Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2017) Volume 6, Issue 1
Ribonucleases (RNases) are small molecules which are highly cytotoxic in nature. They catalyse the degradation of RNA into smaller molecules rapidly in an unprotected environment. The cytotoxic properties of RNases include degradation of RNA leading to blockage of protein synthesis in malignant cells and inducing the apoptosis response. Cytotoxicity of RNases is determined by catalytic activity, stability, non-selective nature of inhibitors, positive charge on molecule and internalization. Onconase, BS-RNase and other RNases exert cytotoxic activity on cancer cells selectively by involving different cellular pathways and/ or enhance the cytotoxicity by mutation. A general mechanism of the cytotoxic activity of RNases includes the interaction of the enzyme with the cellular membrane by non-specific interactions mediated by Coulombic forces, internalization by endocytosis, translocation to the cytosol, degradation of ribonucleic acid and subsequent cell death by activation of caspase-dependent mechanisms, low molecular weight compounds or alteration in protein and NF-κB signal pathway. But still it is unclear that which of these mechanisms is most potent, common and causes cell death in cancerous cells. The problem related to ribonuclease inhibitor (RI) has not fully elucidated. This article looks at the cellular pathways of RNases and mechanism of their cytotoxicity towards the malignant cells which makes RNase as a strong candidate to be considered as chemotherapeutic or antitumor drug. Some of the prominent approaches to exploit RNase molecule as an anticancer therapeutic agent have been discussed.
Keywords: Ribonuclease, Cytotoxicity, Cellular pathways, Apoptosis, Anticancer therapeutics
In recent years, the use of RNase as non-mutagenic anti-cancer drugs has attracted the attention of many researchers. In human body, almost 1000 distinct type of cancer and subtype of tumour can be found within specific organs. RNases are a large group of hydrolytic enzymes that catalyze the degradation of ribonucleic acid (RNA) into smaller components with a very short lifespan in an endangered environment. RNases are a super family of enzymes which catalyze the degradation of RNA by operating at the level of transcription, protein synthesis and thus play key roles in the regulation of vital processes in any organism ranging from virus to human [1]. It is a type of nuclease which has the ability to catalyze cleavages of phosphodiester bonds in cellular RNA and also possess the biological activities like the maturation of mRNA and non-coding RNA, RNA interference, apoptosis induction and protection against viral infections these all process were not possible without RNA degradation [1,2]. RNases are heterogeneous groups of enzymes that are classified according to their RNA processing mechanisms: exoribonuclease and endoribonuclease which are ubiquitously present in prokaryotic and eukaryotic micro-organisms [2,3]. Exoribonuclease enzymes cleave RNA molecules from either the 3' or 5' terminus, and endoribonucleases cleave single-stranded RNA (ssRNA) molecules internally at the 3' end of pyrimidine residues into 3’-phosphorylated mononucleotides and oligonucleotides. Exoribonuclease and endoribonucleases are further comprising several sub-classes. Major types of endoribonuclease are RNase II, PNPase and oligoribonuclease for RNA degradation, and in the case of exoribonuclease such as RNase PH, RNase BN, RNase D and RNase T are responsible for the 3' end maturation of RNA [4]. Some RNases have potential impact on malignant cells. Some of the ribonucleases possess higher molecular heterogeneity, with related protein that reveals high specificity for cleavage of specific loci in RNA [1,5]. RNases are omnipresent, with a very short lifespan in an unprotected environment. Microbial and mammalians ribonuclease are the best characterized enzymes. In 1957, Aspergillus orayze was identified as the first source for RNase production. BaRNase (RNase Ba) has also been identified and purified from first prokaryotic organism (Bacillus amyloliquefacians). RanpiRNase known as Onconase and RNase A as bovine pancreatic RNase are largely produced from animal sources [1,6-8].
RNases also have many potential applications for sequence and structure analysis of RNA, oligonucleotides preparation, RNA removal from Single Cell Proteins (SCP) and intrinsic properties of RNase helps in the treatment of malignant cells. Natural RNases have potential abilities to inhibit animal tumors and viruses and hence are considered as alternative chemotherapeutic drugs. RNases have also been used to produce nucleotides for clinical applications commercially (Table 1) [9].
