Journal of Hematology & Thromboembolic Diseases
Open Access
ISSN: 2329-8790
ISSN: 2329-8790
Review Article - (2014) Volume 2, Issue 5
Soluble cytokine receptors are products of tumor cells infected with viruses such as human T-cell leukemia virus type 1 (HTLV-1). Adult T-cell leukemia/lymphoma (ATLL) is a highly aggressive leukemia/lymphoma caused by HTLV-1. High levels of soluble cytokine receptors are detected in the serum of patients with ATLL. Matrix metalloproteinase (MMP)-9 cleaves the interleukin (IL)-2 receptor α chain (IL-2Rα, CD25), which produces soluble IL-2Rα (sIL-2R) and down-regulates the proliferative capability of T cells that encounter cancer cells. Soluble CD30 (sCD30) impairs the interaction of CD30 ligand+ cells with CD30+ cells, whereas CD30 expression integrates survival signals to NK cells. Accordingly, high level of sIL-2R and sCD30 suggest that not only does impaired tumor surveillance lead to the survival of ATLL cells, but also dysfunction of immune surveillance can cause severe infectious complications. Freshly isolated leukemic cells from acute type ATLL patients express little CD30 on the surface, and predominantly produce sCD30. Proliferating ATLL cells are thought to release sCD30, and excess sCD30 probably blocks CD30L on normal activated T cells and myeloid cells, protecting CD30+ ATLL cells from apoptosis. Regarding CD30, two types of MMPs have been reported, ADAM10 and ADAM17, and Tax expression may be associated with ADAM17 activation in HTLV-1-infected cells. Thus, soluble cytokine receptor, sCD30, is both a serum component and a functional protein. Detection of sCD30 level may be useful for detecting and monitoring Tax-expressing tumor cells in vivo.
Keywords: Adult T-cell leukemia/lymphoma (ATLL); Human T-cell leukemia virus type 1 (HTLV-1); soluble CD30 (sCD30); A disintegrin and metalloproteinase (ADAM)10; ADAM17
Adult T cell leukemia/lymphoma (ATLL) is an aggressive leukemia/lymphoma which was first proposed as a new disease entity in 1977 [1,2]. The interaction between human T-cell leukemia virus type 1 (HTLV-1)-infected cells with different types of cells appears to be one of the basic mechanisms underlying the development of HTLV-1-associated diseases [3]. This interaction may play a major role in determining and mediating clinical features of the diseases. ATLL is classified into at least four clinical subtypes: acute, chronic, smoldering, and lymphoma-type. This classification is based on the number of abnormal T cells in peripheral blood, serum lactic acid dehydrogenase level, tumor lesions in various organs, and the clinical course. Major complications of ATLL are hypercalcemia and serious infections including Pneumocystis jirovecii pneumonia, aspergillosis or candidiasis, and cytomegalovirus pneumonia. Other diseases reported to be associated with HTLV-1 infection include chronic respiratory diseases. Monoclonal proliferation of HTLV-1-infected cells is observed in some virus carriers, who are considered to be at high risk for development of ATLL [4]. Constitutive activation of signal transducer and activator of transcription (STAT) as well as functional impairment and stabilization of p53 protein found in the peripheral blood mononuclear cells (PBMCs) of ATLL patients may be one basis for ATLL development [5,6]. However, the additional factors that lead to the severe clinical conditions of ATLL patients remains to be determined.
Interleukin-2 (IL-2) is a T-cell growth factor and essential in the survival and expansion of CD4+CD25+ activated T cells in PB [7]. Among IL-2 receptor (IL-2R) complex (α, β, and γ chains), matrix metalloproteinase (MMP)-9 mediates cleavage the IL-2R α chain (IL-2Rα, CD25), which produces soluble IL-2Rα (sIL-2R) [8]. CD30, a 120 kDa type I cell surface glycoprotein, is an unusual member of the tumor necrosis factor receptor superfamily, with a membrane form and a soluble form of CD30 (sCD30) by tumor virus cells and viral infection [9]. CD30 expression depends on mitogen or viral activation and proliferation of B and T cells and tumor cells from Hodgkin lymphoma and anaplastic large cell lymphoma express CD30 [10]. Thus levels of sCD30 and sIL-2R are elevated in HTLV-1 carriers and highly elevated in ATLL [11]. Furthermore, sCD30 elevation is followed by acute crisis in the indolent type of ATLL (unpublished data). Our findings suggest that soluble cytokine receptors may play an important role in the development ATLL.
