Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Review Article - (2015) Volume 4, Issue 1
Keywords: Biofilm, Proteases, Staphylococci, Proteinase K, agr
It is generally accepted that in the natural environment, bacteria prefer to live in surface-associated communities because of the myriad advantages such kind of lifestyle offers. These complex sessile structures are widely known as slime layers or biofilms. In a biofilm structure, the bacteria are embedded in a self-secreted extracellular polymeric matrix, and are well protected against environmental stresses, antimicrobial agents and the host immune system. Since biofilm associated bacteria can be upto 1000 times more resistant to antimicrobial agents, they are the cause of a range of problems, including quality and safety issues in food and health industry, commercial problems in shipping and aquaculture industry etc. From a medical perspective, biofilms have been implicated in a variety of human infections, such as endocarditis, osteomyelitis, chronic otitis media, gastrointestinal ulcers, and urinary tract infections, chronic lung infections like cystic fibrosis patients and periodontitis [1,2]. Since bacteria can form biofilm on both abiotic and biotic surfaces, places like implants and medical devices e.g. catheters, mechanical heart valves, pacemakers, prosthetic joints, and contact lenses, as well as tooth surface and various host tissue surfaces are regions with high propensity of getting infected with biofilms [1,2]. Due to the recalcitrant nature of biofilms, infections involving them prove to be chronic in nature. Both Gram-negative and Gram-positive bacteria are capable of forming biofilms. The most common biofilm-forming Gram positive bacteria include members of the genera Enterococcus and Staphylococcus [1,3]. Among Staphylococcus species S. aureus and S. epidermidis are the ones most commonly associated with chronic infections [3,4].
Biofilm formation occurs through a series of steps which begins with initial attachment of planktonic bacteria to a solid surface that is present at the air-water/liquid interface. This step is followed by subsequent proliferation and accumulation of the cells in small multilayer cell clusters known as microcolonies. The microcolonies then further proliferate to form giant assemblages of cells enmeshed in an extracellular matrix, which covers entire surfaces, and protects its inhabitants from detrimental effects of all sorts [4-6]. A mature well established biofilm is not a static structure, rather it is highly dynamic in nature, where old cells are constantly being dispersed and new members being recruited for this surface-associated community to expand, at all times. The composition of the extracellular matrix is very difficult to ascertain and variable among different bacterial species and even within the same species under different environmental conditions [3]. Despite this fact exopolysaccharides are an essential component of virtually all biofilm structures, providing the necessary matrix in which the bacterial cells are initially embedded. Among the many different exopolysaccharides that are known β-1,6-linked N-acetyl glucosamine forms the most common component of the biofilm matrix of many different bacteria [3]. Besides exopolysaccharides, surface proteins and protienaceous components also play an important role in biofilm formation [7]. Most of the surface proteins involved in biofilm formation share several structural and functional features [7,8]. The first member of this group was described in a Staphylococcus aureus bovine mastitis isolate and was named Bap, for biofilm associated protein [1].
Proteases are a large class of enzymatic molecules that catalyze the cleavage of peptide bonds. They are present in all living organisms, in which they display many essential physiological functions ranging from generalized protein degradation to more specific regulatory activity [8]. Proteases can be both intracellular and extracellular in nature. The extracellular proteases are less selective in their substrate recognition and can cleave both self and non-self molecules with equal efficiency. It is therefore essential that these enzymes be expressed as zymogens or in their inactive forms so as to prevent premature proteolytic activity which is injurious to producer cell itself [8,9]. Extracellular proteases are usually activated in a complex cascade involving auto-processing and proteolytic maturation [8]. Initial belief that biofilm matrix is made up mostly of polysaccharides, has been questioned with recent findings that demonstrate that surface and secreted proteins and extracellular DNA (eDNA) are also important factors in biofilm formation, stability, and regulation [6,7]. In this light, the role of proteases becomes clearer and recent reports have shown that the application of various proteases to bacterial cultures have often resulted in the reduction of biofilm formation and in the dispersal of established biofilms [9,10]. Flemming and Wingender [11] have also reported that extracellular proteins play multiple roles in the biofilm, participating in structure and quorum-sensing functions and as extracellular enzymes operating within the matrix. In addition, it is also presumed that extracellular proteases may be involved in regulating biofilm detachment and dispersal.