| RNase type | Source | Mode of action | Application | Reference(s) |
|---|---|---|---|---|
| BS-RNase | Bovine seminal fluid | In vitro, bind to cell membrane and disturb bilayer membrane and rnter to cytosol for degrade the natural RNA in cell | Thyroid cancer | [1,10-12] |
| Onconase | Oocytes and early embryo of leopard frog | Degradation of tRNA and inhibit the protein synthesis which leads to activation of caspases and induced apoptosis | Human lung cancer, pancreatic aden- carcinoma and mesothelioma cancer | [1,13-15] |
| RNase T1 | Fungal (Aspergillus oryzae) | Inhibition of protein synthesis and induced apoptosis | Under clinical trial | [1,2,15] |
| α-Sacrin | Aspergillus giganeus | Cleavage of phosphodiester bond of rRNA, block the protein synthesis | Under clinical trial | [1,16,17, 76] |
| RNase P | Human cell culture | Removal of 5’- leader sequence of tRNA, blockage of protein synthesis and apoptosis | Under clinical trial | [1,2,17] |
| RNAse T2 | Human cell culture | Blocking blood supply to tumours cell by stopping angiogenesis | Under clinical trial | [1,17,18] |
| Actibind | Aspergillus niger | Blocking blood supply to tumours cell by stopping angiogenesis | Under clinical trial | [1,17,18] |
| RNase Sa | Streptomyces aureofaciens | Internalized into cell by increasing net positive charge | Under clinical trial | [15,19,20] |
Table 1: Cytotoxic RNases and their mode of action.
In 1989, Sidney Altman and Thomas R Cech have been awarded a Noble prize in chemistry for their outstanding discovery of catalytic properties of bacterial RNA. RNases have revelation of their therapeutic potentials and ribozyme known as catalytic RNA helps to understanding the origin of life [10-15]. First ribozyme was identified as RNase P which helped to understand numerous cellular RNA metabolism and biochemical studies of the reactions catalyzed by the RNase alone and by the holoenzyme [16-20]. The goal of the present review is to summarize what is known about the microbial RNases mechanism of action and their therapeutics applications in medical field and industries [21].
RNases target the malignant cells for cytotoxic activity through the apoptotic response and thus the RNases are also considered as chemotherapeutic drugs. In previous studies, artificial RNases interaction with target cell, internalisation into cytosol of cancerous cell causing maximum toxicity effect was reported [15,22-24].
RNases have been produced from microbial cells, human and plants as extracellular or intracellular enzymes. These are best known for their ability to cleave RNA and also possess remarkable biological activities [24]. RNase A was the first enzyme to have its sequence determined and the third to have its structure revealed that prompted its used in many early protein-folding studies. In 1972, Stanford Moore, Christian Anfinsen and William Stein were awarded with Noble prize in chemistry together for their collective work on RNase A [25,26].
Different types of RNase molecules as anticancer agent
Bovine seminal ribonuclease (BR-RNase): BS-RNase is only one enzyme which occurs in quaternary structure and was discovered by Hosokawa and Irie, and Dostal and Matousek in 1972 [15]. BSRNase is a natural dimer with two identical subunits held together by two intramolecular disulfide bonds and non-covalent interactions which prevent the binding of RI due to steric reasons and accounts for its cytotoxic effect [1,27,28]. BS-RNase exists as dimmer forms and maintains the allosteric regulation by both substrate and product at nucleotide hydrolysis process. Single monomeric subunit of BS-RNase consists of 124 amino acid residues. BS-RNase is highly cytotoxic in dimmer structure and shows high activity. If, BS-RNase is reduced to monomeric structure then its cytotoxicity will decrease without affecting the ribonuclease activity. BS-RNase amino acid sequence and crystallographic structure are similar to the members of the RNase A superfamily. Lys41, His12 and His119 are catalytic residues of BSRNase and four disulfide bridges of RNase A are strictly conserved. The two consecutive Cys residues at position 31 and 32 are present in BSRNase and these form two disulfide bonds which cause dimerization. However, BS-RNase monomeric unit is highly catalytically active than the dimeric form of BS-RNase, but not cytotoxic [17,27,28]. The BSRNase monomeric unit is energetically inhibited by mammalian RI but does not interact with the natural dimeric enzyme. In general mechanism, BS-RNase cytotoxicity has been revealed by enzyme mixture of both M=M and M×M forms that indicated that both forms of BS-RNase equally enter the cancer cells by absorptive endocytosis. M=M form is dissociated into two monomeric subunits in the cytosol environment and inactivated by RI but M×M dimeric form appears to be stabilized by the non-covalent interactions which thus evades ribonuclease inhibitor and degrades intracellular or cellular RNA, thus leading to cytotoxicity. The analyses of single subunit variants of BS-RNase that evaded RI ribonuclease activity and cytotoxic activity were conducted. These monomeric variants had cytotoxic activity which was 30-fold higher than wild-type BS-RNase [27]. RI is a 50 kDa protein which is present in cytosol and forms a strong complex with mammalian RNases which eventually abolishes the RNase catalytic activity. Microbial and other RNases are unwilling to bind to mammalian RNase inhibitor. The RNase found in bull semen fluid shows antitumor, antispermatogenic and immunosuppressive activities due to its unique structure and enzymatic properties. It belongs to RNases group of enzymes endowed with Special Biological Actions (RISBASES) [17,28,29]. The cytotoxic effect of BS-RNase on tumor cells is accompanied by the induction of apoptosis. This has been proved by ultrastructure and flow cytometry evidence of apoptotic death following BS-RNase treatment in phytohemagglutinin-stimulated lymphocytes and normal cells [30]. Further, BS-RNase monomers are highly toxic to human tumor cells than other variant or homologue of RNase A, and amphibian homologue are already in phase III clinical trials for the treatment of unrespectable malignant mesothelioma. BS-RNase cytotoxicity properties can evident itself as an antitumor, embryo toxic, immunosuppressive and aspermatogenic activity which has the potential therapeutic value. BS-RNase has been found to be the promising tool for the treatment of thyroid cancer [31].
