Journal of Hematology & Thromboembolic Diseases
Open Access
ISSN: 2329-8790
ISSN: 2329-8790
Research Article - (2013) Volume 1, Issue 3
Keywords: CDH13; Methylation; AML; Genetics; Pediatrics
Acute leukemia is the most common malignancy diagnosed in children, representing nearly one third of all pediatric cancers. Pediatric Acute Myeloid Leukemia (AML) comprises up to 20% of whole childhood leukemia. Pediatric AML is a heterogeneous clonal disorder of hematopoietic progenitor cells, is a complex and lifethreatening disease which lose the ability to differentiate normally [1]. Recently, epigenetic and methylation disorders, such as aberrant promoter hypermethylation and abnormal histone modifications have been implicated in the pathogenesis of leukemia [2,3]. These include aberrations in methylation, which is a key epigenetic event responsible for enhanced proliferation and self-renewal, differentiation arrest, and impaired apoptosis of leukemic cells [4]. Inactivation of tumor suppressor genes by promoter hypermethylation has been recognized as key event in the development of pediatric AML. Compared to the incidences of DNA mutations and deletions, the frequency of aberrant DNA methylation of tumor suppressor genes is high in AML. This suggests that aberrant DNA methylation has important roles in this rare cancer. Identifying these methylated genes and deeply study of these genes may provide better understanding of many tumors , including pediatric AML [5].
CDH13 (also known as H-cadherin and T-cadherin) is a member of the cadherin gene super family which was isolated and mapped to 16q24. CDH13 hypermethylation has been documented in breast [6-8] , lung cancers [9-11], pituitary adenomas [12,13], diffuse large B cell lymphoma [14], nasopharyngeal carcinoma [15-18] and cutaneous squamous cell carcinomas [19,20]. CDH13 has been suggested as an early marker for lung cancers [21]. It is generally associated with poor prognosis of patients with lung cancers [22,23], ovarian cancers [24], basal cell cancers [20] and gallbladder carcinomas [25], cervical cancers [26-28] and prostate cancers [29-31].
There are several studies suggested that CDH13 may functions as a tumor suppressor gene and possesses potent antitumor activity in several human cancers both in vitro and in vivo. Over-expression of CDH13 in MDA-MB-435 (human breast cancer cells) can reduce their invasive and tumor formation potential in vitro and in vivo [32]. Loss of CDH13 is associated with tumorigenicity of human non-small cell lung cancers. Over expression of CDH13 in cutaneous squamous cell carcinoma cells can induce a delay in the G2/M cell cycle and inhibit the proliferation of cancer cells [20].
To date, there have been few reports in relation to the expression of CDH13 and the methylation status of its promoter in pediatric leukemia. In this study, we have provided the first evidence of CDH13 methylation in both AML cell lines and pediatric samples. These suggest that CDH13 may function as a tumor suppressor in pediatric AML.
Cell lines
Leukemia cell lines HL-60, MV4-11, SHI-1, U937, DAMI, K562 and SHI-1 were obtained from the American Type Culture Collection (ATCC). Jurkat and 697 cells lines (gifts from Professor Wang Jian- Rong, The Cyrus Tang Hematology center of Soochow University). The entire cell lines were maintained at 37°C in the RPMI 1640 supplemented with 10% fetal bovine serum.
Patients and samples
Bone marrow specimens were obtained at the time of diagnosis during routine clinical assessment of 70 patients with AML, who presented at the Department of Hematology and Oncology, Children’s Hospital of Soochow University between 2000 and 2010. Ethical approval was provided by the Children’s Hospital of Soochow University Ethics Committee (No. SUEC2000-021), and informed consent was obtained from the parents or guardians. AML diagnosis was made in accordance with the revised French–American–British (FAB) classification. Cytogenetic data were available in 64 patients. The main clinical and laboratory features of the patient cohort are summarized in table 1. Additionally, bone marrow samples from 12 healthy donors and 18 patients with Idiopathic Thrombocytopenic Purpura (ITP) were analyzed as controls. Bone marrow Mononuclear Cells (BMNCs) were isolated using Ficoll solution within 2 h after bone marrow samples harvested and immediately subjected for the extraction of total RNA and genomic DNA.
