volume of 20 μl in 70 mM Tris-HCl, 10 mM NaCl, 1 mM EDTA, pH 8.0, 100 μg/ ml BSA and 230 ng substrate (3H-dUMP DNA). The reaction mixture was incubated for 10 minutes at 37°C, and terminated by addition of 20 μl ice cold single stranded calf thymus DNA (1 mg/ml) and 500 μl 10% TCA. After incubation on ice for 15 minutes, free 3H uracil was separated from precipitated material by centrifugation at 16 000 g for 10 minutes, and analysed using a liquid scintillation counter. One unit enzyme is defined as the amount of enzyme required for release of 1 nmol acid soluble uracil per minute at 37°C.
Kinetics
KM and kcat were measured in the presence of eight different [3H]- dUMP substrate concentrations in the range of 0.56 -4.5 μM at 15°C, 22°C and 37°C. The amounts of UNG used were different for each mutant at the different temperatures, giving cpm values between 500 - 5000. Assays were performed in 25 mM Tris-HCl, 50 mM NaCl, 1 mM EDTA and 100 μg/ml BSA, pH 7.5 at 37°C. The calculation of the kinetic constants was performed by the enzyme kinetics module in SigmaPlot (SPSS Inc., Chicago, IL).
Differential scanning calorimetry (DSC)
DSC experiments were conducted on a Nano-Differential Scanning Calorimeter III, model CSC6300 from Calorimetry Sciences Corporation (Lindon, USA). Preparations of the native enzymes were dialyzed overnight at 4°C against 1 L of dialysis buffer [25 mm Hepes, 10 mM NaCl, 1 mM EDTA, 1% (v/v) glycerol, pH 7.4) in a Pierce Slide- A-Lyzer dialysis cassettes from Thermo Fisher Scientific Inc (Rockford, USA) with a 10 kDa cut-off followed by filtration using a 0.2 μm syringe filter (Millipore, Billerica, USA).
The dialysis buffers were used as reference buffers in the DSC runs. Reference buffers and samples were degassed for 10 minutes before loading into the DSC cells. The temperature scans were carried out at a constant pressure of 3 atm in a range 10 or 20 to 70°C with a heating or cooling rate of 1°C /min. Refolding experiments were performed for each protein by a backward cooling scan after unfolding and by incubating the unfolded proteins at 4°C for 1 hour before initiating a new forward scan. CpCalc software supplied with the instrument was utilized to generate molar heat capacity values. The experimental transition curves were fitted on a two-state transition model.
The minor groove intercalation loop (leucine-loop) undergoes significant conformational changes upon binding of UNG to substrate DNA and brings His268 within hydrogen binding distance of the uracil O2, to accomplish the formation of the uracil recognition pocket. It also stabilises the distorted DNA when uracil is flipped into the specificity pocket [13,17,18] It is thus important that the loop consists of amino acids which allow rapid conformational changes upon interaction with DNA. A sequence alignment of cod and human UNG (Figure 1) revealed differences in the amino acid composition close to and within the leucine-loop which suggest improved dynamic properties of the cUNG loop.
