Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2012) Volume 2, Issue 1
rDNA-ITS: ribosomes DNA-internal transcribed space; dNTP: Deoxy-ribonucleoside Triphosphate; CTAB: Cetyl Trimethyl Ammonium Bromide; EDTA: Ethylene Diamine Tetra Acetic Acid; PDA: Potato Dextrose Agar; PSA: Potato Sugar Agar
Apple is one of the most important fruit species and is economically important worldwide [1]. Apple ring rot disease (Physalospora piricola Nose.), which is one type of fungal disease caused by Botryosphaeria berengeriana de Not f. sp. piricola (Nose) Koganezawa & Sakuma, is widely distributed in warm apple growing regions of the world [2]. In recent years, the incidence of ring rot disease in major apple yielding regions has been increasing sharply due to the expansion of the planted areas and prolonged planting, which has resulted in a significant loss of apple production. In normal years, the morbidity of fruits is typically 20%-30%; however, the rate can reach 50%-60% in years with increased morbidity [3,4]. Adding to the seriousness of this pathogenic fungus, its ability to cause latent infections not only leads to the infection of field plants but also to the induction of the postharvest decay of fruits, which causes significant losses during the storage period [5,6].
Research on the common diseases such as ring rot has become one of the popular topics in industrialised apple production [7]. Over the past 30 years, systematic studies on the pathological mechanisms of Physalospora piricola, its susceptibility to pesticides and the effects of chemical prevention have been conducted [8]. However, long-term chemical prevention has resulted in some fungal pathogens developing resistance to these pesticides. Natural selection and evolution allow the pathogens to adapt themselves to variations in different plant species, environmental changes and prevention methods [9]. This adaptation adversely affects pathogenesis research, pathological changes, pathogen isolation and identification, disease control, and screening for diseaseresistant cultivars [10,11].
Together with the rapid development of molecular biology techniques, rDNA-ITS sequence analyses have been widely used in pathogen identification and systems development research, including studies on Colletotrichum, Puccinia and Phytophthora [12-21], which may provide new pathways for advanced research on Physalospora piricola in molecular biology field. In Henan province and Liaoning province, as the traditional major apple producing areas in north China, the apple production was seriously affected by apple ring rot disease. In that case, much research on Physalospora piricola had been performed by tow fruit research institutes in Henan and Liaoning. The experiment orchards had been established locally, in which resistance germplasm resources had been screened and varieties of apple ring rot pathogen strains had been cultured. This study was cooperated with these tow institutes, and followed with the program requirements, ten pathogenic strains of Physalospora piricola were collected in Henan and Liaoning. Using fungal genomic DNA extractions, PCR amplification of rDNA ITS sequences, and bioinformatics analyses, we wish to identify the genetic relationship between different strains and investigate the variations, which might preliminary establish the method of ITS sequences test to classified and variation research on Physalospora piricola. The in vitro cultivation and pathogenicity tests for variant strains were performed to examine any correlations between sequence variations and their biological characteristics or pathogenicities, in the expectation of that it could lay a foundation for the future research on the rules that govern variation and the pathogenic mechanisms of Physalospora piricola [22,23].
Resistance breeding for apples using biotechnology methods such as transforming resistance related genes has yielded some interesting results [10]. Many people have made great efforts to promote resistance germplasm resources screening, verification of transgenic plants function, disease prevention technology, for which huge amounts of conidiospore was necessary as experimental material [14]. Because the rate of conidiospore induction for apple ring rot pathogens is low through in vitro cultivation, we screened for the effective culture conditions for the selected strain that would provide better conidium producing ability. And we wish the result would be beneficial to the resistance testing of transgenic plants, a bioassay of effective fungicides and their experimental pharmacological efficacy against fungi infections in the field or in storage. Thus, these new materials and methods could effectively guide apple production practices [24,25].
Materials
Fungi strains: Ten pathogenic strains of apple ring rot disease were collected in Henan province and Liaoning province from a variety of tissues or cultivars and from different districts (Table 1), and isolate purified by Zhengzhou Fruit Research Institute, China Academy of Agricultural Science (CAAS). The general culture media used for inoculation and preservation were potato dextrose agar (PDA) (potato 200 g/l, glucose 20 g/l, agar 20 g/l) or potato sugar agar (PSA) (potato 200 g/l, sucrose 20 g/l, agar 20 g/l).
| Numbers | Collecting location | Species | Varieties | Tissue | Shape |
| ZZ-01 | Zhengzhou Fruit Research Institute Orchard South Park | Apple | Fuji | Bark | Tumor |
| ZZ-12 | Stone Buddha Orchards, western suburbs of Zhengzhou | Apple | Zaohong | Bark | Tumor |
| ZZ-13 | Stone Buddha Orchards, western suburbs of Zhengzhou | Apple | Zaohong | Bark | Tumor |
| ZZ-23 | Zhengzhou Shilipu Orchard | Apple | Tengmu 1 | Bark | Tumor |
| ZZ-25 | Zhengzhou Shilipu Orchard | Apple | Tengmu 1 | Bark | Tumor |
| ZZ-35 | Zhengzhou Shilipu Orchard | Apple | Redchief | Bark | Tumor |
| ZZ-51 | Zhengzhou Shilipu Orchard | Apple | Redchief | Bark | Lesion |
| ZZ-53 | Nancao Village, Nancao Township, Zhengzhou City | Apple | Golden Delicious | Fruit | Lesion |
| ZZ-55 | Nancao Village, Nancao Township, Zhengzhou City | Apple | Golden Delicious | Fruit | Lesion |
| LW-48 | Wafangdian, Liaoning Province | Apple | Ralls Genet(Kokkō) | Bark | Tumor |
Table 1: Apple ring rot pathogen strains.
