Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2016) Volume 4, Issue 1
Introduction: Probiotics are defined as live microorganisms, which when administered in adequate amounts, confer a health benefit on the host. Health benefits have mainly been demonstrated for specific probiotic strains of the following genera: Lactobacillus, Bifidobacterium, Saccharomyces, Enterococcus, Streptococcus, Pediococcus, Leuconostoc and Bacillus. The human microbiota is getting a lot of attention today and research has already demonstrated that alteration of this microbiota may have far-reaching consequences. One of the possible routes for correcting dysbiosis is by consuming probiotics.
Methods: In this research we investigated the influence of potentially pathogenic challenge bacteria Pseudomonas aeruginosa on multispecies probiotic food supplement and kefir microbiota. The probiotics included various strains of the species Lactobacillus acidophilus, Lactobacillus reuteri, Lactobacillus rhamnosus, Lactobacillus delbrueckii subs, bulgaricus, Lactobacillus plantarum, Lactobacillus lactis subs, lactis, Bifidobacterium bifidum, Bifidobacterium breve, Bifidobacterium longum, Streptococcus thermophilus, Enterococcus faecium. Samples of 40 mL of milk were prepared with added challenge potentially pathogenic Pseudomonas aeruginosa and 1 mL suspension of multispecies probiotics. All samples were incubated for 4 days.
Results and discussion: It was found that both the multispecies probiotic supplement and the kefir microbiota with diverse microbial populations successfully decreased the concentration of challenge potentially pathogenic Pseudomonas aeruginosa for 3 log10 steps. On the other hand the antagonistic effect of the oligospecies probiotics with only three different probiotic species and the monospecies probiotic supplement against P. aeruginosa was not detected. These results show that multispecies microorganisms that create a live communities create a synergistic effect and effective complex interconnecting quorum-sensing regulatory networks that compete with a potentially pathogen bacteria.
Conclusions: Although probiotic administration does not permanently modulate the intestinal microbiota, this does not mean that during acute disruption of the sensitive intestinal microbiota balance such as due to antibiotic consumption, transiently present probiotics do not aid the permanent intestinal microbiota in restoring this balance.
Keywords: Probiotics; Inflammatory action; Fermentation
As a consequence of its large genome Pseudomonas aeruginosa is a common, versatile and adaptable opportunistic pathogen in hospitalized, immune compromised patients [1,2]. The ability of P. aeruginosa to colonize and infect a variety of sites has been attributed to the many virulence factors that it can produce. Infection caused by P. aeruginosa is often life-threatening and difficult to treat because of its primary limited susceptibility to commonly used antimicrobial agents. Therefore utilization of alternative antibacterial therapies such as combining antibiotics and probiotics could prove to be a synergistic combination. It has also been published that probiotics can also weaken the resistance of multiple drug resistance in P. aeruginosa [3,4].
By definition probiotic foods must contain live microorganisms that, when administered in adequate amounts confer a health benefit to the host [5]. The most common probiotic bacteria are certain strains from the genera Lactobacillus (i.e., L. rhamnosus , L. acidophilus, L. plantarum , L. cassei , L. delbrueckii subsp. bulgaricus etc.) and Bifidobacterium (i.e., B. infantis , B. animalis subsp. lactis , B. longum etc.). Other probiotic bacteria include: Pediococcus acidilactici , Lactococcus lactis subsp. lactis , Leuconostoc mesenteroides , Bacillus subtilis , Enterococcus faecium , Streptococcus thermophilus , Escherichia coli etc. Certain yeasts such as Saccharomyces boulardii are also probiotics [6]. Probiotics together with other beneficial microbes are commensals of the gut and differ from pathogenic bacteria in the terms of their actions on immune cells in the gut as they do not stimulate the proliferation of mononuclear cells or trigger an inflammatory action [7].