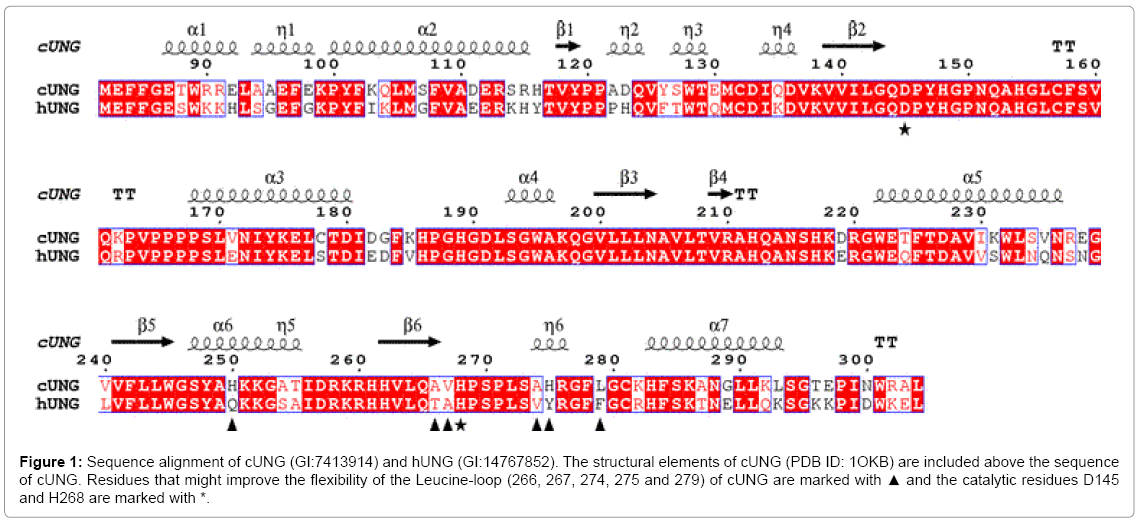
Figure 1: Sequence alignment of cUNG (GI:7413914) and hUNG (GI:14767852). The structural elements of cUNG (PDB ID: 1OKB) are included above the sequence of cUNG. Residues that might improve the flexibility of the Leucine-loop (266, 267, 274, 275 and 279) of cUNG are marked with ▲ and the catalytic residues D145 and H268 are marked with *.
Two of the residues which anchor the leucine-loop to the β4 strand in hUNG, Thr266 and Ala267, is substituted by Ala and Val in cUNG. In addition are Val274, Tyr275 and Phe279 substituted by Ala, His and Leu in cUNG, respectively. Based on this observation, nine different mutants with both single and multiple substitutions were constructed and characterised with regard to catalytic efficiency and temperature stability (Tables 2 and 3). The positions of the substituted amino acids are shown in Figure 2.
| UNG | T(°C) | Vmax(nmol/min/mg) | kcat(min-1) | KM(μM) | kcat/KM(min-1μM-1) | ||
|---|---|---|---|---|---|---|---|
| cUNG[35] | 37 | 49135 | ± 1626 | 1241 | 0.8 | ± 0.09 | 1499 |
| 22 | 24439 | ± 555 | 617 | 0.8 | ± 0.06 | 795 | |
| 15 | 11064 | ± 374 | 279 | 0.5 | ± 0.07 | 607 | |
| A266T | 37 | 53467 | ± 1079 | 1352 | 0.6 | ± 0.04 | 2282 |
| 22 | 24994 | ± 464 | 632 | 1.0 | ± 0.05 | 641 | |
| 15 | 10006 | ± 170 | 253 | 0.7 | ± 0.04 | 389 | |
| V267A | 37 | 47476 | ± 1363 | 1197 | 0.5 | ± 0.06 | 2507 |
| 22 | 15218 | ± 313 | 384 | 0.6 | ± 0.04 | 685 | |
| 15 | 5867 | ± 113 | 148 | 0.3 | ± 0.03 | 500 | |
| Double (A266T/V267A) | 37 | 68898 | ± 1455 | 1740 | 0.7 | ± 0.05 | 2500 |
| 22 | 20891 | ± 320 | 528 | 0.8 | ± 0.04 | 686 | |
| 15 | 10805 | ± 249 | 273 | 0.5 | ± 0.05 | 549 | |
| A274V | 37 | 37379 | ± 611 | 944 | 1.1 | ± 0.05 | 830 |
| 22 | 11746 | ± 244 | 297 | 0.8 | ± 0.06 | 381 | |
| 15 | 8991 | ± 286 | 227 | 1.3 | ± 0.11 | 174 | |
| Triple1 (A266T/V267A/A274V) |
37 | 61843 | ± 2879 | 1562 | 0.7 | ± 0.12 | 2199 |
| 22 | 30544 | ± 733 | 771 | 0.8 | ± 0.07 | 929 | |
| 15 | 14550 | ± 509 | 367 | 0.7 | ± 0.09 | 496 | |
| H275Y [35] | 37 | 23590 | ± 859 | 596 | 0.6 | ± 0.09 | 1017 |
| 22 | 11974 | ± 553 | 302 | 0.6 | ± 0.11 | 509 | |