Onconase: Onconase belongs to the family of RNases A and it was isolated from early embryos and oocytes of Northern leopard frog (Rana pipiens). Onconase shows strong cytotoxic and cytostatic activity and has 30% sequence similarity with superfamily RNase A [1,17]. Onconase enzyme has the potential to degrade RNA and their substrate and also possesses highly cytotoxic effect for several cancer cell lines. Onconase also named as Ranpiranse is the first therapeutic product studied by Alfacell Corporation [1]. Its single chain consists of 104 amino acid residues which are 20 residues less than RNase A. Ranpiranse shows cytotoxic effect in vivo and in vitro in tumor cells. Ranpiransemediated cytotoxic activity observed by targeting the cellular tRNA and also breakdown the double-stranded RNA (dsRNA) at 40°C under same conditions. Currently, new mechanism followed by Ranpiranse activity for inducing cell death/ apoptosis in malignant phenotypes via the RNA interference mechanism involving miRNA and siRNA [1,32]. Onconase is the smallest enzyme in the RNase A superfamily and also cited as the first RNase tested in clinical trial in US and Europe. This has been involved in recent phase III trial for treatment of malignant mesothaelioma. Due to its cytotoxic and cytostatic properties, it has been used externally against the skin cancer [1,15].
In Onconase, Thr and Ser are present in 70:30 ratio of its molecule and both forms are equivalent with respect to their catalytic activity and cytotoxicity [15,33]. RNase A catalytic residues are conserved in all variants of Onconase (His10, Lys31 and His97). According to amino acid sequence and crystallographic studies it has been revealed that three disulfide bonds of RNase A are conserved in all variant of Onconase at position 19-68, 30-75 and 48-90. These catalytic residues and disulfide bond position lead to the prerequisite for cytotoxicity of RNases. Onconase is an exceptionally stable enzyme at temperature 90°C or at 4.4 M, as respective values for RNase A have been found to be 62.4°C and 2.8 M [15,34]. Onconase is very much conformationally stable and its unwilling leads to proteolysis [35]. The unpredictable thermodynamic stability of Onconase appears to be due to the presence of the C-terminal disulfide bond and N-terminal network of hydrogen bonds which interact with hydrophobic cluster [15,34]. Disulfide bonds substantially contribute to enzyme stability and also make Onconase more efficient than RNase A at recovering its catalytically and biologically active structure. The cytotoxicity of Onconase is same as BS-RNase because of catalysis of phosphodiester cleavage in mammalian cells. But Onconase practically does not interact with mammalian RI and its catalytic and antitumour activities are also terminated by alkylation of His residue. Onconase evades RI in cell cytosol because it lacks an amino acid sequence interaction with the RI. Onconase may possibly binds to cell surface receptor like a polypeptide or hormone or moves inside the target tumour cell by endocytosis. After entering into cell cytosol, it cleavages tRNA and inhibits the transcription process [17]. Onconase doesn’t follow specificity when it is degrading natural substrates [36]. The tRNA was reported as primary target for Onconase action in the tumour cells. Onconase degrades natural tRNA with different base specificity than synthetic substrates. Natural tRNA cleavage happens between two guanine bases, which are present in D-arm. Onconase always recognizes the native structural point for preferential cleavages. In synthetic substrate, Onconase has not been found to degrade GG bonds [13,36-37]. Protein synthesis is predominantly abolished by inhibition of transcription which eventually suppresses the inhibitors of apoptosis (IAP). IAP inhibition would lead to activation of the caspases that thus activates the apoptosis pathway(s) [17]. Onconase is internalized into cells due to surface exposed glycoproteins or hormones or endocytosis, where it causes cytotoxic effect and also it has also been found to reduce tumour size in animal model thus enhancing the cytotoxic efficacy in therapeutics process. It also displays nephrotoxicity, aspermatogenic, embryotoxic and immunosuppressive activity. Onconase has reached phase III clinical trials in US and Europe as an antitumor drug for the treatment of unresectable malignant mesothelioma [14,29].