| Patient’s parameter | Status of CDH13 methylation | |||
|---|---|---|---|---|
| Methylated (n=24) | Unmethylated(n=46) | Total | p value | |
| Age (median and range, year) | 6.30(1-13) | 6.72(1-11) | 6.47(1-13) | 0.94 |
| Gender (male and female) | 14/10 | 20/26 | 34/46 | 0.88 |
| Laboratory parameters ( median and range ) | ||||
| WBC ( 109/L) | 16.72(0.8-51.1) | 16.31(0.8-43.6) | 16.43(0.8-51.1) | 0.90 |
| Hemoglobin (g/L) | 75.36(32-176) | 72.11(32-107) | 73.20(32-176) | 0.83 |
| Platelet count ( 109/L) | 68.37(12-310) | 64.12(23-273) | 65.54(12-310) | 0.75 |
| FAB subtype, n | ||||
| M1 | 2 | 10 | 12 | |
| M2 | 12 | 20 | 32 | |
| M3 | 6 | 4 | 10 | |
| M4 | 2 | 3 | 5 | |
| M5 | 5 | 4 | 9 | 0.22 |
| Cytogenetic, n | ||||
| Normal | 10 | 13 | 23 | |
| Abnormal | 24 | 21 | 45 | 0.61 |
| CDH13 transcript | 0.53 | 1.72 | 1.31 | 0.04* |
*p<0.05 FAB, French-American-British; WBC, white blood cells.
Table 1: Correlation of CDH13 methylation with clinical features in pediatric AML patients.
Semi-quantitative RT-PCR
RT-PCR was analyzed according to the manufacturer’s instructions. Primer sequences for CDH13 cDNA were designed according to Sun [18], generating a 203-bp PCR product: CDH13-RT-forward: TTCAGCAGAAAGTGTTCCATAT and CDH13-RTreverse: GTGCATGGACGAACAGAGT. PCR was carried out in a total volume of 20 μl system. Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) was amplified from the same cDNA sample as the internal control. The primer sequences for GAPDH cDNA were: GAPDH-RT-forward: AAGCTCACTGGCATGGCCTT, and GAPDH -RT-reverse:CTCTCTTCCTCTTGTGCTCTTG, generating a 375-bp PCR product. PCR conditions were 94°C for 30 s, 58°C for 30 s (CDH13) or 60°C for 30 s (GAPDH), and 72°C for 30 s, 33 cycles for the CDH13 gene and 24 cycles for the GAPDH gene. The amplified PCR products were then identified on 2% agarose gels. Images were acquired with a CCD camera (Bio-Rad, USA).
Quantitative Real-time PCR
RNA isolation and first-strand cDNA was synthesized as described above. Real-time PCR was performed according to the manufacturer’s protocol (Light Cycler 480 system, Roche). In brief, PCR mixture contained 100 pmol of each primer, Light Cycler 480 SYBR Green I Master (04 887 352 001 Roche, USA) and 2 μl cDNA. PCR conditions were 94°C for 10 s, 60°C for 10 s and 72°C for 15 s, 45 cycles for the CDH13 and GAPDH gene.
Sodium bisulphite modification of genomic DNA
The sodium bisulphite modification procedure was according to the manufacturer’s instructions of EZ DNA methylation Gold Kit (www.zymoresearch.com). Briefly 2 μg of extracted DNA was bisulphite-modified with the EZ DNA methylation Kit which converted all unmethylated cytosines to uracils and leaving methylcytosines unaltered. Modified DNA was resuspended in TE buffer (10 mM Tris/HCl, 1 mM EDTA, pH 7.5).
Methylation-specific PCR
The methylation status of the CDH13 promoter region was determined by methylation-specific PCR. Primers distinguishing unmethylated (U) and methylated (M) alleles were designed to amplify the sequence: CDH13-M-forward: 5-GTTTTTTTGGTGAGTTTTCGTTTTCGTTC-3; CDH13-M-reverse: 5-AATACCAAATCTCCCTATTCTCCGCG-3; CDH13-U-forward: 5-TTGTTTTTTTGGTGAGTTTTTGTTTTTGTTT-3; CDH13-U-reverse: 5-AAAATACCAAATCTCCCTATTCTCCACA-3.
Each PCR reaction contained 20 ng of sodium bisulphite-modified DNA, 250 pmol of each primer, 250 pmol deoxynucleoside triphosphate, 1× PCR buffer, and one unit of ExTaq HS polymerase (Takara, Tokyo) in a final reaction volume of 20 μl. Cycling conditions were initial denaturation at 95°C for 3 min, 40 cycles of 94°C for 30 s, 58°C (M) or 56°C (U) for 30 s, and 72°C for 30 s. PCR products were separated on 2% agarose gels.