| 15 | 6166 | ± 384 | 156 | 0.7 | ± 0.16 | 224 | |
| Leuloop (A266T/V267A/A274V/H275Y) |
37 | 26603 | ± 1293 | 672 | 0.7 | ± 0.12 | 1018 |
| 22 | 10415 | ± 282 | 263 | 0.5 | ± 0.06 | 548 | |
| 15 | 6007 | ± 273 | 152 | 0.5 | ± 0.11 | 292 | |
| L279F | 37 | 31674 | ± 321 | 800 | 0.5 | ± 0.02 | 1459 |
| 22 | 11944 | ± 145 | 302 | 0.5 | ± 0.03 | 611 | |
| 15 | 7977 | ± 252 | 201 | 0.6 | ± 0.08 | 342 | |
| Triple2 (A274V/H275Y/L279F) |
37 | 30569 | ± 676 | 713 | 0.8 | ± 0.06 | 919 |
| 22 | 7551 | ± 160 | 176 | 0.8 | ± 0.05 | 215 | |
| 15 | 4352 | ± 79 | 102 | 0.7 | ± 0.04 | 146 | |
| hUNG[35] | 37 | 25347 | ± 1256 | 647 | 2.1 | ± 0.23 | 309 |
| 22 | 12530 | ± 740 | 320 | 2.4 | ± 0.29 | 135 | |
| 15 | 5580 | ± 399 | 142 | 2.2 | ± 0.34 | 66 | |
Table 2: Kinetic constants of the interactions of cUNG and mutants with substitutions in the Leucine-loop with uracil (3H+ labelled) incorporated calf thymus DNA. The assay was performed at 15, 22 and 37°C in 25 mM Tris/HCl (pH 7.5 at 25°C), 50 mM NaCl, 1 mM EDTA. The Vmax and Km were extracted by fitting the velocity data obtained at 8 different substrate concentrations on the Michaelis-Menten equation using the enzyme kinetics module in Sigma Plot. The standard error (SD) for the fit is indicated.
| UNG | 37°C | 22°C | 15°C |
|---|---|---|---|
| cUNG | 0 | 0 | 0 |
| A266T | -9 | -2 | 9 |
| V267A | 4 | 38 | 47 |
| Double (A266T/V267A) | -40 | 14 | 2 |
| A274V | 24 | 52 | 19 |
| Triple1 (A266T/V267A/A274V) | -26 | -25 | -32 |
| H275Y | 52 | 51 | 44 |
| Leuloop (A266T/V267A/A274V/H275Y) |
46 | 57 | 46 |
| L279F | 36 | 51 | 28 |
| Triple2 (A274V/H275Y/L279F) | 43 | 71 | 63 |
| hUNG | 48 | 48 | 49 |
Table 3: Reduction (%) in kcat at different temperatures for cUNG mutants and hUNG relative to cUNG.
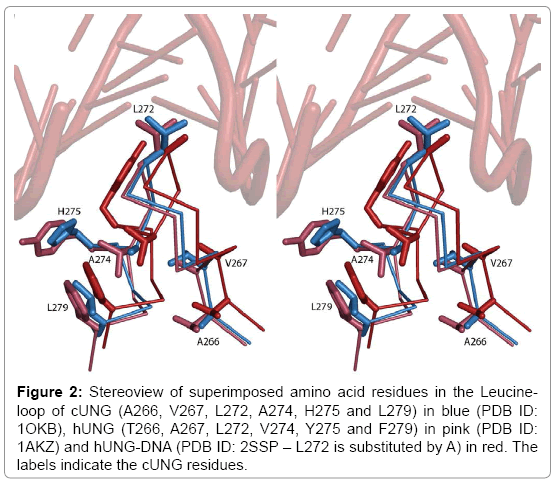
Figure 2: Stereoview of superimposed amino acid residues in the Leucineloop of cUNG (A266, V267, L272, A274, H275 and L279) in blue (PDB ID: 1OKB), hUNG (T266, A267, L272, V274, Y275 and F279) in pink (PDB ID: 1AKZ) and hUNG-DNA (PDB ID: 2SSP – L272 is substituted by A) in red. The labels indicate the cUNG residues.