RNase T1: RNase T1 is type of an endonuclease, which degrades phosphodiester bonds of single stranded RNA (ssRNA) between 3’-guanylic residues and 5’OH residues of adjoining nucleotides with the development of consequent intermediate 2’, 3’- cyclic phosphates.RNase T1 has been isolated from fungal sources, mainly Aspergillus and Penicillium species and it is known as Ribotoxin because of its cytotoxic character [1]. RNase T1 reveals its activity without metal ions and has been used to analyze RNA structure, mapping, RNA protection, and removal of RNA form extracted DNA samples. Neither RNase T1 from Aspergillus oryzae nor RNases U1 from Ustilago sphaerogena was inactivated by RI [25,17]. RNase T1 shows toxic effects by cleaving single phosphodiester bond located within conserved sequence of the large rRNA gene, known as the Sarcin-ricin loop and thus leads to inhibition of protein biosynthesis followed by cellular death by apoptosis. But if somehow RNase T1 is introduced into cells, they can produce Ribotoxin protein which is cytotoxic to tumor cells. Ribotoxins preferentially kill the tumour cells by altered membrane permeability if no specific protein receptor has been present in cell membrane, such as the cells infected with virus or transformed cells [1,15].
RNase T1 is composed of small protein α + β dimer containing 104 amino acid residues and contains four antiparallel beta pleated sheets which cover nearly five turnovers of a long alpha-helix. Cys2- Cys10 and Cys6-Cys103 disulfide bonds help in folding and unfolding of RNase T1 [38]. RNase T1 specifically cleaves the guanine bases for RNA denaturing. Onconase and α-Sarcin structure is related to that of RNase T1 and both are also toxic to tumor cells. Even if, RNase was unable to internalize into tumour cells thus leading to non-cytotoxic behaviour but it can still be manipulated to develop an antitumor drug. Researchers have found that if RNase T1 is incorporated into Hemagglutinating Virus of Japan (HVJ) envelope, this vector can be targeted to tumour cells and to make it to act as a unique anticancer drug. RNase T1 has been internalized into human tumor cells via a novel gene transfer reagent, HVJ envelope vector that resulted into tumour cell death. RNase T1 showed tremendously improved in cytotoxic activity because of the pre-treatment of HVJ envelope vector with protamine sulphate, which is in phase III human clinical trials as a nonmutagenic cancer chemotherapeutic agent [39]. Furthermore, internalized RNase T1 like an onconase or BS-RNase also induces the apoptotic cell death programs in the tumour cells. However, according to previous studies, RNase T1 cytotoxicity is unfortunately not specific to tumor cells, but it could be achieved by HVJ envelope vector thus making the RNase a specific antitumour tool [40,41].
α-Sarcin: Microorganisms have the greatest diversity on earth as they have existence in extreme environments. To encourage their own growth and also to promote colonization they produce extracellular RNases often referred as ‘ribotoxins’ to inhibit the growth of other organisms in the same niche that lessens the use of available nutrients by such organisms. These ribotoxins show specificity for binding on to the conserved domains of RNA in sarcin-ricin loop (SRL) part of ribosome(s). Some fungi such as Aspergillus niger, the black mold produces a ribotoxin known as α-sarcin which is a 150-residues polypeptide toxin, while mitogillin and restrictocin are small (approximately 17 kDa) ribosome-inactivating proteins (RIPs) produced by the Aspergillus giganteus. The α-Sacrin belongs to type 1 group of RIPs and are RIPs are predominant members of the fungal ribotoxins that display 3-D structure. These ribotoxins have been studied in greater details since early 1960s. The α-sarcin translocates mainly into the cytosol of a cell to direct its toxic effect. Inside the cells, it acts on ribosomal RNA (rRNA) associated with the ribosome. Such an action allows the extracellular RNases to be considered as RIPs. The RIPs and ribotoxins both specifically target SRL. The attachment of RIPs to the SRL causes either depurination of a single nucleotide or cleavage of a phosphate bond thus rendering the RNA and its subsequent function altered/ inhibited [17]. Such a function allows the use of α-Sarcin or RIPs as anti-tumor and anti-cancer molecules. The catalytic activity of α-Sarcin is shown by His50, Glu96 and His137 residues [42,43]. The fungal ribotoxins act on specific RNA to accomplish the cleavage of phosphodiester bonds in the universally conserved α-Sarcin domain of 28S rRNA and thus inhibit the protein synthesis [17]. α-Sarcin interacts with lipid bilayer of cell membrane, fuses with the membrane to permit its uptake and subsequent onset of inhibition of protein synthesis and/ or induction of apoptosis [43,44]. α-Sarcin has been previously reported as a potent cytotoxin that promotes apoptosis in human rhabdomyosarcoma cells [30]. Molecularly the ribotoxin belongs to superfamily of RNases. Analysis of the Mitogillin gene and PCRmediated site-specific mutagenesis suggested that positive domains in Ribotoxins, which share homologies with motifs in ribosome-related proteins and liable to be