The poor prognosis of ATLL appears to result not only from the high frequency of drug-resistant ATLL cells, but also from a secondary immune deficiency, which results in a variety of opportunistic infections. Such immune suppression has also been observed in asymptomatic HTLV-1 carriers, who often show an increased frequency of cancer, and expansion of carcinoma metastases in situ [12]. In this context, our previous observation that monocyte-derived dendritic cells are functionally impaired in HTLV-1 carriers may be important [13]. In addition, it was reported that MMP-9 mediates cleavage of IL-2Rα and down-regulated the proliferative capability of cancer-encountered T cells [14], whereas HTLV-1-infected cells transit from IL-2-dependent to IL-2-independent growth via the acquisition of a constitutively activated STAT proteins [15] (Figure 1). On the other hand, expression of CD30 integrates survival signal to natural killer cells, allowing them to prevent viral spread and subsequent disintegration of secondary lymphoid tissue [16]. Accordingly, high levels of sIL-2R and sCD30 suggest that not only can impaired tumor surveillance lead to the survival of ATLL cells, but also dysfunction of immune surveillance can cause severe infectious complications.
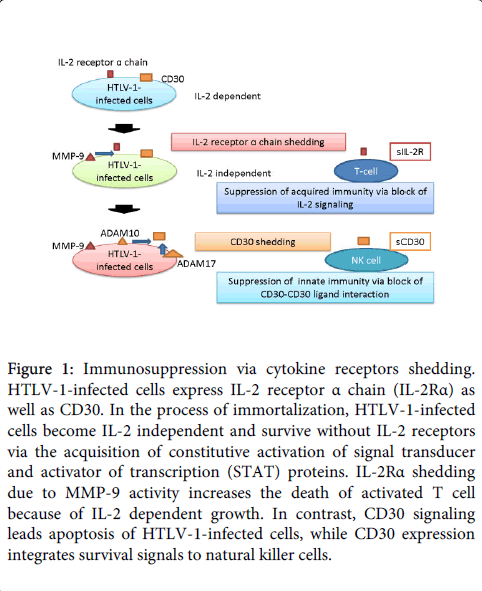
Figure 1: Immunosuppression via cytokine receptors shedding. HTLV-1-infected cells express IL-2 receptor α chain (IL-2Rα) as well as CD30. In the process of immortalization, HTLV-1-infected cells become IL-2 independent and survive without IL-2 receptors via the acquisition of constitutive activation of signal transducer and activator of transcription (STAT) proteins. IL-2Rα shedding due to MMP-9 activity increases the death of activated T cell because of IL-2 dependent growth. In contrast, CD30 signaling leads apoptosis of HTLV-1-infected cells, while CD30 expression integrates survival signals to natural killer cells.
MMPs represent the most prominent family of proteinases associated with tumorigenesis [17]. MMPs are enzymes that mediate tissue destruction and cell infiltration as well as shedding of cell surface cytokine and cytokine receptors. In addition, MMPs regulate signaling pathways that control cell growth, inflammation, or angiogenesis. Regarding CD30, two types of MMPs have been reported, ADAM10 and ADAM17 [18,19]. For instance, bortezomib causes CD30 shedding and human anti-CD30 antibody (Ab) simultaneous or preincubation with bortezomib fails to improve Ab therapy. Interestingly, CD30 ligation activates ADAM10 but not ADAM17, while ADAM17 is initiated by the protein kinase C (PKC) activation (Figure 2). CD30 shedding has been speculated to be controlled in multiple pathways, including those that activate ADAM10 and ADAM17.
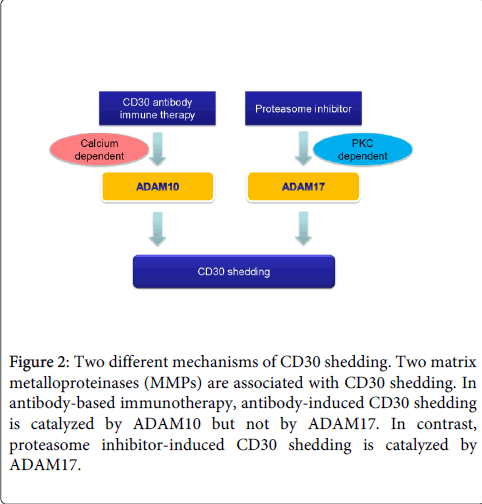
Figure 2: Two different mechanisms of CD30 shedding. Two matrix metalloproteinases (MMPs) are associated with CD30 shedding. In antibody-based immunotherapy, antibody-induced CD30 shedding is catalyzed by ADAM10 but not by ADAM17. In contrast, proteasome inhibitor-induced CD30 shedding is catalyzed by ADAM17.