The present review talks about extracellular protease mediated regulation of biofilm formation in Gram positive bacteria, specifically Staphylococcus aureus and use of extracellular proteases as a possible tool for disruption of bacterial biofilms.
Biofilm formation
Staphylococcus aureus is amongst the most versatile of the Gram positive human pathogens and a major cause of hospital-acquired and implant-associated infections. Due to its ability to elaborate a range of virulence factors S. aureus can establish itself in various tissues and surfaces of the human body. Infections caused by this organism can range from superficial lesions, such as wound infections and abscesses, to the more serious life-threatening conditions such as bacteraemia, endocarditis, meningitis and osteomyelitis [12]. Gene transcription and subsequent expression of virulence factors in Staphylococcus aureus is under strict control of two global regulatory elements, agr (accessory gene regulator) and sarA (Staphylococcal accessory regulator A) [13,14]. agr locus is a cell density dependent quorum sensing regulator which represses synthesis of cell-wall associated proteins in the post-exponential phase and activates expression of extracellular proteins [13]. The S. aureus accessory gene regulator (agr) locus, a quorum-sensing regulator, produces and responds to the QS signal molecule called an autoinducing peptide (AIP). AIP, is an eight-residue peptide with the last five residues constrained in a cyclic thiolactone ring [13]. During growth, AIP is synthesized and secreted outside the cell through a mechanism that is still not very well understood. Once the extracellular concentration of AIP reaches a critical concentration, it binds to a surface histidine kinase receptor, initiating a downstream signaling cascade that controls expression of a bunch of virulence factors, such as proteases, hemolysins, and toxins etc. A very interesting fact worth noting here is that high level of activity of the QS locus agr is responsible for bacterial dispersal from biofilms and biofilm detachment. A succinct explanation for this can be obtained from the fact that since the agr locus is responsible for down regulation of surface molecules, that serve as tools for initial attachment and subsequent biofilm formation, and in addition it also controls the expression of extracellular proteases that play a significant role in biofilm detachment from the substratum, cells expressing increased agr activity cannot form biofilm like structures. Cells of S. aureus that do form stable biofilms have negligible amounts of agr activity. sarA also regulates virulence determinant synthesis either directly or indirectly by the modulation of agr expression [15].
First step of biofilm formation is binding to surface. Staphylococci initially attach either to host tissue or to the surface of a medical device via interaction with surface proteins or molecules respectively [5,16]. The attached bacteria then proliferate and accumulate into multilayered three dimensional cell clusters, which are embedded in an amorphous extracellular polymeric material. A mature stable biofilm contains fine fluid-filled channels innervating the entire structure [3-5]. These channels ensure transport of nutrients and oxygen to inner regions of the biofilm architecture so that cells located deep within can obtain the material required for survival. As the biofilm structure ages small groups of cells from the peripheral region slowly detach and find newer sites for infection establishment [4]. The various molecular mechanisms underlying this process of biofilm dispersal is still unknown but self-produced extracellular proteases seem to play an important part [5,17].
Extracellular matrix composition of S. aureus biofilms
For a long time it was thought that the major structural component of S. aureus biofilm was an extracellular polysaccharide, also known as polysaccharide intercellular adhesion (PIA), which is the product of genes of the ica operon (ica operon consists of the following genes: icaA, icaD, icaB, and icaC) [1,5,11]. However, recent reports indicate that many strains with deletion in the ica operon did not lose their biofilm forming capability showing that PIA is unessential for biofilm formation [14]. It is now assumed, and slowly evidence is being accumulated, that proteins play an important role in biofilm formation in this Gram positive pathogen. S. aureus possesses numerous surface exposed MSCRAMMs (microbial surface components recognizing adhesive matrix molecules) as well as secreted proteins that contain binding domains for matrix proteins [1,5,18]. S. aureus surface proteins that facilitate biofilm formation include the S. aureus surface protein G (SasG) [5,19] and its homologue accumulation associated protein (Aap) in S. epidermidis [18]. These surface associated proteins can also help in biofilm establishment in a polysaccharide independent background. The biofilm-associated protein Bap mentioned previously is involved in S. aureus adherence to abiotic surface, intercellular adhesion, and biofilm accumulation [1,5,7,19]. The Bap-homologous protein Bhp may be involved in biofilm accumulation in S. epidermidis [7]. Some other Staphylococcal surface proteins or cell wall associated proteins that have been implicated in surface attachment/adhesion and possibly biofilm formation include: SasX and SasC (structurally uncharacterized), FnBPs (Fibronectin binding proteins), ClfB and Protein A [5]. Extracellular DNA (eDNA), released from cells with the help of aureolysin, a S. aureus protein, contributes to the structural stability of S. aureus biofilms [5,20].
