Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2016) Volume 4, Issue 2
The aim of this study evaluated the in vitro and in vivo antimicrobial activity of selected lactic acid bacteria (LAB) against UPEC for prevention and amelioration of UTIs. We screened LAB strains with antimicrobial effects on UPEC using a well-diffusion assay, bacterial adherence to the uroepithelium cell line SV-HUC-1 (BCRC 60358), and a co-culture inhibition assay. The results showed that the 7 LAB strains (Lactobacillus paracasei, L. salivarius, two Pediococcus pentosaceus strains, two L. plantarum strains, and L. crispatus) and the fermented probiotic products produced by these multi-LAB strains exhibited potent zones of inhibition against UPEC. Moreover, the LAB strains and probiotic products adhered strongly to the uroepithelium SV-HUC-1 cell line. The growth of UPEC strains was also markedly inhibited after co-culture with the LAB strains and probiotic products in human urine. In addition, the enhanced levels of IL-6, IL-8 and lactic acid dehydrogenase were significantly decreased by treatments with the LAB strains and probiotic products in UPEC-induced SV-HUC-1 cells. Furthermore, oral administration of probiotic products reduced the number of viable UPEC in the urine of UPEC-challenged BALB/c mice. Taken together, this study demonstrates that probiotic supplementation may be useful as an adjuvant therapy for the treatment of bacterial-induced urinary tract infections.
Keywords: Lactic acid bacterium; SV-HUC-1 uroepithelium; Urinary tract infection; Uropathogenic Escherichia coli ; BALB/c mice
ELISA: Enzyme-linked immunosorbent assay; IL: Interleukin; LAB: Lactic acid bacteria; LDH: Lactic acid dehydrogenase; SCS: Spent culture supernatant; UTI: Urinary tract infections; UPEC: Uropathogenic Escherichia coli
Urinary tract infections (UTI) are one of the most prevalent bacterial infections in humans and are a major cause of morbidity [1]. The lifetime risk for acquiring a symptomatic UTI is approximately 50% in women and 12% in men, with a rate of recurrence after six months of approximately 40% [2]. UTIs usually start as bladder infections (cystitis), but can develop into acute kidney infections (pyelonephritis), ultimately resulting in scarring and renal failure. UTIs are caused by a range of pathogens, with uropathogenic Escherichia coli (UPEC) being the most common UPEC (more than 80% of all UTIs) etiological agent [3]. UPEC strains possess harmful factors including fimbrial adhesins, toxins, flagella, auto transporter proteins and iron-acquisition systems, and that contribute to cause clinical diseases [4]. It has been recognized that UPEC can invade host uroepithelial tissue, contributing significantly to the pathogenesis of UTIs by escaping a great number of antibiotics [2]. UPEC can also suppress the innate immune response via expression of specific virulence-associated proteins that result in clinical symptoms [5].
The use of low-dose once-daily or post-coital antimicrobials has been a cornerstone for the treatment and prevention of UPEC-related UTIs, including recurring infections [6]. However, even with urine concentrations of antibiotics far exceeding minimal inhibitory concentrations, UPEC reservoirs in tissues were not effectively eradicated [7]. Therefore, alternative non-antimicrobial based therapeutic approaches such as probiotics that inhibit bacterial adherence and colonization may be of benefit. In our previous studies, probiotics were employed to prevent some pathogenic infections, such as Salmonella , Helicobacter pylori and Escherichia coli [8-10]. The use of probiotics and fermented milk products to combat UTIs has also been previously assessed [11]. However, while there is evidence that lactobacilli have an effect on UTIs, their mechanism of action has thus far not been elucidated, with most studies relying on circumstantial evidence [12-14]. In all, bio-surfactants, bacteriocins, lactic acid and hydrogen peroxide from Lactobacillus sp. seem to be inhibitory for UPEC growth by adversely affecting fimbrial structure and adhesion and up regulating immunogenic membrane proteins [15,16]. The aim of this study was to assess the in vitro and in vivo antimicrobial/ bacteriostatic activity of selected lactic acid bacteria (LAB) against UPEC for the prevention and ameliorating of UTIs.