Enzymes, reagents, sequencing and primer synthesis: Taq polymerase, dNTPs, pMD18-T vector, and gel extraction kits were purchased from TaKaRa Co.. Sequencing was performed by Huada Co., and the primers were synthesised by Yingjun Biotech, Co.
Plant material: Plant material consisted of fresh, similarly shaped Fuji apples (longitudinal diameter × transect diameter ≈ 6 cm × 7 cm).
Methods
PCR amplification and sequence analysis of the rDNA-ITS of the apple ring rot pathogen strains:
I. Extraction of fungal genomic DNA: Pathogenic fungi mycelia from ten strains were inoculated into PDA medium (pH 6.0) and then transferred into an incubator (25°C) under 24 h / d fluorescent (λ= 400-750 nm) lighting cultivation for 7 days. The fungal genomic DNA was isolated using the cetyltrimethylammonium bromide (CTAB) procedure: 1.0 g of mycelia was collected to a centrifuge tube (2.0 ml); after rapid grinding in liquid nitrogen, 1.0 ml of CTAB solution (0.1 M Tris-HCl, 2.0% w/v CTAB, 1.4 M NaCl, 0.5 mM EDTA, 1.0% w/v β-mercaptoethanol) was added to the tube; after a homogenization, the tube was transferred to 30 min water bath (65°C) and inverted every 5 min. After being cooled to room temperature, an equivalent volume chloroform: isoamyl alcohol mixture (24:1 v/v) was added to the homogenate; the new mixture was shaken for 5 min and followed with centrifuging (12000 rpm, 10 min); then absorb supernatants and repeat the extraction with chloroform: isoamyl alcohol mixture (24:1 v/v). The upper, aqueous, DNA containing phase was transferred to a fresh centrifuge tube; then 1/10 volume sodium acetate (NaAC) and double volume ethanol were added and incubated at -20°C for 1 h. The precipitated DNA was pelleted after the centrifugation (10000 rpm, 10 min); the tube was washed twice with 70% ethanol; after air drying, DNA was dissolved with 100 μl ddH2O [26].
II. PCR amplification, cloning and sequencing of the ITS region: The internal transcribed spacer region (ITS) was amplified with universal primers ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) and ITS4 (5’-TCCTCCGCTTATTGATATGC-3’). The PCR reaction mixture (25 μl) included: 2.5 μl 10 × PCR Buffer, 2 μl dNTP, 1 μl primer (ITS1), 1 μl primer (ITS4), 0.2 μl rTaq enzyme, 1 μl template DNA, and 18.3 μl ddH2O. The reaction condition was: 95°C predegeneration for 4 min; (94°C 30 s, 56°C 30 s, 72°C 1 min) × 35 cycles; 72°C elongation for 4 min. The PCR products were detected through agarose gel electrophoresis. After recovery and purification, the DNA fragments were ligated to pMD18-T vector, and then transformed into competent Escherichia coli DH5α cells. Positive clones were screened using white-blue plaque selection on LB agar plates and then cultured in LB liquid medium. After PCR verification, positive clones were sequenced by Huada Gene Co.
III. Bioinformatic analysis of ITS sequences: The homologies between the cloned ITS sequences of ten ring rot pathogen strains were analysed with DNAMAN 5.22, and the homology alignment of the nucleotide sequences was analysed by BLASTn on NCBI (https:// blast.ncbi.nlm.nih.gov/Blast.cgi). A phylogenetic tree was simply constructed using Cluster 1.83 and MEGA 4.0.
Comparisons of biological characteristics of the different apple ring rot strains in in vitro cultures:
I. Growth status of fungal colony: Based on the sequencing results, ZZ01 was selected as the control group because its rDNA-ITS sequence had not been changed. Five pathogenic strains ZZ23, ZZ25, ZZ35, ZZ55 and LW48 with the base-pair mutation and ZZ01 were inoculated onto PDA medium (pH 6.0) in glass plates (Φ = 8 cm, same below). The dishes were cultured in an incubator (25°C) for 7 d under black light (24 h/d). The growth morphology of the colony was determined every 24 h, and the diameter of the mycelia was measured using the crossing method using a randomised block design and repeated 3 times.
II. Ability to induce conidiosporangia and spores: After a 7-d cultivation, the mycelium from the above colonies was scraped with an inoculating loop on day 8 and then allowed to continue cultivating for another 14 d under the same conditions. The total number of conidiosporangia for the different strains was counted on day 21, and the spores were examined using the following method:
For each strain, 30 similarly shaped conidiosporangia were collected from each dish and then slightly grinded in 2 ml of sterile water with a mortar to produce spore suspensions. For counts, 0.5 μl of a suspension was blotted onto a glass slide, and the spore morphology was observed under a microscope at low-magnification (10×10) and high-magnification (10×40). The concentration of the suspension was determined using a haemocytometer (25×16) [27]. The analysis of variance was performed for the data using the Duncan’s Multiple Range Test or Student’s t-test with SAS 6.0. (Same below)
III. Pathogenicity of different strains towards apples: The five pathogenic strains (ZZ23, ZZ25, ZZ35, ZZ55 and LW48) and ZZ01 (control strain) were inoculated onto PDA medium (pH 6.0) in glass plates. The dishes were cultured in an incubator (25°C) for 7 d under black light (24 h/d). On day 8, fungal mycelia were collected from the similar places on the colonies with a puncher (Φ = 3 mm) and then inoculated onto Fuji apples that were previously sterilised with 70% ethanol. The infected fruits were maintained in an incubator (25°C) for 20 d under black light (24 h/d). The growth diameter of the disease spots was measured every 24 h for the first 7 d, and the degree of decay was measured for 21 d.