Extracellular proteases of S. aureus
The proteases encoded in the S. aureus genome are organized into four distinct operons mediating synthesis of a metalloprotease (Aur), seven serine proteases (SspA and SplA-F) (organized in two distinct operons), and two cysteine proteases (Staphopains ScpA and SspB) (Figure 1) [20,21]. The six Spl enzymes coded by splA-F do not require proteolytic activation. The Aur, ScpA, SspA, and SspB proteases are produced as zymogens or pro-enzymes [12,21] (Figure 2). The Aur and ScpA zymogens undergo auto-proteolytic cleavage outside the cell [12] and SspA and SspB are activated in a subsequent proteolytic cascade which is initiated with cleavage of SspA by Aur which then activates SspB [21]. The secreted extracellular proteases have a broad substrate specificity hence can degrade both "self" and "non-self" proteins. Consequence of deletion of all the proteases produced by S. aureus is increased abundance of secreted and surface-associated virulence factors [21]. The proteases aid survival in a protein rich environment especially by helping in nutrient acquirement and by cleaving deleterious antimicrobial peptides.
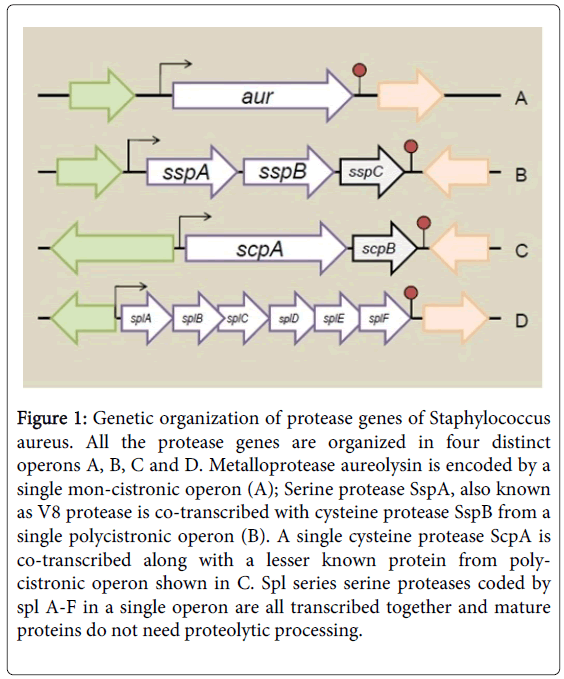
Figure 1: Genetic organization of protease genes of Staphylococcus aureus. All the protease genes are organized in four distinct operons A, B, C and D. Metalloprotease aureolysin is encoded by a single mon-cistronic operon (A); Serine protease SspA, also known as V8 protease is co-transcribed with cysteine protease SspB from a single polycistronic operon (B). A single cysteine protease ScpA is co-transcribed along with a lesser known protein from polycistronic operon shown in C. Spl series serine proteases coded by spl A-F in a single operon are all transcribed together and mature proteins do not need proteolytic processing.
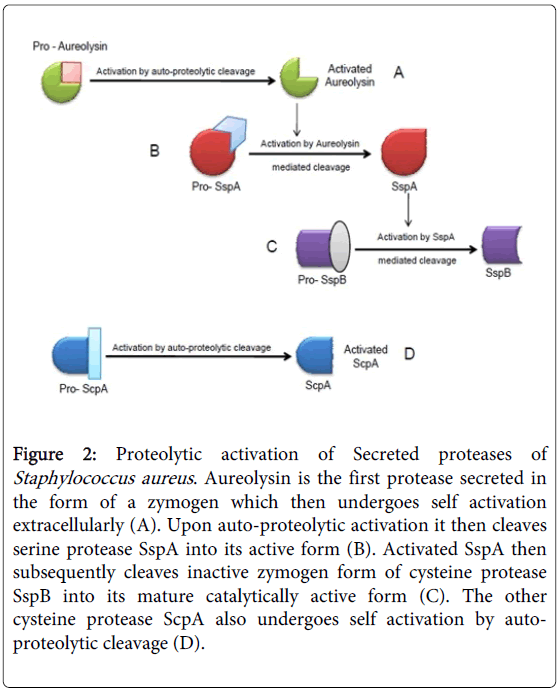
Figure 2: Proteolytic activation of Secreted proteases of Staphylococcus aureus. Aureolysin is the first protease secreted in the form of a zymogen which then undergoes self activation extracellularly (A). Upon auto-proteolytic activation it then cleaves serine protease SspA into its active form (B). Activated SspA then subsequently cleaves inactive zymogen form of cysteine protease SspB into its mature catalytically active form (C). The other cysteine protease ScpA also undergoes self activation by autoproteolytic cleavage (D).