Bacterial strains and culture condition
A total of 366 LAB strains were isolated from fermented vegetables. The basic screening procedures of the probiotic stains were including antimicrobial activity, acid/bile tolerance and adhesion studies. The stock culture collection was maintained at -80°C in 20% glycerol. Bacterial cells were propagated twice in Lactobacilli MRS broth (Difco, Detroit, MI, USA) containing 0.05% L-cysteine by incubation at 37°C for 24 h. Spent culture supernatant (SCS) was obtained by centrifugation (1000 × g for 10 min at 4°C) of 20 h old LAB culture adjusted to N (N=1-9) × 109 CFU per mL. The sterility of the SCS was verified by plating on MRS agar. Because a pH ranging from 4 to 4.5 was observed for different SCSs, the pH of the cultures was adjusted to 4.5 with 1N NaOH to standardize the condition. Bacteria used for the inhibition study were uroseptic Escherichia coli strains (E. coli BCRC10675, E. coli BCRC15479 and E. fergusonii BCRC 15585) from the Bioresources Collection and Research Center (BCRC, Food Industry Research and Development Institute, Hsinchu, Taiwan). The UPEC strains were propagated in nutrient broth (Detroit, MI, USA) for 18 h at 37°C.
Cell culture
The human urothelial cell line SV-HUC-1 (BCRC 60358) was purchased from the BCRC. Cells were grown in Ham's F12 medium and supplemented with 10% FBS at 37°C in a humidified atmosphere of 95% air and 5% CO2. The culture medium was renewed each day. Cells were sub cultured weekly with 0.1% trypsin and 10 mM EDTA in PBS.
The inhibition zone of UPEC growth caused by LAB strains
This experiment was performed according to the method described by Rammelsberg and Radler [17]. The UPEC strains were grown in nutrient broth overnight and diluted to 106 CFU/mL then spread onto nutrient agar. One hundred microliter of the 18 h LAB culture supernatant (1000 × g, 10 min) was dropped into the well on the nutrient agar, and the plates were incubated at 37°C for 12–14 h. The inhibition zone diameters were then measured. LAB strains whose supernatants induced inhibition zones <1, 11–16, 17–22 and >23 mm were classified as strains inducing no−; mild+; strong++; and very strong+++ inhibition, respectively.
LAB resistance to simulated gastrointestinal conditions
One milliliter of culture containing approximately 109 CFU/mL of LAB was transferred into 9 mL phosphate-buffered saline (PBS). The pH was adjusted to 2.0, 3.0 and 7.0 using 0.1 N HCl, and the solution was incubated at 37°C for 3 h. Controls were performed at pH 7.2 under the same conditions and showed no change in LAB viability. After incubation, viable bacterial counts were determined by serially diluting the culture in PBS (pH 7.2) and plating on MRS agar. Acid tolerance was determined by counting the number of viable LAB after incubation at pH 2.0, 3.0 and 7.0 for 3 h. Plates were incubated anaerobically at 37°C for 48 h.
LAB that survived the acid tolerance study (pH, 3 h) were centrifuged (1000 × g, 10 min), washed with PBS (pH 7.2), and then grown in 9 mL MRS broth with or without 0.15% (w/v) oxgall bile (Sigma-Aldrich, St. Louis, MO, USA) for 0, 1.5 and 3 h, as described by Gilliland and Walker [18]. Bile tolerance was determined by comparing the number of viable LAB grown in MRS with and without bile salt.
The Epithelial cell line culture and adhesion assay
The SV-HUC-1 cell line was cultured in 75 cm2 plastic tissue culture flasks (GIBCO). The cells were washed twice with PBS and then transferred (8 × 104 cells/mL) with 0.05% trypsin into a 24-well multidish containing fresh culture medium without penicillin or streptomycin. The mixtures were kept at 37°C in 5% CO2/95% air atmosphere until cell lines formed a monolayer in each well. Prior to the adhesion test, all the bacterial strains were washed twice with PBS and centrifuged for 10 min at 1000 × g. Bacterial cells were resuspended in 1 mL Ham's F12 medium. One hundred microliters of the suspension were transferred to the 24-well multidish (1 × 108 CFU/mL) and incubated for 2 h at 37°C in 5% CO2. After incubation, cells were washed twice with PBS, fixed with 6% formalin for 30 min, washed twice with PBS and then stained with crystal violet for 5 min. The numbers of LAB cells adhered to the cultured cells were enumerated according to the method described by Gopal et al. [19].