In vitro culture techniques for pathogenic fungi to produce conidiosporangia and spores:
I. Comparison between two culture mediums: Two types of potato media were prepared (PDA and PSA, pH 6.0). The pathogenic strain ZZ35 was inoculated onto both kinds of media in glass plates. This strain was selected because of the results from the biological characteristic comparisons in which it performed better. The dishes were cultured in an incubator (25°C) for 7 d under an uninterrupted black light (24 h/d). The growth morphology of the colony was determined every 24 h, and the diameter of the mycelia colony was measured using the crossing method. The mycelium of the above colonies was scraped with the inoculating loop on day 8 and then cultivated for another 14 d under the same conditions. The number of conidiosporangia was counted on day 21, and the conidiospores was examined from the same suspensions.
II. Affects of potato medium with different pH values: Using pasteurised NaOH (1 M) or HCl (1 M), the pH value of the PDA medium was adjusted to four different values for new treatments: pH 5.0, 6.0, 7.0 and 8.0. ZZ35 was inoculated onto the respective media and cultivated for 7 d in an incubator (25°C) under continuous black light (24 h/d). The mycelia were processed as described above.
III. Influence of temperature on in vitro cultures: After being inoculated onto PDA medium (pH 6.0), the pathogenic colonies were cultured for 7 d under four constant temperature conditions (5°C, 15°C, 25°C and 30°C) and continuous black light (24 h/d). Subsequently, the mycelia were processed as described above.
IV. Impact of lighting sources and conditions: Four different combinations of lighting sources and conditions were placed in an incubator (25°C): 24 h/d fluorescent light, 24 h/d black light, (12 h fluorescent light / 12 h black light) /d and 24 h/d darkness. ZZ35 was inoculated onto PDA medium (pH 6.0) and cultured under the four different lighting conditions. The mycelia were processed as described above.
V. Effect of scraping treatment on inducing conidiosporangia and spores: The pathogenic strain of apple ring rot disease ZZ35 was inoculated onto the PDA medium (pH 6.0) and cultivated for 7 d in an incubator (25°C) under black light (24 h/d). On day 8, the fungal mycelium were either scraped with an inoculating loop or not scraped. The colonies for the two treatments were then cultivated for an additional 14 d under the same conditions. On day 21, the number of conidiosporangia was counted and the conidiospores were examined from the suspension.
PCR amplification and sequence analysis of rDNA-ITS
I. PCR amplification and sequence analysis: In this study, we used the genome of ten different strains of apple ring rot disease as a template for PCR reactions using the universal primers ITS1 and ITS4, and 10 bands approximately 580 bp in length were amplified. The PCR products were gel purified, cloned and sequenced. The bands from the strains ZZ25, ZZ35 and LW48 were 583 bp in length, whereas the bands from the other seven strains were 582 bp in length (Figure 1). The entire length of the sequences included a partial 18S ribosomal RNA sequence (18S rRNA, 1-30 bp), an internal transcribed spacer 1 sequence (ITS1, 31-210 bp), 5.8S ribosomal RNA sequence (5.8S rRNA, 211-367 bp), an internal transcribed spacer 2 sequence (ITS2, 368-525 bp) and a partial 28S ribosomal RNA (28S, rRNA 526-583 bp) sequence (Figure 2).

Figure 1: Electrophoresis of rDNA-ITS PCR amplification products from the apple ring rot pathogen strains (M: Marker; 1-10: Apple ring rot pathogen strains: ZZ01,ZZ12,ZZ13,ZZ23,ZZ25,ZZ35,ZZ51,ZZ53,ZZ55,LW48).

Figure 2: Comparison of the rDNA-ITS sequences for the ten ring rot pathogen strains (Pane parts show the sequence differences).
The sequence alignment of the ten strains using DNAMAN showed that the primary sequence differences between the strains occurred in the ITS1 region: at 106 bp, the ZZ25, ZZ35 and LW48 sequences had a C, whereas the other seven had a T; and at 135 bp, these same three strains had a C insertion, whereas in the other strains, this base was deleted. Moreover, there was a G to A substitution in LW48 at base-pair 224, a T to C substitution in ZZ55 at base-pair 309, and an A to G substitution in ZZ23 at base-pair 535. However, the ITS region sequences of the other five strains (ZZ01, ZZ12, ZZ13, ZZ51 and ZZ53) were identical (Figure 2). Based on these results, the ten strains could be classified into two categories: the first included ZZ25, ZZ35 and LW48 and the second contained the other seven strains.
II. Comparison of sequence homologies and the construction of a phylogenetic tree: After cloning and sequencing, the sequence between the rDNA-ITS for the ten strains and the type sequence were aligned and analysed using BLASTn provided by NCBI. We found a 99% homology between the test sequences and the sequences of Botryosphaeria berengeriana de Not f. sp. piricola (Nose) (ascension number: GQ865693) and Alternatia sp. (ascension number: GQ855792), thus indicating that these ten strains belong to Botryosphaeria berengeriana de Not f. sp. Piricola (Nose) Koganezawa et Sakuma of Botryosphaeria dothidea, which is classified into two subvarieties, with sub-variety-I originating mostly from China and subvariety- II originating mostly from Europe, such as Italy or Portugal. We can conclude that the intra-species specific evolution is different for various regions.