Probiotics are resistant to gastric and bile acids, enabling their survival through the gastrointestinal tract. When probiotics reach the small intestine; they multiply and exhibit their positive health effect. Probiotics commonly do not permanently inhabit the intestines, but exhibit a transient effect [5]. The intestinal microbiota is very well adapted, exceptionally stable and very specific for each individual. In normal conditions of stable functioning of the digestive system neutral and beneficial microorganisms dominate. It is estimated that there are 100 trillion microorganisms in the intestine of a human adult and this is 10 times larger than the number of cells in the human body [8,9]. The balance of the intestinal microbiota is negatively influenced by modern lifestyle, leading to increased numbers of pathogenic microorganisms that disrupt microbial balance and cause a reverse from beneficial to harmful functioning. In such cases the external support with probiotics is very welcome and supported by several scientific studies [10]. Recent research has shown that probiotic microorganisms play an important part in enhancing our immune system, competitive inhibition and bacteriocins production against pathogenic microorganisms etc. [9]. The most commonly used sources of probiotics include: probiotic supplements, probiotic yogurts, kefir, sauerkraut and other fermented vegetables [11].
Multispecies probiotics have certain advantages over monostrain preparations. Mixed preparations may complement each other’s effect through synergism and/or symbiosis [6]. Active microbiota in kefir grain ecosystems also exhibit such effects [12]. Kefir grains are a unique symbiotic community of different microbial genera and species, mainly lactic acid bacteria (LAB), yeasts and occasionally acetic acid bacteria (AAB), cohabiting in a natural polysaccharide (called kefiran) and a protein matrix. The composition somewhat varies due to geographical origin, but the predominant bacteria include: Lactobacillus kefiranofaciens , Lactobacillus buchneri , Lactobacillus helveticus , Lactobacillus acidophilus and other Lactobacillus species, Streptococcus thermophilus, Acetobacter genera, Enterococcus sp., Oenococcus sp., Pediococcus sp., Leuconostoc sp. Various yeasts such as Saccharomyces cerevisiae, Dekkera anomala (Brettanomyces anomalus, anamorph) and even filamentous moulds are also present. The complexity of their physical and microbial structures is the reason that the kefir grains are still not unequivocally elucidated. Another important factor in the microbiota composition is the sub-culturing method: viable counts on selective culture media, PCR-DGGE, pyrosequencing give different compositions. Microbiota of kefir grains has been studied by many microbiological and molecular approaches. The development of metagenomics, based on the identification without cultivation, is opening new possibilities for identification of previously non-isolated and non-identified microbial species from the kefir grains [12-14].
In this research the aim was to determine if selected multispecies probiotic preparations and the active microbiota in the kefir grain ecosystem inhibit growth of the potentially pathogenic bacteria Pseudomonas aeruginosa during fermentation in milk and if one microbiota is more efficient than the other.
Reference strain of potential pathogen and growth conditions: An overnight culture of Pseudomonas aeruginosa, ATCC 27853 subcultured in tryptic soy broth for 24 hours at 37°C was used. Selective medium cetrimide agar base (Fluka 22470) with added glycerol was used for incubation and enumeration.
Probiotic supplements: The probiotic supplements are noted in Table 1. Manufacturers’ names are omitted in order to enable impartiality of research. Total colony count was determined by anaerobic incubation on tryptic soy agar (TSA) (Sigma Aldrich 220191) at 37°C for 3 days. Kefir grains were obtained from a private household in Slovenia and originated from the Caucasian mountains. Kefir grains were previously activated by incubation in 1 L bovine milk at room temperature (24°C).
| Sample | Total anaerobic/ facultative anaerobic microorganisms (cfu*/mL) |
Microorganisms | Other additives |
|---|---|---|---|
| Multispecies probiotic supplement | 4.0× 108 | Lactic acid bacteria in lyophilised form: Lactobacillus acidophilus (NIZO 3678); Lactobacillus acidophilus (NIZO 3887), Lactobacillus paracasei (NIZO 3672); Lactobacillus plantarum (NIZO 3684); Lactobacillus rhamnosus (NIZO 3689); Lactobacillus salivarius (NIZO 3675), Enterococcus faecium (NIZO 3886) Bifidobacteria in lyophilised form: Bifidobacterium bifidum (NIZO 3804); Bifidobacterium lactis (NIZO 3680) |
Corn, starch, inulin,amylase,potassium, manganese, magnesium, vanilla aroma |
| Oligospecies probiotic supplement | 1.5×108 | Lactobacillus acidophilus (species L. gasseri), Enterococcus faecium Bifidobacterium infantis |
magnesium stearate, lactose, dextrin, potato starch |
| Monospecies probiotic supplement | 5.9×108 | Lactobacillus reuteri DSM 17938 | sunflower oil, caprylic acid triglycerides, silica dioxide |
| Kefir microbiota | 3.0×109 | Various lactic acid bacteria: Lactobacillus kefiranofaciens, Lactobacillus buchneri, Lactobacillus helveticus, Lactobacillus acidophilus and others Streptococcus thermophilus Enterococcus sp., Oenococcus sp., Pediococcussp., Leuconostoc sp. Various acetic acid bacteria: Acetobacter genera Yeasts: Saccharomyces cerevisiae Dekkeraanomala and others |
kefiran |
Table 1: Characteristic of the probiotic supplements. *cfu: Colony forming units.