Bisulfite genomic sequencing
Bisulfite Genomic Sequencing (BGS) were performed as previously described [33]. BGS primers were CDH13-F1:5- AAAGAAGTAAATGGGATGTTATTTT-3 and CDH13-R1:5- ACCAAAACCAATAACTTTACAAAAC-3.
CDH13-F2: 5- GTGATGTTGTTGTTGATTTATTTGG -3 and CDH13-R2: 5- AACCCCTCTTCCCTACCTAAAA-3. Amplified BGS products were TA-cloned and five to six randomly chosen colonies were sequenced. DNA sequences were analyzed with BiQ Analyzer (http://biq-analyzer.bioinf.mpi-inf.mpg.de). (Additional files 1 and 2).
Statistical analysis
SPSS v11.5 (SPSS Inc., Chicago, IL) was used for statistical analysis. Association between methylated sample data and clinical pathological features of AML patients were analyzed by Pearson chi-square test or Fisher’s exact test. p<0.05 was considered statistically significant.
CpG islands in the promoter of CDH13
Previously, we have analyzed the expression profiles of two AML cells before and after treatment with 5-Aza and found that the CDH13 may be related with promoter methylation in AML cells. We analyzed the sequence of CDH3 promoter and found there are three CpG island areas in the promoter of CDH13 (Figure 1A). The correlation between aberrant methylation and downregulation of CDH13 has been extensively documented in numerous cancers and cell lines, as lung caner, gastrointestinal system, reproductive system and numerous cell lines. There still rare repots about the methylation status of CDH13 in blood system, especially in the pediatric leukemia.
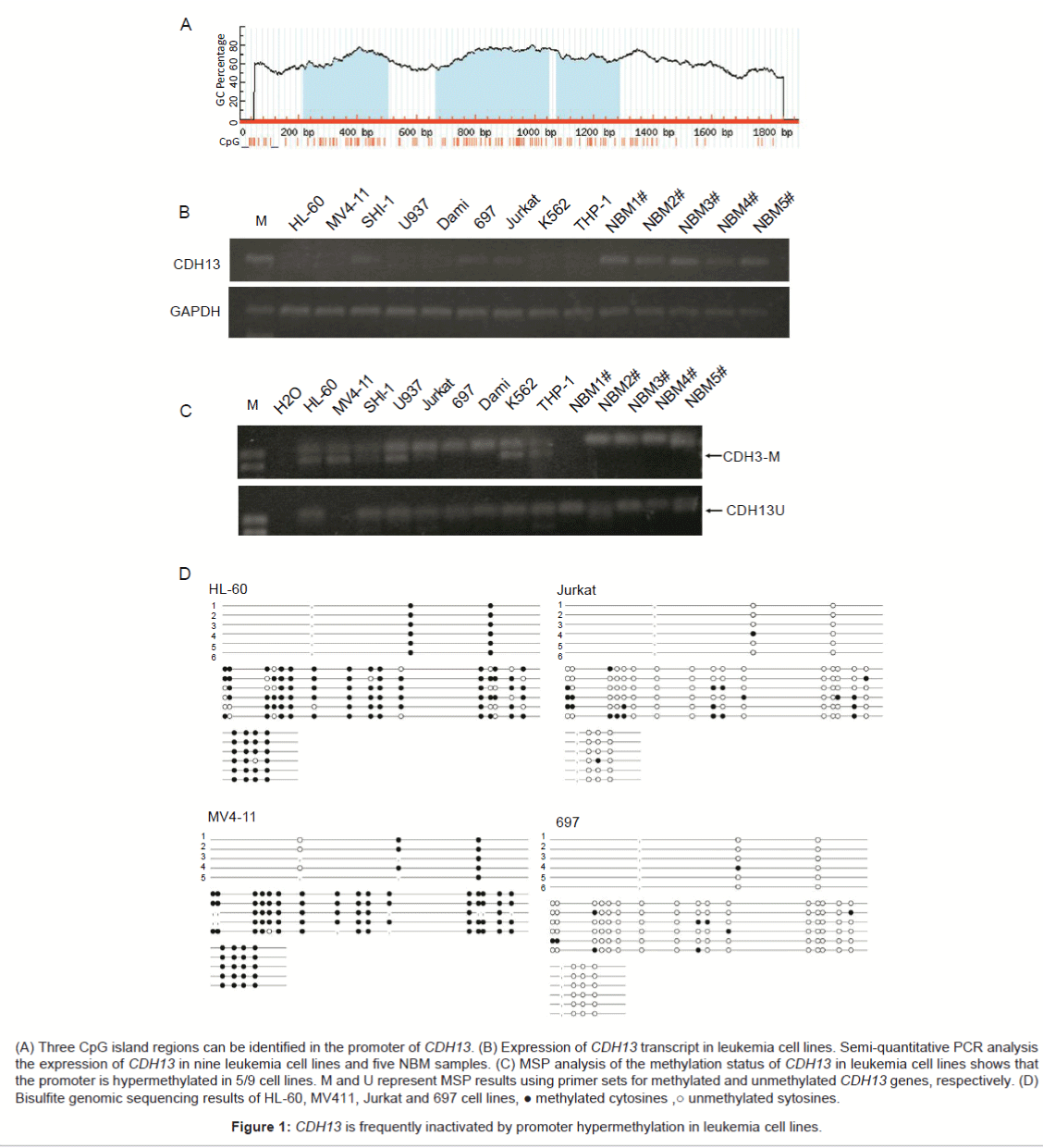
(A) Three CpG island regions can be identified in the promoter of CDH13. (B) Expression of CDH13 transcript in leukemia cell lines. Semi-quantitative PCR analysis the expression of CDH13 in nine leukemia cell lines and five NBM samples. (C) MSP analysis of the methylation status of CDH13 in leukemia cell lines shows that the promoter is hypermethylated in 5/9 cell lines. M and U represent MSP results using primer sets for methylated and unmethylated CDH13 genes, respectively. (D) Bisulfite genomic sequencing results of HL-60, MV411, Jurkat and 697 cell lines, ● methylated cytosines ,○ unmethylated sytosines.
Figure 1: CDH13 is frequently inactivated by promoter hypermethylation in leukemia cell lines.
Expression of CDH13 transcript in leukemia cell lines