targeting the ribotoxins to the ribosome. These ribotoxins/ RIPs or α-Sacrin contains highly positively charged longer loops which account for their cytotoxicity towards tumour cells. Interestingly, the α-Sacrin inactivates the ribosome in cell-free systems and selectively intact with the cells to penetrate the cell membrane without involvement of permeable agent(s) in transformed or virusesinfected mammalian cells [45]. Due to this anti-tumor/ anti-cancer cell inactivation, the ribotoxins are potent therapeutics to manage malignant cells [1,17,43]. Edible mushroom like as Pleurotus sajor-caju possesses an RNase which exerts antiproliferative action on leukemia and hepatoma, as well as antimitogenic effect on mouse spleen cells [1,46]. Other RIPs namely marmorin and hypsine extracted from H. marmoreus, lyophyllin from Lyophyllum shimeiji, velutin from F. velutipes, and pleuturegin from Pleurotus tuberregium have also been obtained from fungi [1,46].
ACTIBIND and RNase T2: ACTIBIND is an extracellular glycoprotein protein containing 32 and 36 kDa peptides also produced extra-cellularly by Aspergillus niger. It belongs to T2 RNase family and has the ability to bind to the ubiquitous actin protein. It basically interferes with intercellular actin network-structure. Actin protein is responsible for the vessel formation and their migration in normal as well as malignant cells. In previous finding, ACTIBIND has been shown to bind with actin protein as a receptor on cell surface, acts as inhibitor for cell extension, cell migration, metastatis as well as circulation of malignant cells in the blood [1,40,41,47]. In previous studies, ACTIBIND was reported to inhibit the development of xenograft tumour in rat and mouse models, human melanoma growth and palpable tumour [47]. RNase T2 just like ACTIBIND binds to actin on the malignant cells and discourages the angiogenesis and metastases in animals and humans [25]. The cytotoxic of the ACTIBIND increases tremendously when it is directly penetrated or injected inside the cell cytosol. This observation shows that ACTIBIND uptake by the malignant/ cancer cell(s) is a rate-limiting factor, and if internalization or intracellular concentration of ACTIBIND in the malignant cells is somehow increased, the greater propensity of anticancer effect could be achieved. ACTIBIND and human RNase T2 could be considered as a front-line therapy in the fight against cancers [1,17].
Ribonuclease P (RNase P): RNase P is an endoribonuclease that catalyses the t-RNA degradation at 5’ end into smaller components. RNase P enzyme is unique from other RNases because it universally composed of both protein and RNA. RNase P enzyme occurs in all three domains of microbes i.e., bacteria, archaea and eukarya [48]. The eukaryotic RNase P differs from other two domains of microbes due to its protein composition and RNA structure. In vivo, bacterial RNase P has been associated with single protein but eukaya and archaea holoenzymes are more complex because both of these contain at least 4 to 9 proteins, respectively [49]. In vitro, human RNase P contains two proteins with RNA sufficient activity [50]. RNase P is a divalent cation-dependent endoribonuclease, which acts as riboenzyme and is encoded by protein subunit rnpA gene and RNA subunit rnpB [2,50]. RNase P functions to breakdown the precursor sequence of RNA on RNA molecules. Ongoing cancer therapy approaches have a major problem to differentiate between the cancer cells and the normal cells. This problem may be overcome by specific chimeric molecules, which are specific to the cancer cells, and thus efficiently act on the specific targets. RNase P has catalytic subunit M1 which catalyses the hydrolytic removal of 5´-leader sequence of t-RNA and also permits the strategy of gene targeting. M1 RNA subunit can be targeted to the mRNA by the addition of guide sequence at the 3´-terminal (M1-GS). M1 RNA becomes M1-GS, which cleaves the mRNA and will thus halt formation of the fusion proteins, which are specific for the cancer cells. M1-GS has been used as a therapeutic tool against BCR-ABL oncogene model [1,51]. BCR-ABL oncogene was created by the translocation of the sequences from ABL gene on CHR 9 to the BCR gene on CHR 22 [52]. BCR-ABL p190 and BCR-ABL p210 were created and both of these differed in BCR nucleotide but possessed identical ABLderived sequences. BCR-ABL p190 and BCR-ABL p210 were chimeric molecules which were highly specific to the cancer cells and served as the excellent targets. BCR-ABL p190 and BCR-ABL p210 oncogenes are responsible for the myelogenous and acute lymphoblastic leukaemia. BCR-ABL oncogenes inhibit the apoptosis by Bcl-2 pathway, inhibition expression leads to reverse phenotype and the cells die by apoptosis. In animal models, BCR-ABL model has not been evaluated the evaluation of efficacy of agent and delivery process is still a major problem. BCRABL system provides a selective, non-toxic and new therapeutic tool for cancer treatment in future. M1-GS should target only at the junction sequences of the transcribed mRNA and selectively damage the cancer cells. M1-GS gene therapy promises to be an effective strategy for the future treatment of the cancer [1,17,53].