We previously reported that ATLL cells from patients expressed CD30, but that freshly isolated leukemic cells from acute type ATLL patients express little CD30 on the surface (Figure 3). Because CD30 is expressed by activated T cells, we first investigated CD25 and CD30 expression in PBMCs from healthy donors and patients with ATLL following stimulation with PHA-P for 72 h (Figure 4A and 4B). PBMCs from healthy donors express little or no CD25 or CD30 in the absence of PHA-P stimulation and CD25+CD30+ cells appear after stimulation. In contrast, CD25+CD30+ cells are detected in PBMCs from ATLL patients in the absence of stimulation. The results for CD4+CD30+ cells were similar to those for CD25+CD30+ cells. Next, we examined sCD30 levels in culture supernatants (Figure 4C). Higher sCD30 level were detected in the culture supernatant of PBMCs from ATLL patient with PHA-P stimulation compared to without stimulation, whereas low sCD30 levels were detected in healthy donors even with stimulation. We also found that strong stimulation with PHA-P (and anti-CD3) suppressed the proliferation of PBMCs from ATLL patients (and HTLV-1 carriers) (Supplementary figure 1 and 2). Thus, the responses are different between PBMCs from ATLL patients (HTLV-1 carriers) and healthy donors under condition of T cell activation. Our findings suggest that ATLL cells predominantly express sCD30, which plays a pivotal role in ATLL tumorigenesis.

Figure 3: Very low number of CD4+CD30+ cells in peripheral blood of patients with acute type ATLL. A small number of CD30- expressing CD4+ cells in PBMCs was observed. A few CD4+ cells express CD30 in PBMCs from acute type ATLL cases. MT-2 (Taxexpressing HTLV-1-infected) cells express the membrane form and soluble form of CD30. The Hodgkin/Reed-Sternberg cell line, L-540, and the anaplastic large cell lymphoma cell line, Karpas 299, were used as positive controls. ATLL5 is a patient that is therapy resistant; ATLL7 is a patient that relapsed.
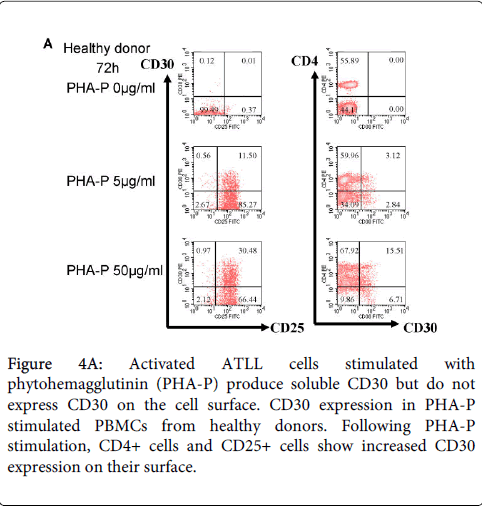
Figure 4A: Activated ATLL cells stimulated with phytohemagglutinin (PHA-P) produce soluble CD30 but do not express CD30 on the cell surface. CD30 expression in PHA-P stimulated PBMCs from healthy donors. Following PHA-P stimulation, CD4+ cells and CD25+ cells show increased CD30 expression on their surface.
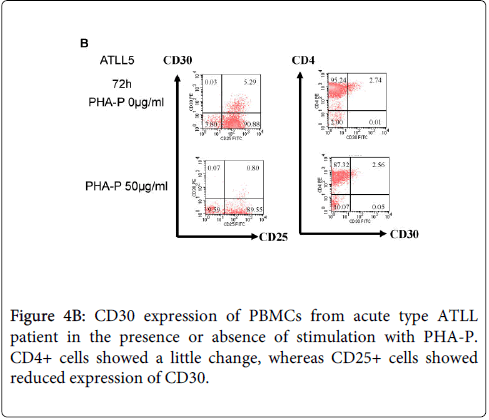
Figure 4B: CD30 expression of PBMCs from acute type ATLL patient in the presence or absence of stimulation with PHA-P. CD4+ cells showed a little change, whereas CD25+ cells showed reduced expression of CD30.