The mutants were produced in large scale and purified following the procedures described earlier and diafiltrated. The yield varied, but was typically between 2-4 mg for each mutant. After purification, the kinetic constants, kcat and KM were determined for the mutants at 15°C, 22°C and 37°C using the optimal buffer conditions for cUNG (pH 7.5 and 50 mM NaCl). In addition, thermal stability was measured by DSC.
The A266T, V267A and A266T/V267A mutants shows similar and higher kcat compared to cUNG
An analysis of the enzyme kinetics shows that the mutations affected the kcat (turnover rate) more than the Michaelis-Menten constant (KM), and in the case of the A266T mutant, the kcat is similar to cUNG at all three temperatures and only slightly increased at 37°C. For the A267V mutant the kcat value is similar to cUNG at 37°C, while it is reduced with approximately 50% at 22 and 15°C (Table 3). However, when these two mutations are combined in the A266T/V267A double mutant, the kcat is increased with 40% compared to cUNG at 37°C, while it is slightly decreased with 14 and 2% compared to cUNG at 22 and 15°C, respectively (Table 3).
As mentioned above, A266 and V267 anchor the leucine-loop to the β4 strand in UNG and the substitutions to Thr and Ala, respectively, probably allow more elastic movements of the leucine-loop which give rise to a more efficient interaction with the damaged DNA. Why the kcat is reduced by approximately 50% for the V267A mutant compared to cUNG at lower temperatures is difficult to explain. However, since electrostatic interactions are more strengthened at low temperatures, substituting the Valine with Alanine may affect one of the two observed hydrogen bonds between His268 and Gln144 or Leu272 and Arg276 in the leucine-loop in such a way that they get stronger, and thus reduces the dynamic properties of the loop and the kcat at 22 and 15°C.
The H275Y substitution severely affects the kcat
The kcat of the V274A, H275Y and L279F mutants are strongly reduced compared to cUNG with most effect on the H275Y mutant which has an approximate reduction of 50% in kcat at all temperatures (Table 3), and indicates that His275 is very important for the high catalytic efficiency of cUNG. This has previously been observed through theoretical calculation studies [36], where one of the experiments showed that the side chain of Arg276 is closer to DNA in a cUNG-DNA complex than in hUNG-DNA, which is caused by a hydrogen bond between His275 and the O´5 atom on the adenine base opposite of the uracil pulling the DNA strand closer to the enzyme. From the structure of cUNG, it does not seem like His275 is able to interact strongly with DNA, since it is pointing out from the minor groove of DNA (Figure 2). However the crystal structure of hUNG in complex with DNA, shows that the side chain of Tyr275 has changed conformation upon binding of DNA, pointing into the minor groove of DNA (Figure 2), and even though the crystal structure of cUNG in complex with DNA has not been determined, we assume that the same will happen with His275. This movement was observed in the model made of cUNG in complex with DNA in the theoretical study [30].
Thus His275 in cUNG probably interacts more strongly with DNA compared to Tyr275 in hUNG, and explains why the catalytic efficiency is severely reduced for the H275Y mutant (Table 2). However, the H275Y mutant shows no significant difference in KM compared to cUNG, rather the reduction in catalytic efficiency is caused by a reduction in kcat. One explanation for this could be that the His275 residue binds stronger to the transition state than to the ground state, affecting kcat instead of KM [39]. A strong interaction between the leucine-loop in cUNG and DNA is supported by a mutational study of Arg276 in hUNG, which shows its importance for the activity of hUNG due to favourable interactions with DNA [40].