Serine proteases
Serine proteases are enzymes that contain a histidine, serine, and/or an aspartate residue at their active site. The seven serine proteases secreted by S. aureus are all members of the trypsin family of enzymes [22]. The SspA and SspB protease are encoded by a single poly-cistronic operon wherein SspA is a serine protease and SspB is a cysteine protease. The six Spl proteases (SplABCDEF) are encoded by a separate polycistronic operon. SspA, also known as V8 protease, is a glutamyl endopeptidase that has the distinction of being the first purified and characterized proteolytic enzyme of S. aureus [12,21]. SspA has narrow substrate specificity and plays an important role in proteolytic activation of SspB. S. aureus Spl proteases are less studied and poorly characterized. Of all the six Spl proteins; SplA, SplB, and SplC have been structurally characterized and share homology to SspA [21,22].
Staphopains
Cysteine proteases are characterized by two nucleophilic cysteine residues joined together by a thiol bond. S. aureus secretes two cysteine proteases, ScpA and SspB, which have been termed Staphopain A and Staphopain B, respectively [21,23,24] Each Staphopain belongs to the papain protease family and the mature forms of the enzymes are approximately 20 kDa each in size and have almost similar three dimensional crystal structures [23,24].
Metalloprotease
Aureolysin (Aur) is the only metalloprotease secreted by Staphylococcus aureus, and it requires both zinc (Zn2+) and calcium (Ca2+) ions for its activity [12,21,22]. Aur is required to activate SspA by proteolytic cleavage and this initiates a proteolytic cascade of activation (Figure 2). Aur also cleaves surface-exposed clumping factor ClfB phenol-soluble modulins (PSMs), AgrD, some host proteins and the antimicrobial peptide LL-37 [21].
Extracellular protease-mediated biofilm disruption in S. aureus
The S. aureus biofilm matrix contains protein components that maintain biofilm integrity. As mentioned above, S. aureus secretes its own proteases and these enzymes have known to degrade self proteins, some of which target biofilm matrix constituents thus playing a role in biofilm regulation. There is preliminary evidence that the SspA (V8) serine protease might be important in biofilm remodeling [12,22,24] but the contribution of the other proteases is less clear. Mootz et al. [21] have shown that Cysteine proteases Staphopains also play a significant role in regulating biofilm formation. They have shown that Staphopains have deleterious effects on biofilm integrity. Mootz et al. [21] demonstrated that when S. aureus biofilms were treated with over-expressed and purified SspB or ScpA it eliminated biofilm formation itself and ScpA was also able to disperse already established biofilm. It was also observed that and the biofilm inhibitory properties of ScpA were conserved among different strains of Gram positive pathogen S. aureus (Figure 3) [21].