Semi-quantitative PCR analysis showed CDH13 transcript is very low in nine leukemia cell lines. The expression of CDH13 in only three cell lines can be detected (3/9). Mean while, the expression of CDH13 in NBM group is significantly higher; the expression of CDH13 in all of 5 NBM samples can be detected with PCR (Figure 1B).
CDH13 promoter is hypermethylated in leukemia cell lines
Methylation-specific PCR (MSP) assays were performed to detect the methylation status of the CDH13 promoter in 9 leukemia cell lines. The MSP primer was designed using MethPrimer (http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi) to encompass the CpG islands of the CDH13 promoter identified in figure 1A. The CDH13 promoter was hypermethylated in 5 of 9 leukemia cell lines (HL-60, MV4-11, U937, K562 and THP-1). SHI-1, Jurkat, 697 and DAMI cells were unmethylated representative results of MSP were shown in figure 1C. Promoter methylation of HL-60, MV411, Jurkat and 697 cell lines was analyzed with bisulfite genomic sequencing. And the results consistent with the MSP assay (Figure 1D). In summary, these results showed that the CDH13 promoter was consistently significantly methylated in leukemia cells, such as HL-60, MV4-11, U937, K562 and THP-1. In contrast the CDH13 promoter was unmethylated in human lymphoblastic leukemia cells, such as Jurkat and 697. Based on these findings, we proposed that the promoter of CDH13 may be methylated in pediatric AML patients.
The promoter of CDH13 is methylated in Pediatric AML patients
We next examined the methylation status of the CDH13 promoter in pediatric AML samples and NBM/ITP (normal bone marrow/idiopathicthrombocytopenic purpura) control samples. Aberrant methylation of CDH13 was observed 2 (6.7%) in the 30 bone marrow samples from controls and 24 (34.3%) cases among70 pediatric AML samples (Figure 2A). Aberrant methylation of the CDH13 gene could be detected in all FAB subtypes and in all cytogenetic risk groups. There were no significant differences in clinical features, such as sex, age, initial hemoglobin level, white blood cell counts, platelet counts, and chromosomal abnormalities between patients with and without CDH13 methylation (Table 1). The survival time of the patients also has no relationship with the methylation status of CDH13 (Figure 2D).