The multiple mutants confirm the importance of the H275Y substitution
When we combine the A274V, H275Y and L279F mutations in the A274V/H275Y/L279F mutant, the kcat value is even more reduced than for the H275Y mutant at 22 and 15°C, showing an approximate reduction of kcat of 70 and 60% compared with cUNG, respectively, while it is slightly higher than for the H275Y single mutant at 37°C (Tables 2 and 3). The main contributor to the reduced kcat is obviously the H275Y mutation, however it seems like the two other mutations has additional effects. Looking into the structures of cUNG and hUNG again (Figure 2), it seems likely that Val274, Tyr275 and Phe279 generates a hydrophobic cluster in the leucine-loop which may rigidify the loop. The equivalent residues in cUNG are Ala274 and Leu279 in addition to His275 which probably allow more elastic movements of the loop and thus a more efficient interaction with DNA (Figure 2). The V274, Y275 and F279 in the A274V/H275Y/L279F mutant probably generate a similar hydrophobic cluster in the leucine-loop, thus making the loop more rigid and preventing it from rapid conformational changes upon binding to DNA. Results from a molecular dynamic (MD) simulation of cUNG supports this theory showing reduced dynamic properties of the leucine-loop in hUNG compared to cUNG [36]. The same study showed that substituting only His275 with Tyr results in a reduced flexibility of the leucine-loop compared to native cUNG.
The high kcat of the A266T/V267A/A274V mutant is difficult to explain but might be caused by an increased elasticity which is allowed by the A266T and V267A. However, by adding the H275Y substitution in the A266T/V267A/A274V/H275Y mutant, the kcat is reduced. As seen before, the H275Y single mutant has severe effect on kcat of cUNG alone, and is probably causing the kcat to drop also for this multiple mutant, due to less efficient interactions with the DNA.
The substitutions affect the stability of cUNG in an unexpected manner
In order to analyse whether the amino acid substitutions in the leucine-loop had any effect on the overall stability of the protein, Tm was determined of the mutants by DSC. For every experiment in the order of 1-2 mg of protein was used, and all the measurements were performed by increasing the temperature by 1°C every minute until the melting curve was completed. The reversibility of the unfolding was tested by slowly cooling down the protein solution in the sample cell and re-running the DSC experiment, however we were not able to detect any new melting curves, thus the unfolding was irreversible. This was confirmed by visual inspections of the protein solutions after the experiment which showed white protein aggregates, and is in accordance with previous results from thermal stability analysis of cUNG and hUNG [37].
The results from the DSC experiments show that Tm was only slightly decreased or increased for the mutants compared to cUNG (Figure 3). The mutants with Tm lower than cUNG were A274V (43.4°C), H275Y (43.8°C), Triple2 (44.0°C), Leuloop (44.4°C) and Triple1 (44.6°C) while mutants with higher Tm were Double (45.3°C), A266T (46.1), L279F (46.5°C) and V267A (46.5°C) (Table 4). The differences in Tm between native cUNG and the mutants is not large, with only a 1.7 and 1.3°C reduction in Tm for the A274V and H275Y substitutions respectively and a 1.0 and 1.6°C increase for the A266T and V267A substitutions. Such subtle changes in Tm caused by mutations analyzed by DSC is a common phenomenon and have been reported for other cold adapted enzymes like α-amylase from Pseudoalteromonas haloplanktis (AHA) [41] and chitinase from the Antarctic Arthrobacter Sp. TAD20 [42].