Antimicrobial testing
The viability of UPEC strains was determined using the method described to Chapman, et al. [20]. Briefly, 18-h-old UPEC (1 × 109 CFU/mL) cultures were resuspended in PBS and centrifuged at 1000 × g for 10 min at 4°C. The supernatant was discarded, and the cell pellet was washed once with sterile PBS and re suspended in nutrient broth. Colony count assays were performed by incubating 1 mL of a UPEC (107 CFU/mL) culture mixed with 1 mL LAB-SCS (109 CFU/mL) and 4 mL MRS broth. At predetermined time intervals, aliquots were removed, serially diluted and plated on Mac-Conkey agar (37°C, 18 h) to determine the bacterial colony counts.
Taxonomic identification for LAB strains
In addition to the phenotypic tests, genetic identification methods were performed. Screened LAB strains were identified through Gram staining, morphology examination, and identification with an API 50 CHL kit (La Balme Les Grottes, Montalien, Jeraen, France). In addition to the above processes, we also performed 16S rDNA sequence alignment and fatty acid composition analysis. Furthermore, the GC content of LAB strains were analyzed, and DNA hybridization assays were performed using total DNAs from type strains of specific LAB strains as probes.
The LAB fermented conditions
First, LAB strains were screened for the strongest antimicrobial and adhesion activities, and then these LAB strains were combined to produce LAB fermentation products. The fermentation process was authorized to New Bellus Enterprise Co., Ltd. (Tainan, Taiwan). The LAB fermented products were named PP366 (Strain PM68), PP365 (Strain PM201), LP142 (Strain PM206), Yi Sheng Mi, Shu Mi Chang and Yi Sheng Mi plus. The products PP366, PP365 and LP142 were single-strain probiotics, and Yi Sheng Mi, Shu Mi Chang and Yi Sheng Mi plus were multi-strain probiotics. The LAB fermented products were subsequently tested for their antimicrobial and anti-adhesion properties.
Urothelial cell culture and challenge
The urothelial cell challenge was performed according to the method described by Karlsson and Jass [21]. Briefly, the SV-HUC-1 cells (6 × 104 cells/mL) were seeded in 24-well tissue culture plates and allowed to attach for at least 48 h before addition of bacteria. After incubation, cells were washed twice with PBS, and then 480 μL of fresh culture medium (without penicillin–streptomycin) was added. In a 24- well format, 10 μl of the LAB suspension (1 × 109 CFU/mL) and 10 μl of the heat-killed UPEC (1 × 109 CFU/mL) were added to wells containing urothelial cells. All challenges were performed in triplicate and for 24 h. Supernatants, to be used for ELISA, were stored at -20°C until cytokine analysis.
Enzyme-linked immunosorbent assay
SV-HUC-1 cells were cultured in a 24-well tissue plate and incubated in medium with or without treatment for 24 h. The conditioned medium was collected to assay the levels of interleukin-6 (IL-6) (BD Biosciences, CA, USA) and IL-8 (BD Biosciences, CA, USA) secretion following the manufacturer’s instructions for each ELISA kit.
Detection of lactate dehydrogenase levels
SV-HUC-1 cells (1 × 104 cells/mL) were co-incubated with UPEC and LAB strains for 1 h, and the lactate dehydrogenase (LDH) levels in the supernatants were analyzed using the CytoscanTM-LDH Cytotoxicity Assay (G-Biosciences, MO, USA). The analyses were performed according to protocol provided by the manufacturer.