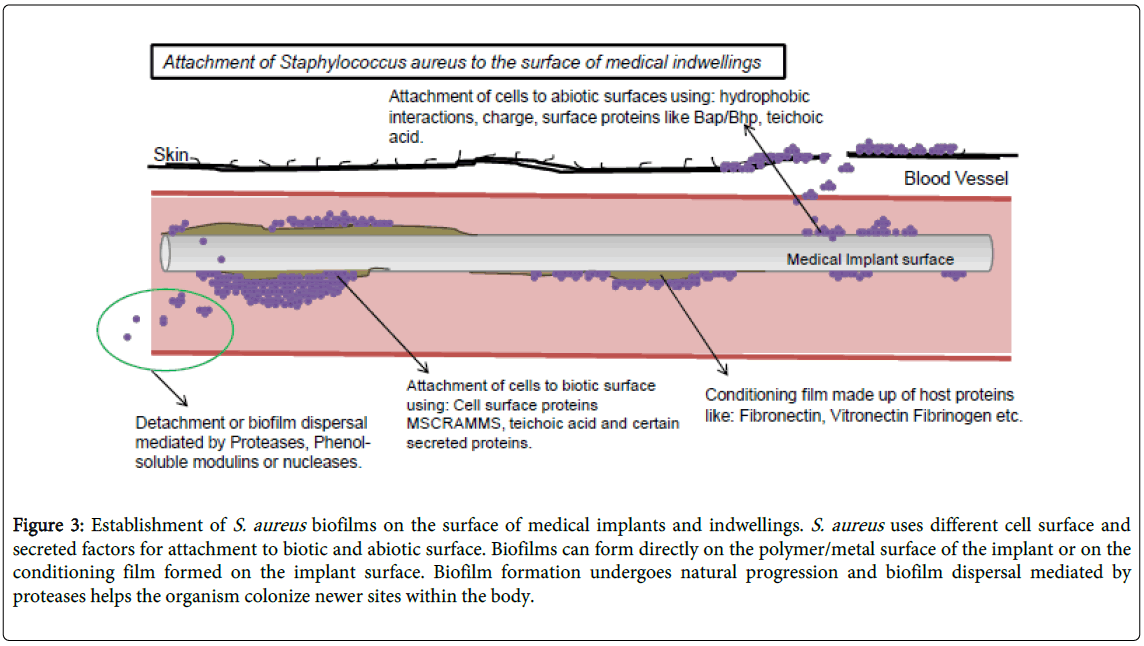
Figure 3: Establishment of S. aureus biofilms on the surface of medical implants and indwellings. S. aureus uses different cell surface and secreted factors for attachment to biotic and abiotic surface. Biofilms can form directly on the polymer/metal surface of the implant or on the conditioning film formed on the implant surface. Biofilm formation undergoes natural progression and biofilm dispersal mediated by proteases helps the organism colonize newer sites within the body.
The treatment of biofilms with proteases that have broad specificity, such as Proteinase K and Trypsin, leads to biofilm disassembly [13,21]. The serine proteases proteinase K (from Tritirachium album) and trypsin have frequently been used as efficient biofilm removal agents that hinder bacterial adherence and biofilm formation in S. aureus [25] presumably through degradation of surface structures [13,25,26]. Kumar et al. [27] also reported that protienaceous biofilms formed by S. aureus with the help of Bap proteins were susceptible to Proteinase K mediated detachment and dispersal. Biofilm assays done in 96 well microtitre plates showed that Proteinase K hampered both biofilm adhesion and development in Bap expressing S. aureus cultures.
Gilan and Sivan [25] have described a strain of Gram positive bacterium Rhodococcus ruber, R. ruber C208, which displays formation of highly hydrophobic and dense biofilm on the surface of polyethylene films while utilizing the polyethylene as carbon and energy sources. In their study they investigated the effects of several commercially available proteases on C208 biofilm formation and stability [25]. Serine protease trypsin significantly reduced biofilm formation by R. ruber C208, and the resultant biofilm appeared monolayered. However, proteinase K enhanced biofilm formation, the reasons for which can only be speculated.
Elchinger et al. [28] investigated the anti-biofilm activities of three commercial proteases, namely, Flavourzyme, Neutrase and Alcalase, against biofilm formed by two Staphylococcus species, S. aureus and S. epidermidis. Flavourzyme is an enzyme obtained from Aspergillus oryzae, which is actually an aminopeptidase [28]. Neutrase, from Bacillus amyloliquefaciens, exhibits endoprotease activity in neutral conditions, and its anti-biofilm activities have already been tested before against a range of bacteria [28]. Alcalase, isolated from Bacillus licheniformis, is a serine endopeptidase essentially composed of subtilisin A and it has recently been reported to dislodge multispecies biofilms in an industrial application [28]. Biofilm formation by the test organism was monitored using BioFilm Ring Test (BRT) which is designed to evaluate the kinetics of biofilm formation. No significant inhibition of S. epidermidis biofilm development was detected from 0 to 6 h, at all tested Neutrase concentration. However, an anti-biofilm effect was observed at Neutrase concentrations of 50 and 10 mU/ml after 24 h of incubation. So neutrase was found to effective only after proper biofilm establishment and its anti-biofilm activity was attributed to its endoprotease activity against proteins from the S. epidermidis biofilm. The effect on the S. aureus aureus biofilm was markedly different. A 72% inhibition of S. aureus subsp. aureus biofilm formation was observed after 4 h of incubation with 50 mU/ml of Neutrase and 47% inhibition with 10 mU/ml of Neutrase [28].