| UNG | ΔHcal (kJ/mol) | Tm (°C) |
|---|---|---|
| cUNG | 351 | 45.1 |
| A266T | 564 | 46.1 |
| V267A | 406 | 46.7 |
| Double (A266T/V267A) | 489 | 45.3 |
| A274V | 443 | 43.4 |
| Triple1 (A266T/V267A/A274V) | 472 | 44.6 |
| H275Y | 460 | 43.8 |
| Leuloop (A266T/V267A/A274V/H275Y) | 448 | 44.4 |
| L279F | 420 | 46.5 |
| Triple2 (A274V/H275Y/L279F) | 350 | 44.0 |
Table 4: Thermodynamic stability parameters of native and mutant cUNG in Hepes buffer by DSC. The heating rate was 1°C per min. The experiments were performed at least twice, and resulted in reproducible Tm values irrespective of the protein concentration used. The molar heat capacity data extracted from experimentally obtained transition curves were deconvoluted on a two-state transition model using CpCalc software. The unfolding was irreversible.
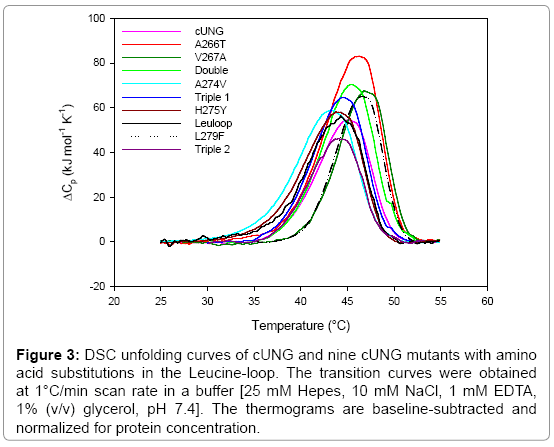
Figure 3: DSC unfolding curves of cUNG and nine cUNG mutants with amino acid substitutions in the Leucine-loop. The transition curves were obtained at 1°C/min scan rate in a buffer [25 mM Hepes, 10 mM NaCl, 1 mM EDTA, 1% (v/v) glycerol, pH 7.4]. The thermograms are baseline-subtracted and normalized for protein concentration.
Our hypothesis for this experiment was that the substitutions introduced in the leucine-loop would alter the rigidity of the loop and increase or reduce the overall stability accordingly. According to the hypothesis about an inverse relationship between activity and stability [23], we expected to observe reduced stability of the mutants with increased elasticity, and increased stability of the mutants with reduced elasticity. However, as observed from the results, this is not the case. Instead the mutants with increased hydrophobicity (A274V, H275Y, Triple1, Triple2 and Leuloop) possessed reduced stability, while mutants with reduced hydrophobicity (A266T, V267A, Double and L279F) possessed increased stability. Thus, the mutations affected the overall stability of the protein, however it seems like the changes can only be related to overall instead of local structural rearrangements which occur due to the substitutions and has to be evaluated separately from the changes in activity which are observed of the mutants.
Free energy (ΔG) of hydration explains unexpected stability results
The free energy (ΔG) of hydration is generally unfavourable for hydrophobic amino acids in contact with water at higher temperatures and destabilises proteins [43]. This effect is probably causing the reduced thermostability which is observed of the A274V, H275Y, Triple1, Triple2 and Leuloop mutants (Figure 3).
This theory also counts for the observed increased stability of the A266T, V267A and Double mutant, which probably is caused by the less unfavourable effect the reduced hydrophobicity has on the free energy (ΔG) of hydration.
Some of the activity measurements have been performed at 37°C, and as we can see from the DSC experiments (Figure 3), it seems like the unfolding process has started for both A274V and H275Y at temperatures below this temperature. Based on this result it is tempting to explain the reduced kcat of these mutants with reduced overall stability at 37°C instead of an increased rigidity of the leucine-loop. However, the activity measurements are performed in the presence of DNA, which most likely increase the overall stability of the protein at the same time as it induce conformational changes of the loops and residues in the loops (e.g., Tyr275 in hUNG) which are involved in catalysis [12,13]. This is also well supported by results from a heat inactivation experiment we previously performed of the H275Y mutant, which showed an apparent increased stability of the H275Y mutant compared to cUNG upon substrate binding [35].