Fermentation: 40 mL of previously sterilised low-fat bovine milk was inoculated with the potential pathogen P. aeruginosa or with a mixture of the pathogen in the presence of the multispecies probiotic food supplements or the mixture of kefir microbiota. Fermentations were conducted with agitation (250 min-1) at 25°C for 4 days. Two separate experiments were conducted and the average was calculated for each sample.
Enumeration of culturable bacteria: Viable cell counting was determined by making 10-fold serial dilutions. Cetrimide agar with added glycerol was used to enumerate cfu of P. aeruginosa. TSA agar was used to enumerate the total cfu of anaerobic or facultative anaerobic microorganisms.
The enumeration of the potentially pathogenic bacteria Pseudomonas aeruginosa after the four day fermentation in milk at room temperature with or without added beneficial microorganisms is presented in Figure 1.
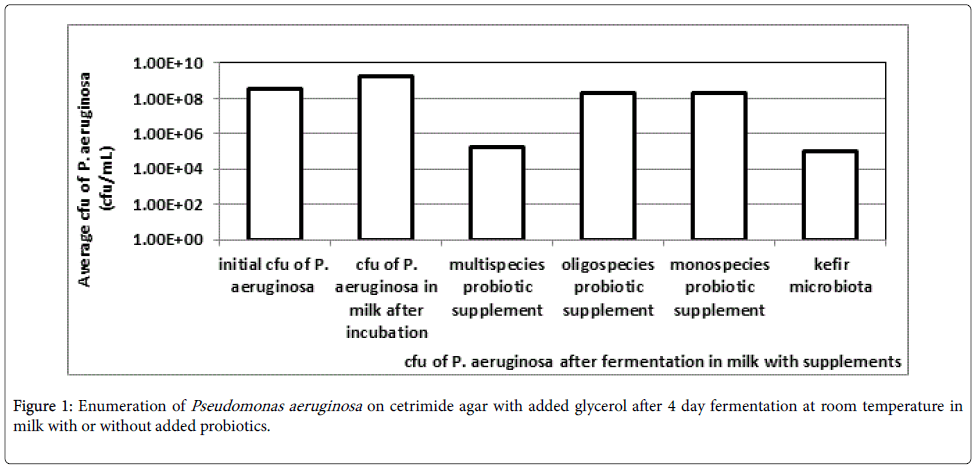
Figure 1: Enumeration of Pseudomonas aeruginosa on cetrimide agar with added glycerol after 4 day fermentation at room temperature in milk with or without added probiotics.
The results in Figure 1 show a 10-fold increase in the concentration of the potential pathogen after the four day fermentation in milk. All chosen probiotic supplements caused a decrease in the concentration of Pseudomonas aeruginosa after the four day fermentation in milk. However the decrease was less than 10-fold for the oligospecies probiotic and the monospecies probiotic. On the other hand a 3 log step reduction was found for both the multispecies probiotic and the kefir microbiota. Preliminary experiments conducted without agitation (data not shown) yielded much lower counts of P. aeruginosa fermented in milk or in milk with added probiotics or kefir microbiota. This is perhaps due to the fact that P. aeruginosa is an obligate aerobe bacterium that thus cannot survive without oxygen. With our experiments agitation enabled better aeration of the samples.