Similar experiment was conducted by Elchinger et al. with Alcalase. This endoproteolytic extract contains subtilisin, a serine endopeptidase with a broad spectrum of activity [28]. Subtilisins are known to be regularly used by bacteria in biofilm regulation due to their specific activity against adhesions [28]. However, Alcalase showed no anti-biofilm activity against S. aureus subsp. aureus biofilms and only mild inhibition of S. epidermidis biofilms.
A significant anti-biofilm effect of Flavourzyme against S. epidermidis biofilm was visible at 6 h which was maintained for 24 hours. The anti-biofilm action against biofilm formed by S. epidermidis was proportional to enzyme quantity and bacterial growth was unaffected by the presence of Flavourzyme. Flavourzyme was unable to disturb Staphylococcus aureus aureus biofilm formation at all used enzyme concentrations [28].
Artini et al. [29] reported the use of some commercially available proteases, which included: three serine proteases (proteinase K, PK; trypsin, TRY; and chymotrypsin, CHY) and two metallo-proteases (serratiopeptidase, SPEP; carboxypeptidase A, CpA) as tools for inhibiting biofilm formation by S. epidermidis and S. aureus both and human cell invasion by Staphylococcus aureus alone. In case of S. aureus, CHY and SPEP were reported to show significant inhibition capability (allowing only 36 and 59% of biofilm formation respectively) while PK had a weak inhibitory effect by allowing 69% biofilm formation [29]. Trypsin did not show any effect whereas CpA enhanced biofilm formation. In case of S. epidermidis, SPEP, PK and CHY showed an almost equivalent inhibitory activity by allowing 70–80% of biofilm formation [29], Trypsin again had no effect, while CpA favored biofilm formation tremendously. According to Artini et al. none of the proteases tested showed an effected the planktonic growth rate of the Staphylococcal species under study. It was concluded by the authors that among all the proteases tested, SPEP showed most potential to be developed as a novel anti-virulence tool against Staphylococcus biofilm formation and disease progression [29].
The antifouling (AF) potential of the serine protease Esperase HPF (subtilisin) was evaluated by Hangler et al. [30] for its ability to prevent the biofilm formation of a four-species bacterial species. One Gram positive bacterium namely Microbacterium phyllosphaerae and three Gram negative bacterial strains namely, Shewanella japonica, Dokdonia donghaensis and Acinetobacter lwoffii were all isolated from green alga Ulva australis. The effects of time and amount of Esperase HPF, was tested on the robustness and the oxidative metabolism of biofilm developed in 96 well plates [30]. Esperase HPF did not inhibit the oxidative metabolism of the bacterial biofilm or inhibit planktonic growth, but the enzyme inhibited biofilm formation by its proteolytic activity and heat inactivated enzyme had no effect. The enzyme concentration causing maximum biofilm inhibition in case of the different test organisms was determined after 72 hours [30].
Park et al. [31] identified some novel biofilm inhibitors of S. aureus, by screening culture supernatant of 458 actinomycete isolates. The culture supernatants in the ratio of 1% (v/v) of more than 10 Actinomycete isolates inhibited S. aureus biofilm formation by more than 80% without inhibiting growth of the test culture. Of the 10 positive isolates, nine belonged to genus Streptomyces and one belonged to Kribbella genus. For further study, two of the most potent cultures whose supernatants gave best anti-biofilm activity were chosen and were named as Streptomyces sp. BFI 250 and the strain Kribbella sp. BFI 1562 [31]. The culture supernatants of these biofilm reducing actinomycetes were found to contain a protease which in addition to inhibiting biofilm formation itself also disrupted pre-established biofilm structures. This study by Park et al. [31] suggests that protease treatment could be a feasible tool to reduce or eradicate biofilm mediated persistent S. aureus infections [31].
Iwase et al. and Sugimoto et al. [32-34] have shown that Esp, a serine protease form Staphylococcus epidermidis, disassembles already formed biofilms of S. aureus and inhibits its establishment and pathogenesis. Esp possibly degrades surface structures that help in providing the cohesive strength of S. aureus biofilms. Similar results were also shown by Vandecandelaere et al. [35] where authors explained the role of S. epidermidis extracellular protease in inhibition of S. aureus biofilm formation both in vivo and in vitro. They have also demonstrated that although Esp protease degraded S. aureus biofilm matrix components, it did not affect oxidative metabolism of individual cells. Staphylococcus aureus biofilms treated with the protease containing supernatant from S. epidermidis were thinner and had almost no extracellular matrix. Also an increased survival of Caenorhabditis elegans was reported, when infected with S. aureus strains in the presence of S. epidermidis protease positive supernatant [35].