RNase Sa: Streptomyces RNases Sa (strain BMK), Sa2 (strain R8/26), and Sa3 (strain CCM 3239) are microbial RNases belonging to the RNase T1 family. These RNases structurally resembled with each other and have identical amino acids at 48 out of 96 positions [53]. From these RNase, only RNase Sa3 strongly exerts toxicity towards a tumor cell line [22]. The cytotoxicity of Onconase is only 10-fold higher than RNase Sa3. The 3D structure of RNase Sa3 and Sa are quite similar to each other, however only RNase Sa3 is cytotoxic [54]. RNase cytotoxicity depends on some structural elements and sequence motifs. In case of α-Sacrin, a N-terminal β-hairpin present on it and absent in other non-toxic microbial RNase, is responsible for specific ribosomeinactivating protein. N-terminal β-hairpin of α-Sacrin is involved in protein-membrane interaction and has been shown to be perturbing [22]. RNase Sa may show toxic effect by replacement of Asp and Glu residues with Lsy which is present on the surface of RNase Sa [55]. These replacements produce mutant with variation in their charge, it was changed from acidic to basic protein by reversing five charges. By reversing charges, it could generate sufficient cytotoxic effects in RNase Sa [22,56]. Cytotoxic activity of RNase Sa correlates with the change in net charge from negative to positive. Site-directed mutation is one of approaches which allow the creation of charges on enzymes and there by produces toxic effect(s) with bare minimum side effects on cells. RNases mutant may be used as efficient therapeutics against cancer [19,20,22].
Mechanisms of RNase-mediated cytotoxicity towards cancer cells
Before internalisation, extracellular RNase interacts with the surface of target cell by membrane lipids, ion channels, receptors and non-specific electrostatic binding on the cell surface. Membrane proteins as well as lipids both are involved in interaction with RNase specifically. Onconase and BS-RNase have interaction with receptorlike site on plasma membrane. These receptor-like sites are non-protein receptor molecules that are not specific to other RNase [57]. For BSRNase, the existence of cellular membrane receptor has not been demonstrated but in previous studies there has been an evidence for existence of nonprotein receptor-like sites on the target cell surface. In some studies, it has been found that BS-RNase enters the cells by adsorption-mediated endocytosis (Figure 1). Interaction between BS-RNase and cellular membrane is facilitated by sulfhydryl–disulfide interchange reactions between cell surface sulfhydryl’s and the intersubunit disulfides that link the two subunits [58]. In gliome 9L cells line, the receptor for Onconase was reported that differed from amphibian lectin [59]. According to some authors, internalisation of Onconase is not suitable in HeLa cell line and little effect of protease treatment was noticed on the cells [60]. RNase A variant and HP-RNases are internalised in A4310r K562 cell line by endocytosis fluid-phase uptake process [59,61].
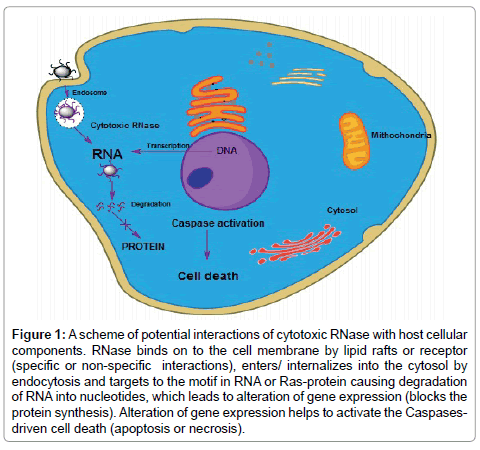
Figure 1: A scheme of potential interactions of cytotoxic RNase with host cellular components. RNase binds on to the cell membrane by lipid rafts or receptor (specific or non-specific interactions), enters/ internalizes into the cytosol by endocytosis and targets to the motif in RNA or Ras-protein causing degradation of RNA into nucleotides, which leads to alteration of gene expression (blocks the protein synthesis). Alteration of gene expression helps to activate the Caspasesdriven cell death (apoptosis or necrosis).