A phylogenetic tree was constructed from the sequences, that showed a homology ≥ 98% using Cluster 1.83 and MEGA 4.0 (Figure 3). The ten pathogenic strains of apple ring rot disease were classified as sub-variety I. However, ZZ25, ZZ35 and LW48 were more closely related than the other seven strains, and LW48, which was from the Liaoning province, branched separately from ZZ25 and ZZ35 (score not shown).
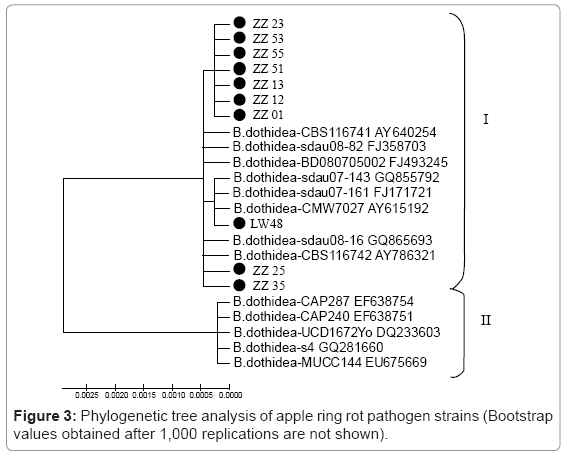
Figure 3: Phylogenetic tree analysis of apple ring rot pathogen strains (Bootstrap values obtained after 1,000 replications are not shown).
Comparison of biological characteristics in in vitro culture conditions for the different apple ring rot strains:
I. Growth rate of the fungal colonies: Based on the daily increase in colony diameter (Figure 4), the growth rate of ZZ35 was the best, the spreading speed of which was faster than the other strains. The growth rate of LW48 was slightly slower than ZZ35 but was better than the other four strains. ZZ25 grew slowly over the first three days but then increased its growth rate after day 4. There was no obvious difference in the growth rate between ZZ55 and ZZ23. ZZ01 and the control strain grew poorly as seen by its slow spreading speed.

Figure 4: Diameter of the colonies of different inoculated strains over a 7-d period (The mycelia diameter was measured using the crossing method every 24 h during the first 7 d post inoculation. Randomised block design, repeated 3 times, same for Figure 7).
II. Ability to induce conidiosporangia and spores: The number of conidiosporangia on the plates was determined for the six strains after a 3-week cultivation period (Table 2). The results showed that the number of sporangia from ZZ35 and LW48 was significantly higher than the other four strains. There were more conidiosporangia for ZZ25 and ZZ55 than for ZZ23 and ZZ01, with no obvious disparities between Z55, ZZ23 and ZZ01 (Figure 5a and 5b).
| Strain | Number of conidia capsule per dish/a | Concentration of spore suspension/105·ml-1 | ||||
| ZZ1 | 7.67* | d** | C*** | 0.00 | - | - |
| ZZ23 | 29.33 | cd | BC | 0.00 | - | - |
| ZZ25 | 113.33 | b | B | 2.67 | c | C |
| ZZ35 | 205.33 | a | A | 8.33 | b | B |
| ZZ55 | 72.33 | bc | BC | 0.50 | d | C |
| LW48 | 200.33 | a | A | 12.17 | a | A |
*Data in table are mean of three repeats. The same below.
**The small letters represent comparison at P<0.05, same letter indicates no significant differences, different letter indicates significant differences. By Duncan’s Multiple Range Test. The same as below.
***The capital letters represent comparison at P<0.01, same letter indicates no significant differences, different letter indicates extremely significant difference. The same below.
Table 2: Comparison of conidiosporangium growth and sporulation capacity of different strains.

Figure 5: Biological status of six strains at different culture stages.
a: Status of ZZ01, ZZ23 and ZZ55
b: Status of ZZ25, ZZ35 and LW48
We could observe via microscopy that there were no spores in the suspensions for ZZ23 or ZZ01, which might suggest that the culture conditions we used were not suitable for the developing conidiosporangia to mature for these two strains. The analysis of variance data for the other four strains with significant variation showed that the concentration of spores in the suspension of LW48 was highest, whereas ZZ55 had the lowest relative concentration (Figure 6).

Figure 6: Spore and mycelia phenotypes of a variety of strains under different magnifications.
III. Pathogenic capacity of different strains on apples: Based on the daily increase in the spot diameters of the six strains over a 7-d period post-infection (Figure 7), the pathogenic capacity of ZZ35 and LW48 was markedly better than the other strains. After day 4, the spreading speed of LW48 exceeded that of ZZ35, while ZZ23 tended to exceed ZZ55 over the same period. The spot spreading speed of ZZ01 was the slowest.