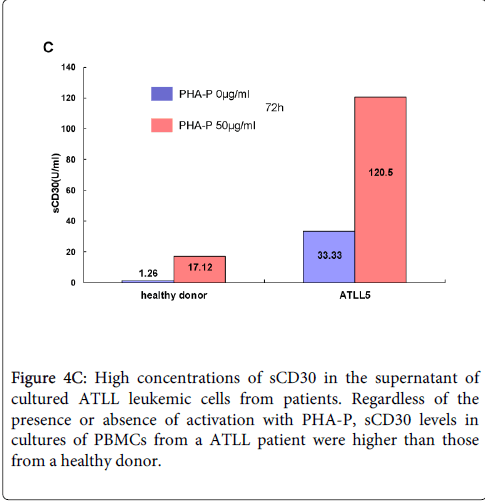
Figure 4C: High concentrations of sCD30 in the supernatant of cultured ATLL leukemic cells from patients. Regardless of the presence or absence of activation with PHA-P, sCD30 levels in cultures of PBMCs from a ATLL patient were higher than those from a healthy donor.
More than 50 years are required for HTLV-1 carriers to develop ATLL, but the reasons for this long lag period are not known. The phenotype of ATLL cells is mature CD4+CD25+ T cells. HTLV-1-infected T cells produce cytokines and soluble cytokine receptors. Subsequently, soluble cytokine receptors may support clonal proliferation and invasion of ATLL cells, creating a tumor microenvironment.
sCD30 binds to CD30 ligand (CD30L) with high affinity, decreases the availability of CD30L on lymphocytes, and blocks apoptosis of CD30+ tumors [20]. Su et al. also reported that CD30 inhibits T-cell proliferation via a CD30 signaling pathway [21]. CD30L is expressed on activated T cells, granulocytes, eosinophils as well as neoplasms with an activated peripheral T-cell phenotype including ATLL (2 positive / 2 tested) [22]. In our patient with acute type ATLL, sCD30 levels increase according to the increasing number of ATLL cells. However, we detected no CD30L expression of leukemic cells from PB (data not shown). Thus, proliferating ATLL cells are thought to release sCD30, and excess sCD30 probably blocks CD30L on normal activated T cells and myeloid cells, protecting CD30+ ATLL cells from apoptosis.
Tax is a 40-kDa nuclear phosphoprotein that transactivates the transcription of HTLV-1 genome and a variety of cellular genes such as IL-2 and IL-2Rα [23]. Tax interacts with PKC and stimulates membrane translocation and triggering of the PKC pathway [24]. Actually, Okada S, et al. showed that sCD30 levels are 5 times higher in supernatant of MT-2 (Tax highly expressing cell line) than MT-4 (Tax lower expressing cell line), whereas CD30 was highly expressed on the surfaces of MT-4 [25,26]. Furthermore, Rabson AB, et al. reported that T cell activation cooperates with Tax for cellular gene expression [27,28]. It is suggested that enhancing sCD30 production results from synergistic activation by PHA and Tax (Figure 4C). Thus, Tax expression may be associated with ADAM17 activation in HTLV-1-infected cells (Figure 5). On the other hand, sCD30 itself may stimulate CD30 shedding via sCD30/anti-CD30 antibody immune complexes. sCD30/CD153 (CD30 ligand) immune complex may also play a role.
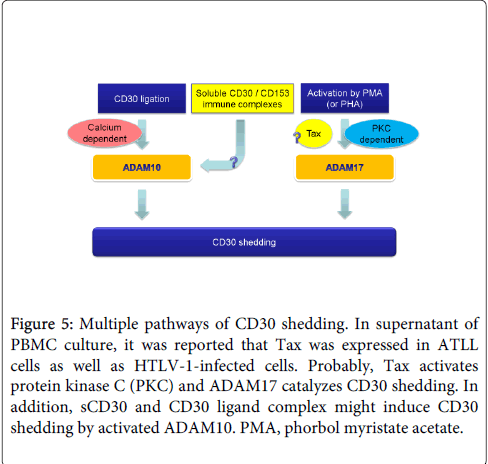
Figure 5: Multiple pathways of CD30 shedding. In supernatant of PBMC culture, it was reported that Tax was expressed in ATLL cells as well as HTLV-1-infected cells. Probably, Tax activates protein kinase C (PKC) and ADAM17 catalyzes CD30 shedding. In addition, sCD30 and CD30 ligand complex might induce CD30 shedding by activated ADAM10. PMA, phorbol myristate acetate.
The soluble cytokine receptor, sCD30, is not only a serum component but also a functional protein. The serum level of sCD30 could be used to detect and monitor Tax-expressing tumor cells in vivo.
We would like to thank the member of the Department of Hematology at NHO Kumamoto Medical Center and the Department of Hematology and Respiratory Medicine, Kochi Medical School, Kochi University for collaboration.