The viability of UPEC, LAB strains and LAB-fermented products in human urine
The UPEC, LAB strains and LAB-fermented product viabilities in human urine was performed using the method described by Chapman et al. [20]. Urine was collected from healthy human volunteers, passed through a 0.22 μm filter and stored at 4°C for subsequent experiments. The 18-h-old UPEC (1 × 109 CFU/mL) and LAB strains (1 × 109 CFU/mL) were resuspended in PBS. Colony count assays were performed by incubating 1 mL UPEC (107 CFU/mL) culture mixed with 1 mL LAB (109 CFU/mL) and 8 mL human urine. At predetermined time intervals, aliquots were removed, serially diluted and plated on Mac Conkey agar and MRS agar (37°C, 18 h) to determine bacterial colony counts.
Murine model of UTI
Ten-week-old female BALB/c mice were purchased from BioLASCO, Taipei, Taiwan. The mice were divided into eight groups (six mice in each group) and were housed in individual ventilated cages in an air-conditioned room at 23 ± 1°C, 55 ± 5% relative humidity, with a 12 h light/dark cycle. The mice were maintained on a 1324 SPF 10 mm diet (Altromin, Germany) and water was available ad libitum throughout the experimental period.
The induction of UTI was performed according to the method described by Asahara et al. [22]. The mice were anesthetized by administration of isoflurane. After sterilization of the periurethral area with 70% ethanol, a sterile 24-gauge intravenous catheter (inner diameter, 0.47 mm; length, 19 mm; Terumo Corporation, Philippines) was inserted into the bladder through the urethra. Before inoculation with bacteria, the bladder mucosa was traumatized by infusing 50 μL of 0.1N HCl solution for 45 s, followed by neutralization with 50 μL of 0.1 N KOH and flushing with sterile saline. A 20-μL inoculum containing 1 × 108 CFU/mL of E. coli BCRC10675 (resuspended in sterile saline) was then infused into the bladder through a catheter over 30s, and drinking water was given to mice in 1 h after bladder mucosa traumatization. Six groups of mice were orally administered 130 mg/kg bw/day of the fermented probiotic products (PP366, PP365, LP1, Yi Sheng Mi, Shu Mi Chang or Yi Sheng Mi plus) for 14 consecutive days after induction of UTI. The UPEC group (orally administered sterile saline) received the same sensitization schedule, and the vehicle group was treated with sterile saline.
Collection and enumeration of UPEC from the urine of BALB/c mice
For UPEC enumeration, the urine from mice was collected, 5, 9 and 13 days after mice were challenged with UTI and gavaged with probiotic products. At predetermined time intervals, aliquots were removed, serially diluted and plated on EMB agar (37°C, 18 h) to determine the bacterial colony counts.
Statistical Analysis
All data are expressed as the mean ± SD. ANOVA was used to evaluate differences between multiple groups. Significant differences were subjected to Duncan’s test to compare the means of two specific groups. A p value
Effect of the spent culture supernatants of LAB strains and probiotic products on the growth of UPEC
A total of 366 LAB strains obtained from fermented vegetables were studied. The experimental data on the effect of several treatments on the antimicrobial activity of LAB strains and probiotic products are presented in Table 1. When UPEC strains BCRC 15585, 10675 and 15479 were used as indicator bacteria, seven LAB strains (PM, PM68, PM78, PM20, PM206, PM229 and RY2) were able to better inhibit the growth of these bacteria than other LAB strains. The diameters of the inhibition zones for five LAB strains were all >23 mm, indicating that these LAB strains strongly inhibited the growth of three pathogen UPEC strains. These seven strains in the Table 1 were identified through API 50CHL tests as Lactobacillus paracasei (strain PM2), Lactobacillus salivarius (strain PM78), Lactobacillus plantarum (strains PM206 and PM229), Lactobacillus crispatus (strain RY2) and Pediococcus pentosaceus (strains PM68 and PM201).