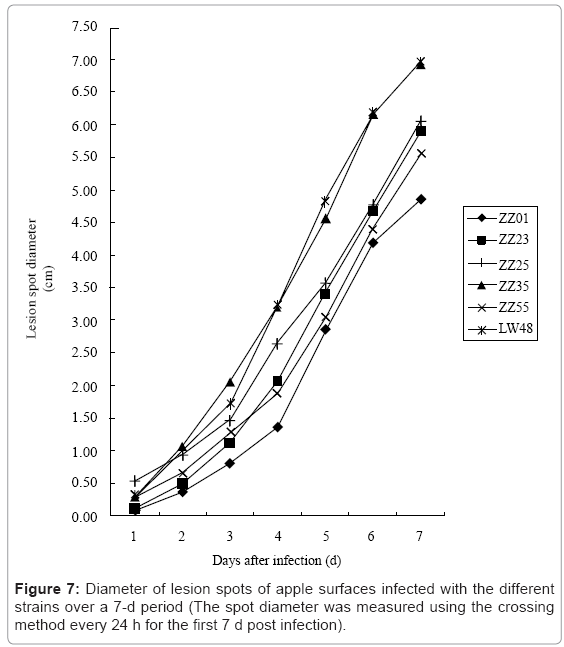
Figure 7: Diameter of lesion spots of apple surfaces infected with the different strains over a 7-d period (The spot diameter was measured using the crossing method every 24 h for the first 7 d post infection).
Using a comprehensive judgment method of grading evaluation over a 15-d period (Table 3), the degree of surface decay of the apples infected with ZZ35 and LW48 was greatest on the same days, whereas that of ZZ01 was the least. On day 4, the lesion spots on the apples infected with ZZ35 and LW48 began to turn black and soft and exudate could be observed in the spots. Similar symptoms were not observed until day 9 for apples infected with ZZ01. The degree of decay was eventually the highest for ZZ25, ZZ35 and LW48 on day 15, whereas that of ZZ01 was far lower (Figure 8).
| Strain/Decay degree | Days after infection | Fastest decay days | |||||||
| 1 | 3 | 5 | 7 | 9 | 11 | 13 | 15 | ||
| ZZ1 | - | - | - | - | +* | + | ++ | ++ | 8th |
| ZZ23 | - | - | - | + | + | ++ | +++ | +++ | 7th |
| ZZ25 | - | - | + | + | ++ | +++ | ++++ | +++++ | 5th |
| ZZ35 | - | - | + | ++ | +++ | ++++ | +++++ | +++++ | 4th |
| ZZ55 | - | - | + | + | + | +++ | +++ | ++++ | 5th |
| LW48 | - | - | + | ++ | ++ | ++++ | +++++ | +++++ | 4th |
*+: The fruit surface begins to perform browning stains and crimple around the infection spots with juice exudates.
++: The spots begin to turn bigger and soft with more juice exudates.
+++: The spots begins to turn black or more brown with irregular fragrance from the fruit.
++++: The black or brown parts have covered half of the fruit surface with foul smell.
+++++: The whole fruit has been changed to brown or black with many cankers.
Table 3: Degree of decay for the apples infected by the different strains.
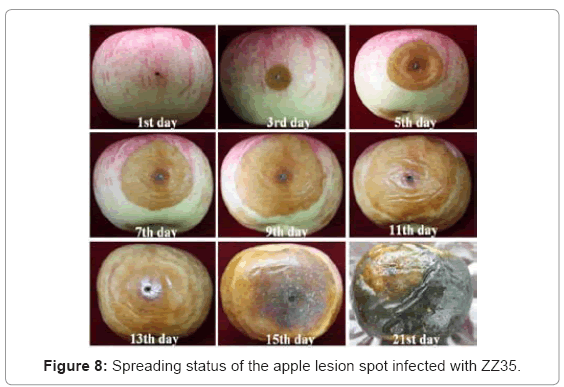
Figure 8: Spreading status of the apple lesion spot infected with ZZ35.
Techniques for the in vitro culture of pathogenic fungi on the production of conidiosporangia and spores
I. Comparison between the effects of two culture media: After a 3-week cultivation on PDA and PSA, the spreading speed of the ZZ35 mycelium on PDA was a little faster than that of PSA over the first four days; however, this discrepancy was not observed later (Figure 9).
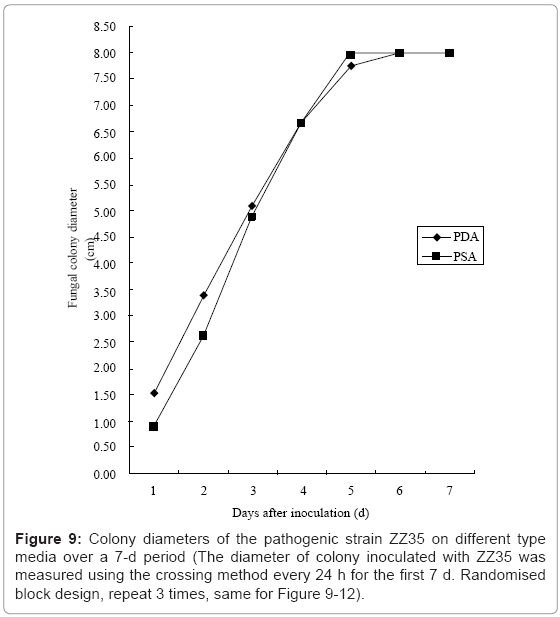
Figure 9: Colony diameters of the pathogenic strain ZZ35 on different type media over a 7-d period (The diameter of colony inoculated with ZZ35 was measured using the crossing method every 24 h for the first 7 d. Randomised block design, repeat 3 times, same for Figure 9-12).