Role of bilayer membrane and cytosol
After binding to target cell surface, the cytotoxic RNase is internalised by endocytosis. Cytotoxic RNases have been localised in endosome but in some RNases cytotoxicity is blocked by energydependent process. Endocytic mechanism is still less understood but both Onconase and RNase A variants follow Clathrin and Dynaminindependent pathway. BS-RNase enzymes are transferred into malignant and non-malignant cells by endocytosis process to the extent but these enzymes are relatively more toxic to the malignant cells [27]. Observation of transportation of RNase from extracellular to cytosol follows two different approaches for RNases toxicity. In first approach, intercellular transport disrupts the cytotoxicity of RNase by drugs. Demonstration with Onconase indicated that intracellular routing happens in the absence of drugs, and is also independent of low pH environment. According to above observation, disrupting intercellular trafficking may account for indirect consequences on cellular pathways in the cell lines [27,58]. In case of BS-RNase, Monesin led to an increase in the cytotoxicity of enzymes in 9L gliome cells rather than Brefeldin A [59]. The second approach is based on the use of fluorescence label with specific marker for organelles to trace the intercellular route. This approach was implemented on Onconase and BS-RNase for tracking intercellular route in the target cells. RNases evade the cell membrane via non-protein receptor-dependent mechanisms or endocytosis or direct translocation. β-Amyloid proteins, prions, Calcitonin and unchaperoned positively charged molecules are able to directly translocate the RNase to the cytoplasm, or induces membrane damage and cell malfunction mediated by ion channel formation [22,62]. α-Sarcin molecules translocate into the cytosol by artificial lipid vessels that involved internalization through acidic endosomes in in vivo experiment. Endocytosis mediated acidic vessel is responsible for internalization of RNases form extracellular matrix to the cytoplasm. For α-sarcin, Onconase and toxic G88RR RNase A, the internalization mechanism appeared to be Clathrin-independent. Moreover, for Onconase and G88R RNase A it is Dynamin independent [27,43,63].
Degradation of RNA in cytosol
The third possible step of RNase pathway involved the degradation of the cellular RNA, and their transport from endosomes to the Golginetwork and the endoplasmic reticulum (ER) [57]. RNases have been shown to harbour ribonuclease activity in cytosol by its chemical modification or addition of residues through site-directed mutation(s) for greater stability of enzymes in cytosol environment. RI protein is found in cytosol, which quite likely acts as safeguard against extracellular RNase [27]. RNases have some residues which are sensitive to RI binding that prompts the blocking or inhibition of the ribonucleolytic activity of the RNase in the cytosol. In case of Onconase, it evades RI action in vivo, by neither increasing nor silencing the intracellular levels of RI thereby causing intense cytotoxicity. RI molecules being partially silenced in case of BS-RNase has been reported [64]. The dimeric structure of BS-RNase plays an important role because its dimeric forms are maintained by non-covalent interactions in the reducing environment of cytosol which are probably not inhibited by RI. BS-RNase progression from the endosome compartment to the Golgi complex was found in tumour cells but not in the normal cells [27,65]. The cytotoxic activity of RNase 1, α-Sacrin, BS-RNase, Onconase and G88RR severely disrupts the retrograde transport from the Golgi complex to ER, Angiogenin, RNase 1, BS RNase, Onconase and G88R RNase A [27,66,67]. The RNases are once translocated to the cytosol from the pre-ER compartment, they degrade RNA. It also seems that nuclear RNA is also possibly degraded by external RNase after electrostatic binding of nuclear membrane with the enzymes. Plasma membrane and endoplasmic reticulum contain RNA as an integral and structural part of membrane-associate ribosomes. Cytotoxicity of RNase might be due to cleavage of RNA molecules which leads to the protein synthesis inhibition and disruption of the regulatory processes in the cell cycles. For example, signal recognition particles, which target proteins to ER, contain RNA as a component of highly conserved RNAprotein core [22]. Hydrolysis of small non-RNA into micro-RNA causes the alteration of gene expression and thus causes the killing of malignant cells. Binase enzyme interaction with ionic pathways might be involved in the cell proliferation control and phenotypes. These interaction of Binase with the ionic pathways may block Ca2+ -activated K+ channels and thus inhibits the proliferation of ras-transformed fibroblasts, without any effect on the normal cells and in cells-transformed by src or fms oncogenes [22]. Thus, ras oncogene of expressing cells was more sensitive to Binase than the non-expressing cells. Likewise, Onconase exhibited cytotoxic activity towards ras-transformed mouse fibroblasts. These data suggested that Ras-targeting RNases and Ras proteins provide therapeutic possibilities to cancer therapy [67]. Degradation of cellular RNA by RNases arrests protein synthesis as well as induces apoptosis in the affected cells. In case of Onconase, there are two lines of evidences which triggered apoptosis i.e., cytotoxic and cytostatic effect [27]. Both effects of Onconase were observed apparently after 24-48 h of drug administration in the treated cells. Cylcoheximide or emetine was used for rapid inhibition of protein synthesis within 2-4 h incubation period. The growth of human histocytic lymphoma U93784 and leukaemia HL6019 cell lines was arrested by Onconase at G1/S checkpoint of the cellular cycle [68].