| Inhibition zonea (mm) for UPEC strains | |||
|---|---|---|---|
| BCRC15585 | BCRC10675 | BCRC15479 | |
| LAB strains | |||
| PM2 | 29 (+++) | 21 (++) | 26 (+++) |
| PM68 | 23 (+++) | 23 (+++) | 23 (+++) |
| PM78 | 24 (+++) | 25 (+++) | 23 (+++) |
| PM201 | 21 (++) | 21 (++) | 29 (+++) |
| PM206 | 24 (+++) | 24 (+++) | 33 (+++) |
| PM229 | 23 (+++) | 24 (+++) | 33 (+++) |
| RY2 | 27 (+++) | 24 (+++) | 24 (+++) |
| Probiotic products | |||
| PP366 | 10 (-) | 10 (-) | 18 (++) |
| PP365 | 16 (+) | 18 (++) | 23 (+++) |
| LP142 | 18 (++) | 17.5 (++) | 27 (+++) |
| Yi Sheng Mi | 17.5 (++) | 20 (++) | 28 (+++) |
| ShuMi Chang | 16 (+) | 17 (++) | 30 (+++) |
| Yi Sheng Mi plus | 20 (++) | 21 (++) | 32.5 (+++) |
Table 1: Antimicrobial effects of the spent culture supernatant (SCS) broth from lactic acid bacteria and their probiotic products against three uropathogenic Escherichia coli strains. a. Interpretation of zone diameter of inhibition.-,
Similarly, the probiotic products (PP365, LP1, Yi Sheng Mi, Shu Mi Chang and Yi Sheng Mi plus) strongly inhibited the UPEC strain BCRC 15479; however, they only mildly inhibited the growth of UPEC strains BCRC 15585 and 10675 (inhibition zones were between 11 and 22 mm in diameter) (Table 1). These results revealed that the screened LAB strains and the probiotic products they fermented possess certainly antimicrobial activity against the growth of UPEC strains.
Effect of LAB strains and their probiotic products on the inhibition of UPEC adhesion to urothelial SV-HUC-1 cells
The adhesiveness of each probiotic strain was quantified via light microscopy using crystal-violet stain. Twenty microscopic fields were randomly counted to measure the number of bacteria attached to the SV-HUC-1 cells. The average number of adhered bacteria per 10 epithelial cells was calculated from these data and defined as the adhesion index (a measure of adhesiveness of a strain). The adhesive indices of these seven LAB strains are shown in Figure 1A. Most of the LAB strains adhered to SV-HUC-1 cell lines, and strain PM78 (43.4 ± 2.9 bacteria/cell) was found to adhere particularly strongly to human SV-HUC-1 cells.
Furthermore, SV-HUC-1 cells were co-incubated with the LAB strains and UPEC strains to evaluate whether the LAB strains could inhibit UPEC strain adhesion to SV-HUC-1 cells. The results showed that the adhesion of the UPEC strain to SV-HUC-1 cells was significantly (p<0.05) inhibited by treatments with any of the LAB strains or probiotic products (Figure 1B). These data indicate that, in addition to the factors in the SCS, the cell bodies of probiotics also play roles in inhibition of UPEC invasion.
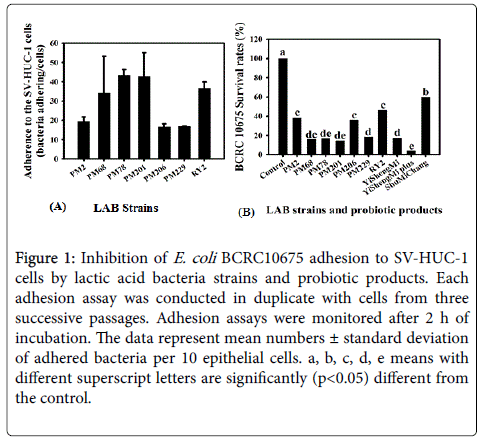
Figure 1: Inhibition of E. coli BCRC10675 adhesion to SV-HUC-1 cells by lactic acid bacteria strains and probiotic products. Each adhesion assay was conducted in duplicate with cells from three successive passages. Adhesion assays were monitored after 2 h of incubation. The data represent mean numbers ± standard deviation of adhered bacteria per 10 epithelial cells. a, b, c, d, e means with different superscript letters are significantly (p<0.05) different from the control.