There were fewer conidiosporangia when ZZ35 was grown on PDA compared to PSA. However, there were no significant differences based Student’s t-test for the concentration of the spores in the suspensions extracted from the media with glucose or sucrose (Table 4).
| Medium | Number of conidia capsule per dish/a | Concentration of spore suspension/105·ml-1 | ||||
| PDA | 216.00 | t*=3.1456 | 8.33 | t=2.2188 | ||
| PSA | 267.67 | 11.00 | ||||
* t0.05=2.776; t0.01=4.604. ν=4; by Student’s T Test; if |t| < t0.05 or t0.01, then P > 0.05 or 0.01.The same below.
Table 4: Comparison between two potato mediums on the induction of conidiosporangium and sporulation.
II. Affect of growth on potato medium with different pH values: The spreading speed of the colony on PDA with different pH values showed a distinct phenotype. The spreading speed of the mycelia on media at pH 6.0 and pH 7.0 were faster, and the changing curve of the mycelia on media at pH 8.0 was the slowest and yielded unusual colony morphologies (Figure 10).

Figure 10: Colony diameters of the pathogenic strain ZZ35 on media at different pH values over a 7-d period.
Both the number of sporangia and the concentration of spores in the suspensions presented a downward trend as the pH value increased (Table 5). The more acidic medium (pH 5.0) contributed to the induction of sporangia and the generation of spores, whereas the more alkaline environment (pH 8.0) showed a relative inhibitory effect. There was no obvious diversity between the media at neutral pH values (pH 6.0 / 7.0).
| pH value | Number of conidia capsule per dish/a | Concentration of spore suspension/105·ml-1 | ||||
| 5.0 | 397.67 | a | A | 14.83 | a | A |
| 6.0 | 274.00 | b | AB | 12.33 | b | AB |
| 7.0 | 206.67 | b | BC | 11.17 | b | B |
| 8.0 | 113.67 | c | C | 1.33 | c | C |
Table 5: Affection of the pH value of potato medium on the induction of conidiosporangium and sporulation.
III. Influence of temperature conditions on in vitro cultures: The culture temperature had a strong impact on colony growth, sporangium induction and spore generation. At 30°C, the mycelia had a tendency to grow more vigorously, whereas the colony spreading was slower at 25°C. The slower tendency was even pronounced for the plates cultured at 15°C than for the two higher temperatures, in addition to irregular colony morphologies (Figure 11). The pathogenic colonies failed to grow at 5°C and showed no noticeable changes (not shown in graph).
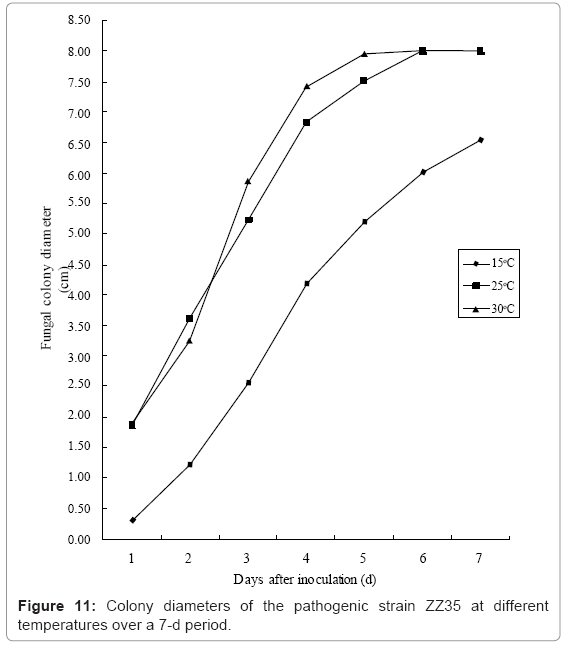
Figure 11: Colony diameters of the pathogenic strain ZZ35 at different temperatures over a 7-d period.
In the treatment at 25°C, the number of sporangia and the concentration of the spores in the suspensions were both at their highest level. When the environmental temperatures were too low (≤ 15°C), no conidiospores could be observed via microscopy (Table 6).
| Temperature | Number of conidia capsule per dish/a | Concentration of spore suspension/105·ml-1 | ||||
| 5°C | 0.00 | - | - | 0.00 | ||
| 15°C | 4.33 | c | B | 0.00 | ||
| 25°C | 240.33 | a | A | 10.33 | t=5.0142 | |
| 30°C | 172.00 | b | A | 6.50 | ||
Table 6: Influence of temperature conditions in vitro culture on the induction of conidiosporangium and sporulation.
IV. Impact of lighting sources and lighting conditions: Under the light provided by the fluorescent tube for 24 h/d, the spreading speed of the mycelium was faster than other treatments (Figure 12); however, there were fewer conidiosporangia which had low maturity levels, so we could not observe any spores (Table 7). The constant ultraviolet light (λ=360-370 nm) from the black light lamp contributed to conidiosporangia induction and maturation. Under the 12 h/d alternating light of the fluorescent tube and black light lamp, the levels of both colony growth and conidia production were higher than those of the 24 h/d black light treatment. The mycelia colonies grew more slowly and anomalously in total darkness, and did not generate any conidiosporangia.