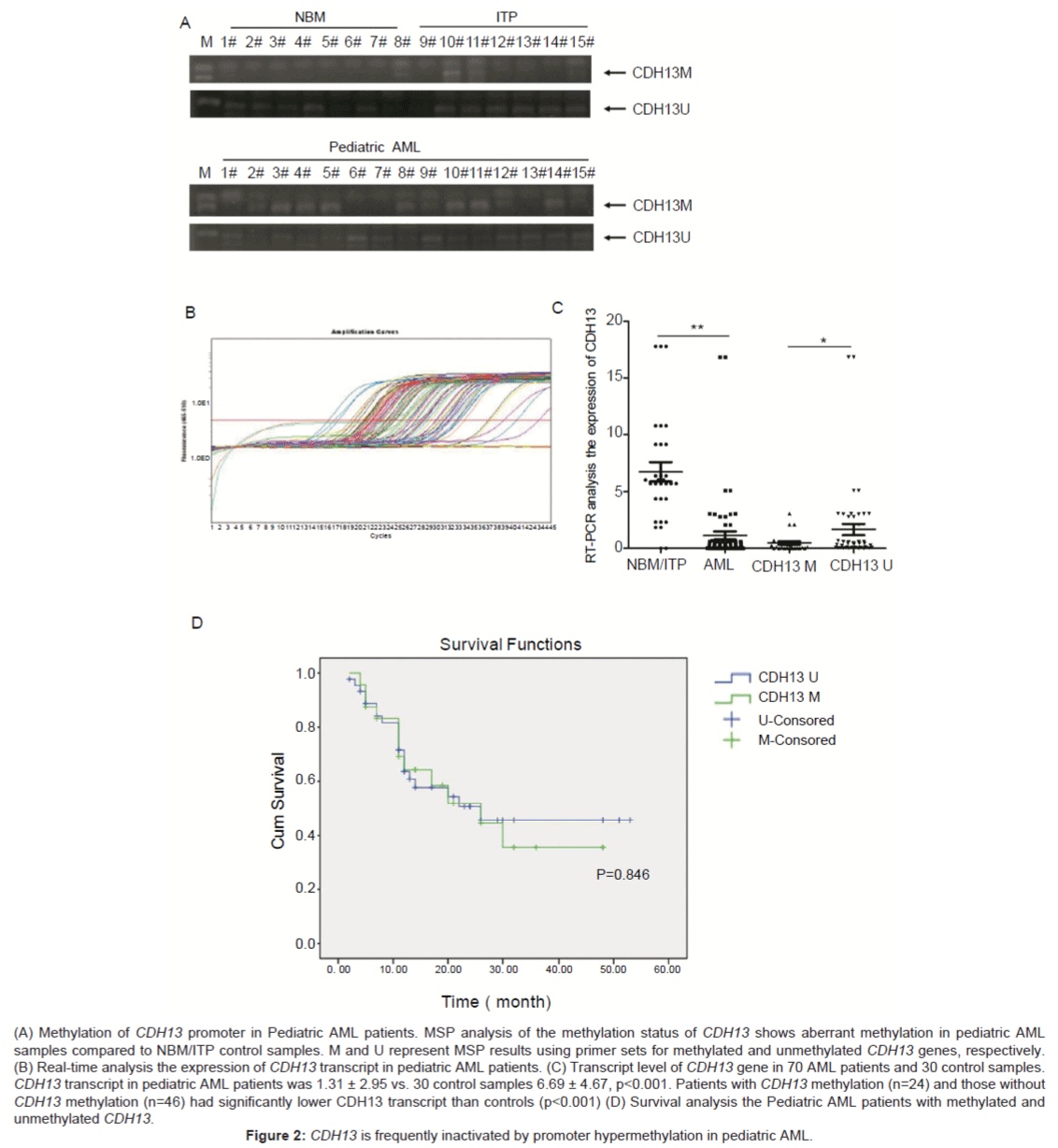
(A) Methylation of CDH13 promoter in Pediatric AML patients. MSP analysis of the methylation status of CDH13 shows aberrant methylation in pediatric AML samples compared to NBM/ITP control samples. M and U represent MSP results using primer sets for methylated and unmethylated CDH13 genes, respectively. (B) Real-time analysis the expression of CDH13 transcript in pediatric AML patients. (C) Transcript level of CDH13 gene in 70 AML patients and 30 control samples. CDH13 transcript in pediatric AML patients was 1.31 ± 2.95 vs. 30 control samples 6.69 ± 4.67, p<0.001. Patients with CDH13 methylation (n=24) and those without CDH13 methylation (n=46) had significantly lower CDH13 transcript than controls (p<0.001) (D) Survival analysis the Pediatric AML patients with methylated and unmethylated CDH13.