Evaluation of antibacterial activity of the LAB strains and probiotic products on UPEC strains in human urine
The viabilities of the three UPEC strains were strong reduced by treatments with all seven of the LAB strains (data not shown). We next evaluated the viabilities of LAB strains, probiotic products and UPEC strains in human urine to simulate the conditions of human infection. The seven LAB strains showed potent inhibition of the growth of UPEC strains in a time-dependent manner (Table 2). Additionally, the survival rates of these three UPEC strains were strongly diminished by co-culture with the probiotic products, in particular by treatment with Shu Mi Chang (Table 2). The results suggest that secretion of antibacterial compounds and the synergistic effect provided by different lactic acid bacteria might contribute the powerful antimicrobial activity of probiotics.
| Survival rates of UPEC strains (%)a | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BCRC 10675 | BCRC 15479 | BCRC 15585 | ||||||||||
| Time (h) | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 |
| LAB strains | ||||||||||||
| PM2 | 85.2 | 16.15 | 0.87 | 0.05 | 31.45 | 22.34 | 0.22 | 0.01 | 10.59 | 0.55 | 0.24 | 0.01 |
| PM68 | 89.6 | 20.52 | 0.8 | 0.03 | 8.55 | 0.19 | 0.00 | 0.00 | 2.65 | 0.23 | 0.00 | 0.00 |
| PM78 | 56 | 4.9 | 0.06 | 0.00 | 10.32 | 0.29 | 0.00 | 0.00 | 3.88 | 0.39 | 0.02 | 0.00 |
| PM201 | 13.2 | 0.55 | 0.02 | 0.00 | 17.47 | 11.66 | 0.32 | 0.01 | 2.88 | 0.18 | 0.01 | 0.00 |
| PM206 | 9.12 | 0.07 | 0.00 | 0.00 | 1.69 | 0.03 | 0.00 | 0.00 | 0.98 | 0.04 | 0.00 | 0.00 |
| PM229 | 11.56 | 1.17 | 0.02 | 0.00 | 0.82 | 0.04 | 0.00 | 0.00 | 2.27 | 0.1 | 0.02 | 0.00 |
| RY2 | 37.4 | 6.35 | 0.94 | 0.06 | 12.74 | 1.53 | 0.16 | 0.00 | 11.96 | 2.39 | 0.57 | 0.03 |
| Probiotic products | ||||||||||||
| Yi Sheng Mi | 78.69 | 4.96 | 0.01 | 0.00 | 89.13 | 0.06 | 0.00 | 0.00 | 1.04 | 0.76 | 0.00 | 0.00 |
| Yi Sheng Mi plus | 8.46 | 0.02 | 0.00 | 0.00 | 0.09 | 0.00 | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.00 |
| ShuMi Chang | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
Table 2: Survival of UPEC co-cultured with lactic acid bacteria strains or probiotic products in urine. Each value represents the mean value from three independent experiments.
Lactic acid bacteria and probiotic products suppressed inflammatory injury in UPEC-induced SV-HUC-1 cells
Interleukin-6 and IL-8 are known to be strong chemokines that enhance neutrophil microbicidal activities and are used as markers of UTI caused by UPEC [23]. We found that E. coli BCRC10675 significantly (p<0.05) increased the level of IL-6 compared to the control group. Furthermore, the increased level of IL-6 in SV-HUC-1 cells was significantly (p<0.05) attenuated by the treatment with any of the LAB strains and probiotic products (Figure 2A). However, the levels of IL-8 were not markedly changed by treatment with E. coli BCRC10675 or LAB strains, whereas Yi Sheng Mi plus significantly (p<0.05) inhibited the level of IL-8 in UPEC-induced SV-HUC-1 cells. We also observed that the strain PM229 significantly (p<0.05) enhanced IL-8 levels in SV-HUC-1 cells both in the present or absence of E. coli BCRC10675 (Figure 2B).