The ΔHcal is increased for all the mutants compared to cUNG except Triple2, indicating that an increased amount of energy is needed to break the intramolecular interactions during the unfolding process of the proteins and thus an enhanced stability of the mutants. These results contradict the Tm values showing that several of the mutants possess reduced stability compared to cUNG. However, ΔHcal is also a measure of energy which takes into consideration hydration effects upon unfolding in addition to bond breaking, and due to the aggregation of the protein we would thus be careful to draw any conclusion regarding the ΔHcal of our mutants.
Stability/flexibility and activity relationships
The high catalytic activity of cold adapted enzymes is explained by an increased flexibility (and thus reduced stability) of the proteins which will allow rapid conformational changes at low temperatures [23]. There is growing evidence based on mutagenesis studies that increased catalytic activity of cold adapted enzymes is not always associated with reduced stability, and reduced stability is not always indicative of increased molecular flexibility [44]. Such delicate balance between activity and stability have been reported for subtilisin S41 from the Antarctic Bacillus TA41 [45], chitinase from the Antarctic Arthrobacter Sp. TAD20 [42] and alkaline phosphatase (AP) from the Antarctic strain TAB5 [46]. This has also been demonstrated in our study which shows that variants with reduced kcat compared to the wild type enzyme were not found to be more stable, and vice versa. A274V, H275Y, Leuloop, Triple1 and Triple2 were characterized by reduction of both enzymatic activity and stability while an improvement of both parameters was observed for A266T, V267A and Double. The only mutant which behaves according to the simple inverse relationship between activity and stability is the L279F mutant who possesses a 1°C increase in Tm and a 40% reduction in activity at 37°C compared to cUNG. Since we do not have the structure of this mutant, the explanation for this behaviour will only be speculative, but through its location at the edge of the leucine-loop (Figure 2) it might generate stabilising interaction with His275 in the loop making it less prone to hydration by surrounding water molecules, and increase the stability of the leucine-loop and thus the overall stability of the enzyme. The features of the Triple1 mutant are also different from the others by possessing both increased stability and activity. A similar result is observed for a mutant of subtilisin from the Antarctic bacillus TA39 [22] and reveals interesting options for industrial application of cold adapted enzymes.
Through theoretical calculation studies it has been suggested that the leucine-loop is more flexible in cUNG than in hUNG, which in combination with an optimized electrostatic surface potential, probably explains the increased catalytic efficiency of cUNG compared to hUNG [36]. We have through this study shown that an increased hydrophobicity of the leucine-loop reduces the activity of cUNG. However, the expected increased rigidity of the loop is not reflected in an increased overall stability, instead showed mutants with expected increased stability reduced stability and vice versa. This corresponds to the findings published by suggesting that a maximised flexibility in regions involved in catalysis in combination with an increased overall rigidity in other parts of the protein is a good compromise for increasing the reaction rate of cold adapted enzymes [32]. Thus, we propose that the increased hydrophobicity which has been introduced in the leucine-loop mutants resulted in an increased rigidity of the loop which has been compensated for by reducing the overall stability of the protein and vice versa.
The minor groove intercalation loop (leucine-loop) is very important for efficient removal of uracil in DNA by bringing the catalytic residue His268 within hydrogen binding distance of uracil, accomplishing the formation of the substrate binding pocket and stabilising distorted DNA when uracil is flipped into the specificity pocket [13,18]. Here, we have studied how substitutions of hydrophobic residues close to and in the leucine-loop affect the activity and overall stability of Atlantic cod UNG (cUNG). Based on our results we suggest that the reduced hydrophobicity of the leucine-loop in cUNG, increases its dynamic properties and thus optimizes the interaction between the leucine-loop and damaged DNA compared to hUNG. Our results confirm the importance of the leucine-loop for catalysis by UNG, and show how the capacity of this enzyme can be altered by manipulating the amino acid content of this loop.
We thank the Research Council of Norway, the National Functional Genomics Program (FUGE) and the University of Tromsø for funding this project.