The total cfu of microorganisms incubated in anaerobic conditions after four day fermentation in milk with probiotics and with or without P. aeruginosa are shown in Figure 2. This test was conducted to determine the total number of microorganisms under anaerobic conditions to see if the four day fermentation itself with or without P. aeruginosa influenced the number of probiotic bacteria. Although P. aeruginosa itself does not grow under anaerobic conditions as it is an obligate aerobe, its presence influences the total number of the anaerobic bacteria.
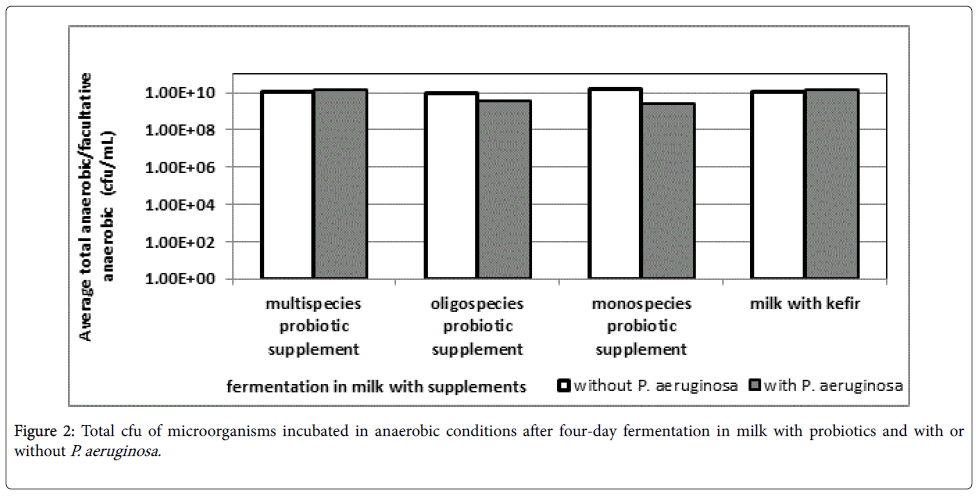
Figure 2: Total cfu of microorganisms incubated in anaerobic conditions after four-day fermentation in milk with probiotics and with or without P. aeruginosa.
The total cfu of microorganisms of all samples incubated in anaerobic conditions with probiotics and without P. aeruginosa reached around 1010 cfu/mL. However, adding P. aeruginosa to oligospecies probiotic and the monospecies probiotic resulted in a 10-fold decrease of total cfu incubated in anaerobic conditions. These samples also exhibited a lower decrease of the potential pathogen noted in Figure 1. On the other hand, the total cfu of microorganisms of multispecies probiotic and the kefir microbiota incubated in anaerobic conditions exhibited slight increase. These two samples also yielded a higher decrease of the potential pathogen noted in Figure 1.
Effective probiotics should exhibit tolerance and adaptation to stress conditions typical of the gastrointestinal tract. Adhesion to human intestinal epithelial cells and competitive exclusion of pathogens are also considered important. Such effects are much more efficient in a collective population. For many years, bacterial cells were considered primarily as selfish individuals, but in recent years it has become evident that far from operating in isolation, they coordinate collective behaviour in response to environmental challenges using sophisticated intercellular communication networks [15]. The antagonistic activity of one microorganism against another can be caused by competitive exclusion, immune modulation, stimulation of host defence systems, production of organic acids or hydrogen peroxide that lower pH, production of antimicrobials such as bacteriocins, antioxidants, production of signalling molecules that trigger changes in gene expression etc. [16-18]. Another important fact that cannot be overlooked is that bacteria are not limited to communication within their own species but are capable of intercepting messages and coerce cohabitants into behavioural modifications [15].