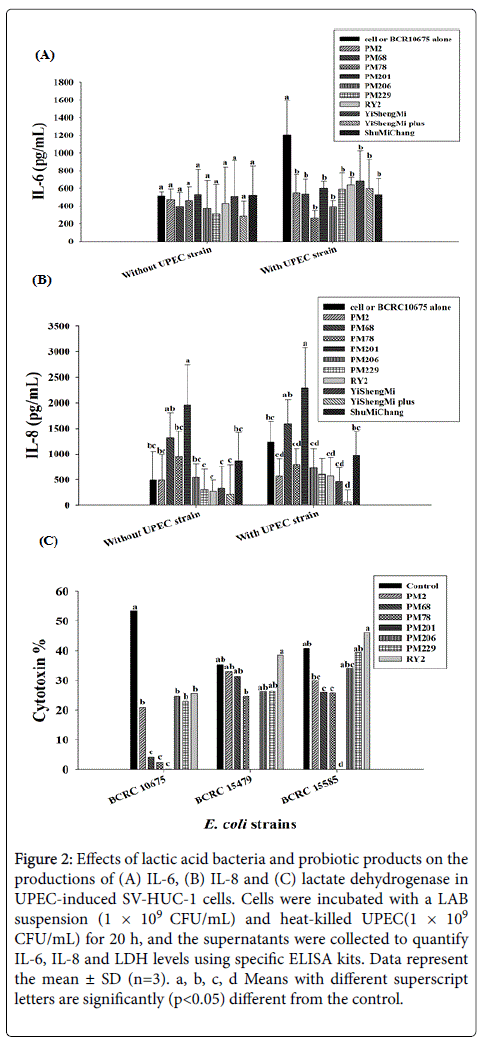
Figure 2: Effects of lactic acid bacteria and probiotic products on the productions of (A) IL-6, (B) IL-8 and (C) lactate dehydrogenase in UPEC-induced SV-HUC-1 cells. Cells were incubated with a LAB suspension (1 × 109 CFU/mL) and heat-killed UPEC(1 × 109 CFU/mL) for 20 h, and the supernatants were collected to quantify IL-6, IL-8 and LDH levels using specific ELISA kits. Data represent the mean ± SD (n=3). a, b, c, d Means with different superscript letters are significantly (p< 0.05) different from the control.
Lactate dehydrogenase is a soluble cytoplasmic enzyme present in almost all cells and is released into the extracellular space when the plasma membrane is damaged [24]. We therefore evaluated the status of LDH in SV-HUC-1 cells treated with three UPEC pathogens co cultured with LAB strains. Figure 2C shows that UPEC strains significantly (p<0.05) increased LDH release, indicating an enhanced level of cytotoxicity in SV-HUC-1 cells. The level of LDH was significantly (p<0.05) decreased by co-culture with the LAB strains, in particular with strain PM201. These results indicate that the LAB strains and their probiotic products protect against the injury of urothelial SV-HUC-1 cells through their antimicrobial activity as well as their immunomodulatory and anti-inflammatory capabilities.
Probiotic products prevented urethral damage in UPEC challenged BALB/c mice
We then studied the in vivo effect of the probiotic products on UPEC-challenged UTIs. As shown in Figure 3, after infection with E. coli BCRC10675 the viable counts of E. coli increased to 104 CFU/mL in the urine of BALB/c mice. Administration of probiotic products at a dose of 1 × 1011 CFU/mL for 14 consecutive days slightly reduced E. coli growth, whereas Yi Sheng Mi plus treatment significantly (p<0.05) inhibited the viable counts of E. coli in the urine of BALB/c mice during the treatment period. These results indicate that probiotic supplementation can inhibit pathogen growth and thus prevent urethral infection caused by UPEC strains in BALB/c mice.
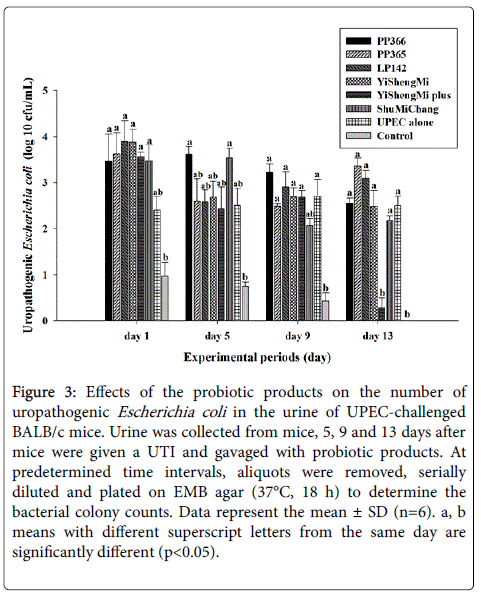
Figure 3: Effects of the probiotic products on the number of uropathogenic Escherichia coli in the urine of UPEC-challenged BALB/c mice. Urine was collected from mice, 5, 9 and 13 days after mice were given a UTI and gavaged with probiotic products. At predetermined time intervals, aliquots were removed, serially diluted and plated on EMB agar (37°C, 18 h) to determine the bacterial colony counts. Data represent the mean ± SD (n=6). a, b means with different superscript letters from the same day are significantly different (p<0.05).