Figure 2: CDH13 is frequently inactivated by promoter hypermethylation in pediatric AML.
Expression of CDH13 transcript in pediatric AML patients
The transcript level of CDH13 gene was examined in 70 AML patients with available materials using Real-time PCR. As shown in table 1, CDH13 expression was significantly decreased in AML patients (1.31 ± 2.95) compared to 30 NBM/ITP controls (6.69 ± 4.67, p<0.001); Both patients with CDH13 methylation (n=24) and those without CDH13 methylation (n=46) had significantly lower CDH13 transcript than controls (p<0.001) (Figures 2B and 2C). Furthermore, CDH13 transcript was significantly lower in patients with methylated CDH13 than those without methylated CDH13 (p=0.036) (Table 1).
CDH13, instances of both hypermethylation and loss of function have been documented in numerous cancers. In our study, hypermethylation of the CDH13 promoter was detected in 5 of 9 (55.6%) leukemia cell lines. A high frequency (34.3%) of CDH13 promoter hypermethylation was also found in pediatric AML primary tumor cells, which implied that silencing of the CDH13, may be involved in the tumorigenesis of pediatric AML. Promoter hypermethylation of CDH13 is frequently found in AML, but it is not associated with sex, age and patient’s survival in our series. This is inconsistent with observations in non-small-cell lung cancer: a high methylation rate of CDH13 is generally associated with poor prognosis in lung cancers [23], but consistent with colorectal cancers [34].
Several reports have implied the effects of CDH13 gene on the proliferation and apoptosis of cancer cells. Melanoma cells that re-express CDH13 show a reduction in the rate of tumor growth in a nu/nu mouse tumor model [35]. CDH13 over-expression in hepatocellular carcinoma also increases sensitivity of tumor cells to TNFa-induced apoptosis [36]. Positive relationship between CDH13 and Cyclin D2 methylation was reported in prostate cancer [37]. There still no report about the effects of CDH13 in leukemia cells. Leukemia cells may share the same mechanism with solid tumor cells, or maybe there is totally different molecular mechanism in leukemia cells.
Besides the methylation of CDH13, gene mutation and spliced mRNAs may also play important role in pediatric AML. Two single nucleotide polymorphism in distinct introns of CDH13 have been associated with greater sensitivity of lymphoblastoid cell lines to apoptosis induced by cisplatin and daunorubicin [38]. CDH13 produces a lot of spliced mRNAs, many of these encode proteins are predicted to be secreted and thus, like proteolytic fragments of the major CDH13 isoforms, might function as extracellular ligands. Until now little is known about these smaller proteins. Next step, we will explore the relationship between mutation/spliced mRNA of CDH13 and pediatric leukemia.
This work demonstrated that inactivation of CDH13 by promoter hypermethylation is a tumor specific and frequent event in pediatric AML.
This work was supported by grants from the National Key Basic Research Program No. 2010CB933902, National Natural Science Foundation for youth No. 81100371, Natural Science Foundation of Jiangsu Province No. BK2011308, Universities Natural Science Foundation of Jiangsu Province No. 11KJB320014 and Talent’s subsidy project in science and education of department of public health of Suzhou City No. SWKQ1020. Medical innovation team and leading talent of Jiangsu Province. No. LJ201126. Major scientific and technological special project for “significant new drugs creation” No. 2012ZX09103301-040.
The authors have no conflicts of interest to disclose.

Additional file 1: Bisulfite genomic sequencing of HL-60 cells.