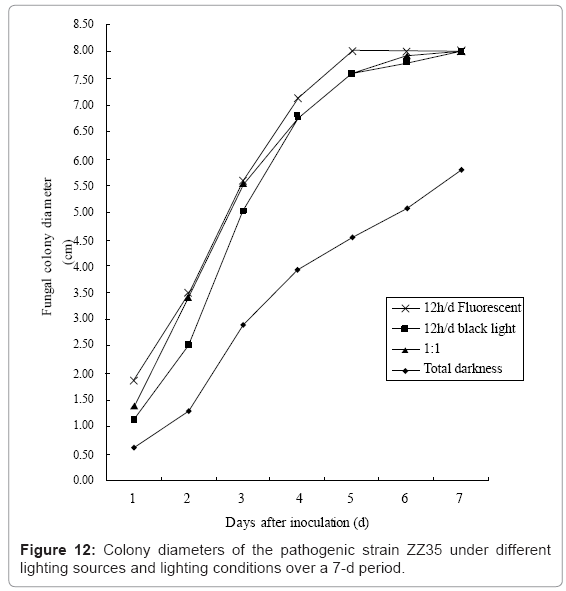
Figure 12: Colony diameters of the pathogenic strain ZZ35 under different lighting sources and lighting conditions over a 7-d period.
| Light source / Light conditions | Number of conidia capsule per dish/a | Concentration of spore suspension/105·ml-1 | ||||
| 24h/dDarkness | 12.00 | d | C | 0.00 | ||
| 24h/d Fluorescent lamp | 108.67 | c | B | 0.00 | ||
| 24h/d Black light | 265.67 | b | A | 11.67 | t=4.4721 | |
| (12h Black light / 12h Fluorescent lamp)/d |
356.33 | a | A | 18.33 | ||
Table 7: Impact of lighting sources and conditions on the induction of conidiosporangium and sporulation.
V. Effect of scraping treatment on the production of conidiosporangia and spores: At six days post-inoculation, the whole Petri dish could be covered by mycelium. If the cultures were maintained, the vigorous growth of the hypha might inhibit sporangium induction or spore generation. An increase in the number of sporangia and spores might appear after scraping the fungi mycelium. There were very significant differences in the number and concentration of sporangia in the suspension between the two treatments as determined by Student’s t-test (Table 8).
| Treatment | Number of conidia capsule per dish/a | Concentration of spore suspension/105·ml-1 | ||||
| Not strike off | 25.33 | t=11.2204 | 0.67 | t=10.5877 | ||
| Strike off | 230.67 | 17.50 | ||||
Table 8: Effect of scraping treatment on the induction of conidiosporangium and sporulation.
In traditional microbial taxonomy, in which fungi classification is determined via morphological, physiological, and biochemical characteristics or the antigen structural characteristics of the pathogen, false positive or negative results may have appeared that have effected fungal classification and determination. With the rapid development of molecule biology, PCR amplification and sequence analysis of the ITS ribosomal regions of the pathogen have been considered effective methods for fungal bioassays and disease diagnoses.
There is a heterogeneous homology for 5.8, 18 and 28S rRNA sequences, which are highly conserved in rDNA. There are few evolutionary pressures for natural selection on the ITS regions, which may have led to high mutation rate. Because there are extensive sequence polymorphisms in eukaryotes, sequence diversities can be observed in ITS regions of even two closely and genetically related species and may reveal the most recent evolutionary characteristics. Consistent with previous research, the evolutionary rate of the ITS region is ten times higher than that of the 18S rDNA. Therefore, the ITS sequence alignments can be used as a theoretical basis for microorganism population identification and community analysis [28]. For the moderately conserved ITS1 and ITS2 regions, sequence conservation is relatively uniform for intra-species specific and interspecies specific differences. These characteristics are beneficial when using molecular biology via ITS sequences to identify fungal species and determine the phylogenetic relationship between congeneric species or in communities with significant intra-species specific diversity.
In this study, we used rDNA-ITS sequence analysis to study ten strains of Physalospora piricola Nose, and observed two obvious sequence discrepancies in the ITS1 region; one was a one base-pair deletion and the other was a base-pair mutation (T/C). By comparing homologies to registered sequences in the NCBI database via BLASTn, we constructed a phylogenetic tree with Cluster 1.83 and MEGA 4.0 and determined that the ten strains were Physalospora piricola Nose, which belongs to Botryosphaeria dothidea which can be divided into two sub-varieties; the first sub-variety mostly originates from China, whereas the second originates mostly from Europe. The phylogenetic distance of these pathogenic fungi is different for different regions. The ten tested strains from Malus pumila Mill all clustered with the first sub-variety; however, the only strain from the Liaoning province (LW48) was phylogenetically distinct from the other strains from the Henan province.
Although we collected two samples from same district and from tissues with identical symptoms, these two strains (ZZ23 and ZZ25) had different ITS sequences, which might correlate with resistance to previously used pesticides. There was a base mutation in 28S rRNA for ZZ23, when ZZ25 preformed tow same base mutations in ITS1 region with ZZ35 and LW48. However, there was one more base mutation for LW48 in ITS1 region, which was obtained from Liaoning province. Through the resistance comparison of different varieties of apple germplasm to Physalospora piricola [29], ‘Fuji’ and ‘Zaohong’ were considered as high susceptible varieties, while ‘Tengmu’, ‘Redchief’, ‘Golden Delicious’ and ‘Ralls Genet’ as resistant cultivars relatively. Combined with the strains mutations, there may be some correlation between the disease resistance and base mutations. Strains from susceptible varieties ‘Fuji’ and ‘Zaohong’ showed no mutation (ZZ01, ZZ12, ZZ13). And for most stains from more resistant cultivars, many base mutations could be observed, however, there was no significant correlation between the mutagenesis and disease tissues or pathological lesions. Based on these results, for the pathogenic strains from same district, variety and from tissues with identical symptom, the sequences of ITS region can display genetic variation, and for strains from different districts, varieties or tissues, the ITS sequences may also display mutations.