Pseudomonas aeruginosa is one of the important bacteria that can cause huge burdens for public health today due to its ability to adapt its genome and physiology during chronic infections. Major features of P. aeruginosa, making it a very successful opportunistic pathogen of chronic diseases include: reduction of virulence, biofilm formation, motility and quorum sensing [19]. Most of these features have been extensively researched. Emerging knowledge of interspecies polymicrobial interactions is showing that P. aeruginosa is capable of converting isolates; which are alone avirulent or even beneficial; to synergistically exacerbate the pathogenicity of P. aeruginosa in a polymicrobial community or that P. aeruginosa employs quorum sensing molecules that exhibit biological functions beyond signalling including: antibacterial and quorum-quenching activity towards Gram-positive bacteria [15,20,21]. Although recent research has found that the autoinducer-2 (AI-2) signal is present in a variety of bacteria, information regarding quorum-sensing of lactobacilli is very sparse. The AI-2 signal system plays an important role in the response of probiotic lactobacilli to the surrounding environment. A recent research has found that by monitoring the AI-2 activity of two strains each of Lactobacillus rhamnosus and L. plantarum under various stress conditions, a correlation was found between the AI-2 inhibition and reduction in stress, thus suggesting that quorum signalling of probiotic lactobacilli may be a way of adapting to the host’s ecosystem and interaction with the intestinal environment [22]. Likewise, a recent publication has shown for the first time that bifidobacteria also exhibit biofilm production and quorum sensing as they produce autoinducer-2 (AI-2) molecules [23].
In this research it was found that the decrease of the potential pathogen P. aeruginosa was dependant on the type of probiotic microbiota and that both multispecies microbiota (multispecies probiotic supplement kefir microbiota) with a much more diverse population than the other two samples (oligospecies probiotic supplement and the monospecies supplement) exhibited an efficient synergistic effect [6]. Both samples also exhibited a higher increase in the total population of anaerobic microorganisms after fermentation with P. aeruginosa thus indicating that a more successful quorum-sensing regulatory network was established and yielded antagonistic effects against the potential pathogen P. aeruginosa. The results also show that the potential pathogen P. aeruginosa also created a successful quorum-sensing regulatory network that survived the antagonistic efforts of all probiotic bacteria as no combination of probiotics completely inhibited this potential pathogen. These results indicate that the antagonist properties of probiotics are not universal to all other microorganisms.
Ratsep et al. [18] found that various strains of Lactobacillus plantarum exhibited antagonistic activity against the common pathogen Clostridium difficile in vitro. The antagonistic activity was strain specific and in the case of using L. plantarum culture supernatant dependant on the temperature and pH. Lazarenko et al. [24] found that several strains of lactobacilli and bifidobacteria had an efficient antagonistic effect against Staphylococcus aureus in vitro (as well as and on models of experimental intravaginal staphylococcosis of mice). Aminnezhad et al. [1] found that cell-free supernatant Lactobacillus strains synergistically interacted with amikacin and gentamicin to inhibit the growth of P. aeruginosa. These experiments are important in vitro experiments that mainly determine the inhibition zones of pathogens on culture media with various probiotics; however they do not simulate real conditions.
In this research the method of determining the antagonistic effect of various probiotics on the growth of the potential pathogen P. aeruginosa included a simulation where the probiotic microorganisms actively function, communicate and create quorum-sensing regulatory networks. Also the effectiveness of various bacteriocins of probiotics may be hindered by the proteolytic activity of microbial enzymes that are secreted only during active fermentation [25]. In this research the potential pathogen P. aeruginosa also created a successful network as no probiotic combination completely inhibited this pathogen, but only reduced its growth. Our results along with other research previously mentioned implicates that P. aeruginosa is successful in surviving many different environments and under various stress conditions.
The human microbiome is very diverse and unique to every individual. The gut microbiota expresses over 3.3 million bacterial genes, while the human genome expresses only 20 thousand genes [26], more and more studies are confirming that manipulation of non-pathogenic strains in the host can stimulate the recovery of the immune response to pathogenic bacteria causing diseases. One important approach for controlling dysbiosis is the use of nutraceutics including probiotics with their entire quorum sensing systems or the development of alternative next-generation antibiotics based on pro- and anti-quorum sensing therapies (disruption of quorum sensing) that function by modulating bacterial chemical communication circuits [27]. However most of this research is at its beginning. Thus further research on the exact mechanisms of interspecies communications between chosen probiotic microorganisms and potential pathogens is necessary.
The financial support of the Slovene Human Resources development and Scholarship Fund grant number OP RCV 2007-2013 with the acronym PROBIORES was greatly acknowledged. The author is also grateful for the conscientious efforts of all the students (Natalija Grah, Jure Žganec, Dunja Šulc, Tadeja Godec, Kristina Martinovic and Tina Režek) during the preliminary experimental work in the frame of the project PROBIORES. These preliminary experiments were the starting point for all the work subsequently conducted by the author for this article.