Park et al. [36] also tested the supernatants of 28 bacteria to identify new anti-biofilm molecules against S. aureus. The culture supernatant (1% v/v) of Pseudomonas aeruginosa PAO1 inhibited biofilm formation by the test organism by more than 90% without affecting its planktonic cell growth. The P. aeruginosa supernatant containing protease activity inhibited both biofilm formation and detached existing biofilms. Investigation using 13 protease-deficient P. aeruginosa mutants by Park et al. helped showed that LasB elastase was the enzyme responsible for the anti-biofilm activity [36].
Lysostaphin an endo-peptidase produced by another species of Staphylococcus Staphylococcus simulans, has also been reported to show potent biofilm disrupting potential [37,38]. Biofilms formed by both Staphylococcus aureus and Staphylococccus epidermidis are susceptible to Lysostaphin mediated biofilm dispersal [37,38].
Papa et al. [39] as well reported that endopeptidase from Serratia marcescens, also known as serratiopeptidase, has the capability to disassemble S. aureus biofilms by cleaving the protienaceous determinants proving structure and stability to the biofilms. The article claims that serratiopeptidase has potential to be used as an anti-infective agent since it inhibits Staphylococcus biofilm formation on the surface of implants and host tissues thus regulating biofilm associated virulence of the said pathogen.
Monnappa et al. [40] describe the anti-biofilm/anti-virulence activity of extracellular proteases produced by a host-independent Bdellovibrio bacteriovorus (HIB) for the first time. Bdellovibrio sp. is a Gram negative obligate aerobic bacteria which feeds on other Gram negative bacteria by invading their periplasmic space and surviving on the host bacterial biomolecules like proteins and nucleic acid. To sustain this kind of lifestyle Bdellovibrio genus is known to produce a large number of proteolytic enzymes and nucleases. Monnappa et al. report that addition of 10% of HIB supernatant resulted in 75% or more reduction in S. aureus biofilm formation as well as dispersal of pre-formed biofilms [40]. LC-MS-MS analyses revealed that host independent B. bacteriovorus supernatant contained various proteases, including two significant serine proteases that were named as Bd2269 and Bd2321 [40]. Authors carried out experiments with various protease inhibitors and significant inhibitory effect of AEBSF on the proteolytic activity of the HIB supernatant confirmed that serine proteases were present in the supernatant and that they were the ones responsible for S. aureus biofilm disruption [40]. The HIB supernatant was also to possess minor DNAse activity which could have also effected S. aureus biofilm establishment. Monappa et al. also report that treatment of S. aureus with HIB supernatant weakened its ability to invade human epithelial cells. Authors conclude that B. bacteriovorus hydrolytic enzymes, especially proteases, can adversely affect virulence of Gram-positive bacterial pathogens [40].
Recently, major advances have been made in elucidating the different structural components of the biofilm matrix, and identifying molecules involved in maintain biofilm integrity and robustness. The initial belief that, almost all microbial biofilms have polysaccharides as the major ‘sticking’ substance has been altered, with increasing evidence pointing to the role of proteins as crucial biofilm matrix components. This is especially true in case of Gram positive bacteria. One of the most well studied Gram positive pathogen S. aureus shows formation of biofilms in which proteins form a major component of ECM in addition to eDNA and other host factors. This finding highlights the role proteases as enzymes with potential to disassemble already well established biofilms or prevent initial attachment itself that is essential for biofilm formation [41,42]. This review highlighted the role of self-produced extracellular proteases of S. aureus in biofilm dispersal as well as role of commercially available broad spectrum proteases in biofilm disruption of other Gram positive bacterium. With increasing incidences of biofilms associated infections and fouling associated economic debacles, looking for newer and sustainable anti-biofilm strategies are the need of the hour. Extracellular Proteases may prove to be the solution for this problem as they bring with them the advantage of being easy to apply and easy to produce in bulk for large scale application.
RM and AP wish to thank University Grants Commission for PhD fellowship.