| Pediatric AML | ||||
|---|---|---|---|---|
| NBM/ITP | Pediatric AML | CDH13 methylated | CDH13 unmethylated | |
| 1 | 5.985902 | 0.009562268 | 0.009562268 | 0.4312417 |
| 2 | 10.75226 | 0.008266937 | 0.008266937 | 16.81374 |
| 3 | 17.77242 | 0.5850244 | 0.5850244 | 2.773232 |
| 4 | 5.663008 | 3.045267 | 0.009562268 | 5.086131 |
| 5 | 4.351668 | 0.5850244 | 0.5850244 | 0.4312417 |
| 6 | 5.682669 | 0.2729235 | 0.2729235 | 2.094446 |
| 7 | 2.307881 | 0.3442596 | 0.3442596 | 0.05243117 |
| 8 | 0.000212032 | 0.0282926 | 0.0282926 | 0.05170934 |
| 9 | 9.136038 | 0.6743495 | 0.6743495 | 0.05170934 |
| 10 | 1.823321 | 0.008266937 | 0.008266937 | 0.07363669 |
| 11 | 6.347538 | 0.09615919 | 0.09615919 | 0.07363669 |
| 12 | 5.985902 | 0.3741189 | 0.3741189 | 3.045267 |
| 13 | 10.75226 | 2.992951 | 0.008266937 | 3.045267 |
| 14 | 17.77242 | 0.1143532 | 0.1143532 | 0.5850244 |
| 15 | 5.663008 | 0.06990641 | 0.06990641 | 3.045267 |
| 16 | 4.351668 | 0.003992667 | 0.003992667 | 0.5850244 |
| 17 | 5.682669 | 0.5850244 | 0.5850244 | 0.2729235 |
| 18 | 2.307881 | 0.01853722 | 0.01853722 | 0.3442596 |
| 19 | 0.000212032 | 0.04057032 | 0.04057032 | 0.0282926 |
| 20 | 9.136038 | 0.2351362 | 0.2351362 | 0.6743495 |
| 21 | 1.823321 | 3.045267 | 3.045267 | 0.008266937 |
| 22 | 6.347538 | 2.094446 | 2.094446 | 0.09615919 |
| 23 | 5.985902 | 0.5402011 | 0.5402011 | 0.3741189 |
| 24 | 10.75226 | 3.045267 | 3.045267 | 2.992951 |
| 25 | 17.77242 | 0.4312417 | 0.1143532 | |
| 26 | 5.663008 | 16.81374 | 0.06990641 | |
| 27 | 4.351668 | 2.773232 | 0.003992667 | |
| 28 | 5.682669 | 5.086131 | 0.5850244 | |
| 29 | 2.307881 | 0.4312417 | 0.01853722 | |
| 30 | 9.136038 | 2.094446 | 0.04057032 | |
| 31 | 0.05243117 | 0.2351362 | ||
| 32 | 0.05170934 | 0.002774741 | ||
| 33 | 0.05170934 | 0.0106838 | ||
| 34 | 0.07363669 | 0.5402011 | ||
| 35 | 0.07363669 | 0.4868565 | ||
| 36 | 0.009562268 | 0.4312417 | ||
| 37 | 0.008266937 | 16.81374 | ||
| 38 | 0.5850244 | 2.773232 | ||
| 39 | 3.045267 | 5.086131 | ||
| 40 | 0.5850244 | 0.4312417 | ||
| 41 | 0.2729235 | 2.094446 | ||
| 42 | 0.3442596 | 0.05243117 | ||
| 43 | 0.0282926 | 0.05170934 | ||
| 44 | 0.6743495 | 0.05170934 | ||
| 45 | 0.008266937 | 2.992951 | ||
| 46 | 0.09615919 | 3.045267 | ||
| 47 | 0.3741189 | |||
| 48 | 2.992951 | |||
| 49 | 0.1143532 | |||
| 50 | 0.06990641 | |||
| 51 | 0.003992667 | |||
| 52 | 0.5850244 | |||
| 53 | 0.01853722 | |||
| 54 | 0.04057032 | |||
| 55 | 0.2351362 | |||
| 56 | 0.002774741 | |||
| 57 | 0.0106838 | |||
| 58 | 0.5402011 | |||
| 59 | 0.4868565 | |||
| 60 | 0.4312417 | |||
| 61 | 16.81374 | |||
| 62 | 2.773232 | |||
| 63 | 5.086131 | |||
| 64 | 0.4312417 | |||
| 65 | 2.094446 | |||
| 66 | 0.05243117 | |||
| 67 | 0.05170934 | |||
| 68 | 0.05170934 | |||
| 69 | 2.992951 | |||
| 70 | 3.045267 | |||
| AV | 6.698232841 | 1.310098949 | 0.533199123 | 1.716575293 |
| SD | 4.672074863 | 2.959071041 | 0.892060633 | 3.540122523 |
Real-time PCR analysis the expression of CDH13 gene in NBM and pediatric AML patients.