Uropathogenic Escherichia coli are a group of pathogens that cause a wide spectrum of diseases that affect all ages worldwide. The increasing incidence of disease caused by UPEC, the associated costs, and the burgeoning problems associated with the emergence and spread of multidrug-resistant UPEC strains indicate that an effective strategy against UPEC infection is urgently needed [25,26]. We performed antimicrobial tests against UPEC strains and adherence assays to the human urothelial SV-HUC-1 cell line to select potential LAB strains from fermented vegetables. The results showed that LAB cells could inhibit the adherence of pathogenic bacteria to host cells. Moreover, the LAB strains and probiotics products might release lactic acid, short chain peptides, or protease-resistant bactericidal substances, even when grown in human urine and thus inhibit UPEC growth. The modulation of antimicrobial immunity (IL-6 and IL-8) by LAB was also observed and thus protected host cells against the pathogen-induced inflammatory response. Mathoera et al. [27] suggested that adherent LAB strains are able to protect urothelial cells from infection by pathogenic bacteria. Reid and Bruce [28] and Falagas et al. [29] also indicated that such inhibitory effects of probiotics were mediated via a mechanism of non-specific steric hindrance on the receptors for pathogen binding. The adhesion of LAB to host urinary tract epithelium may result in the competitive exclusion of adhesion of pathogenic bacteria [11]. Moreover, lactobacilli are the dominant microbes in the vaginal flora of many healthy women, which produce lactic acid and other substances that keep the vaginal pH low and prevent the overgrowth of pathogens [30]. Delley et al. [2] also reported that specific lactobacilli strains have the ability to interfere with the adherence, growth and colonization of human bladder cells by uropathogenic bacteria. The results presented here show that the LAB strains we selected have the potential for probiotic use. Furthermore, multi-strain probiotics appear to show greater efficacy than single strains, which might be due to the synergistic interactions between strains, though it may also be a consequence of the higher probiotic dose used in this study.
In vitro experimentation is useful for clarifying the ability of probiotics to inhibit the growth of uropathogens. There is sufficient evidence from in vitro studies to elucidate the mechanism of action of probiotics in preventing UTIs. We also employed mice to determine the effect of probiotics in an in vivo model of UPEC-challenged UTIs. The results indicated that the probiotic product Yi Sheng Mi plus repressed E. coli growth and thus prevented bladder tissues damage caused by UPEC pathogens (data not shown). The inhibitory mechanism of probiotic products on UPEC pathogen infection is at least partly mediated through vigorous antimicrobial activity. de Arellano et al. [31] reported that orally administration of Lactobacillus plantarum LPLM-O1 for seven days reduced infection, decreased the concentration of urinary leukocytes, and lowered the bacterial load in an E. coli-induced animal model of urethral infection. Asahara et al. [22] also suggested that intraurethrally administered probiotic Lactobacillus casei strain Shirota inhibited the growth of E. coli and inflammatory responses in a murine UTI model. Moreover, the production of bacteriocin and organic acids by strain Lactobacillus fermentum L23 suppressed the pathogenic microorganism within the vaginal environment in mice [32]. In the present study, our observations also provide powerful evidence for using probiotics to prevent UTIs caused by uropathogenic bacteria.
In conclusion, the results presented here show that the LAB we selected are potential strains for probiotic use. These probiotics show antagonistic activity against UPEC infection in vitro and in vivo and suggest that these probiotics, which were isolated from fermented vegetables, may be effectively utilized in the prevention of urinary tract infections.
This study was funded by the 103B-29-011 project from Ministry of Education, Taiwan.