From the comparisons of biological characteristics and pathogenicity of different strains, we could infer that the strains with more base-pair mutations had stronger growth rates, more spore production and increased infection compared to the strains with less or no variation. The biological characteristics of the three strains with variable ITS1 region sequences were significantly better than the other strains, as were their pathogenic capabilities, including LW48, which had one more base-pair mutation in the same region and induced the most spores and had the strongest infectious capabilities. Compared to ZZ25, which had a similar origin, ZZ23 had a slower hyphae spreading speed and decreased pathogenicity. This strain did not have a mutation in its ITS1 region, which might suggest that this mutation in the ITS1 region correlates with certain features including pathogenicity in a variety of strains. The differences in biological characteristics and pathogenicity between ZZ23 and ZZ55 were obviously different and correlated with a variety base-pair mutations, with no mutation in ITS1 region. Thus, we can preliminarily conclude from our results that there are certain relationships between the level of mutation and the biological characteristics and pathogenic capabilities of a strain. The base-pair mutations in the ITS1 region may have increased both the growth parameters and disease-causing capabilities, which might have a positive correlation with the mutational level of the entire sequence. Moreover, these experimental results suggest that there should have been positive correlations between the growth rate of the fungal colonies and spore production abilities and the pathogenicity toward the fruit. From the correlation between the mutagenesis and variety resistance, it could be inferred that there might be more mutational probabilities to the strains obtained from resistant cultivars. It could also suggest that mutation stains have powerful pathogenicity against the resistance. But for the high susceptible varieties it was easy to get disease, so the mutational level was lower for the strains to infect the cultivars with less pathogenicity. However, further research with more strains is needed to investigate the specific relationship between these characteristic variations for different strains and base-pair mutations in the ITS regions in the rDNA sequence, and the interrelationships between different mutations.
Based on our comprehensive analysis of the cultivation results for the pathogenic colonies of Physalospora piricola Nose, the appropriate conditions for in vitro growth include a temperature of 25-30°C and cultured on dishes with a neutral or slightly acidic PDA or PSA medium (pH 5.0 - 7.0).
Similar to the relationship between the vegetative growth and the reproductive growth of plants, the correlation between mycelial growth and conidiosporangium induction and spore production has been investigated. A constant temperature of 25°C was considered to be most beneficial for sporangium development and spore production. A possible explanation for the lack of conidiosporangium induction at low temperatures (≤ 15°C) may be that the mycelium growth was too poor to provide adequate nutrition for conidiosporangium induction, whereas when the culture temperature was relatively high (≥ 30°C), it was difficult for the colonies to transition to the reproductive development phase from the overgrowth of mycelium. In the pH 6.0 medium, the fungal hyphae grow better, whereas more conidiospores were induced in this partially acidic environment (pH 5.0). There was no significant variation between PDA and PSA media for the analysed biological characteristics.
We observed that the lighting sources and lighting conditions had decisive effects on the induction and development of conidiosporangia and spores. Continuous irradiation with short-wave black light (λ=360-370 nm) effectively induced the conidiosporangia and spore production. Under the 12 h/d alternate irradiation of black light and fluorescent light, the mycelium may have been growing vigorously for the first few days, whereas both the sporangia and spores could be effectively induced at a later time. Conversely, under the 24 h / d lighting with a fluorescent lamp, the colonies showed a fast hyphal spread, whereas the development of sporangia or spores was poor. The mycelial colonies grew with irregular colony morphologies had the slowest growth rate in total darkness and no conidiosporangia found. We can therefore conclude that although the growth of the mycelia benefits from fluorescent irradiation, the adequate short wave light was necessary to induce conidiosporangia formation and subsequent massive spore production [30].
Removing the mycelia after their growth period had a significant effect in promoting the generation of conidiophores [31]. Moreover, the colonies that were not scraped had few conidiosporangia or spores, whereas excessive mycelia covered the entire dish. One plausible explanation for this effect could be that the scraping process inhibited the constant mycelial growth, which then promoted the reproductive development phase of the fungal pathogen.
In the process of natural selection and evolution, pathogens induce mutation to adapt themselves to variations in host plant species, environmental changes, and chemical prevention methods. Together, the development of molecular biotechnology and intensive study of disease pathogenesis via DNA isolation and ITS sequence analysis of pathogenic fungi have enabled phylogenetic classifications and investigations into extent of DNA mutation. From the conidiospore induction of Physalospora piricola Nose in vitro cultures and the examination of biological characteristics and the pathogenic capabilities of different strains, new prevention methods can be developed through regulating the types and doses of pesticides and improving of cultivation conditions, which may promote the control the occurrence or spread of disease and provide guidelines for industrialised apple production.
However, further research should be performed to determine the specific details of condition-related regulation processes for colony growth and the induction of conidiosporangia and spores for Physalospora piricola Nose.
This research was supported by a Grant-in-Aid for Scientific Research from the National High-Tech Research and Development Program (National ‘863’ Program, No. 2008AA10Z157) and Jiangsu Science and Technology Support Project (No. BE2008316). The pathogenic strains were acquired from the Zhengzhou Fruit Research Institute, Chinese Academy of Agriculture Sciences. One of the strains was delivered from Institute of Pomology, China Academy of Agricultural Science, Xingcheng, Liaoning.