Ardelt and colleagues proposed that the degradation of RNA can trigger apoptosis pathways in malignant cells. They also observed that besides degrading t-RNA, micro-RNA or small interfering RNA produced from degraded RNA can play roles in specific cell regulation. Above studies illustrated that the enzyme-RNA interaction as well as other direct and mediated effects of cytotoxic RNases are important for understanding the mechanisms of RNase cytotoxicity towards malignant cells [17,62]. The above information helps to identify cellular targets of cytotoxic RNases, distinguishes between their direct and indirect effects of ribonucleolytic action. Caspase-dependent process, low molecular weight compounds, alteration in protein and NF-κB signal pathway are cell death mechanisms which involve in response to cytotoxic RNases [69]. In a previous study, Onconase decreased the NF- κB1 tanscription factor in pleural mesothelioma cells and restrained the canonical NF-κB dependent pathways. Binase induced cell death by reduced the mitochondrial potential and ligand-dependent apoptosis in Kasumi-1 and B-16 cells [68,70]. Mitochondrial potential reduced by binase via formation of mitochondrial pores activated the caspase 8 for increased Ca2+ level and decreased reactive oxygen species (ROS). It was also observed that tumor necrosis factor (TNF) present on B-16 cells surface help to increase in response to binase but TNF increased by factor of 16 in Kasumi-1 cells [71]. In a previous study it has been reported that genes of canonical NF-κB dependent signal pathway and proinflammatory caspase 1 and 4 gene were activated by binase. Binase shows the cytotoxic effect against cancer cell lines by activating the TNF through caspase 8 and NF-κB which leads to activate the caspase 3, 4 and 7 for eventual cell death [69,70]. Currently, we are still unclear which of these targets is more effective in triggering apoptotic process and thus more promising for cancer therapy. In conclusion, the role of cytotoxic RNases to target the cellular RNA in the cancer/ tumour cells by untraceable routes and pathways leading to cell death is still incomplete. In future, the RNase which was known as a nasty molecule to conduct mRNA purification and transcription studies may provide the scientists a promising alternative to develop non-mutagenic antitumour drugs with extended therapeutic applications.
RNase-ligand interaction mechanism
RNases are known as RNA-hydrolysing enzymes that exert numerous biological effects exogenously for degradation of cellular RNA apart from performing their main functions in the cell. RNase A (cow) and RNase 1 (human) have toxic effect for cancer cells due to acidic lipid raft, hepran sulphate and other glyan on cell surface. But selective toxicity of ribonuclease variants for malignant and non-malignant cells is still unclear [72]. In previously studies, unusual pattern of tumorassociated carbohydrate antigen (TACAs) in cancer cell surface helps to differentiate the cancer cells from normal cells (Table 2) [73]. TACA’s are expressed as membrane bound-glycolipids or glycoprotein and selected for vaccine design must be uniquely expressed on the surface of cancer cells but not on the normal cells. Acidic lipids, glycoproteins, heparin sulphate containing proteoglycans, actin, and RNA act as potentially acceptors for exogenous RNases on the cell surface. The cancer cell surface is more anionic than normal cell due to these larger exposures of membrane bounded glycosaminoglycan profile, phospholipid, and glycosphingolipid [72-74].
| Receptor | Binding site | Mechanism of internalization | References |
|---|---|---|---|
| Globo H (acidic lipid), sialic acid-containing glyco-proteins | Fucosylated glycan has significant affinity for RNase A and RNase1. | Endocytosis protein helps to initiated the endocytosis pathways | [72-74] |
| Heparan sulphate proteoglycans, chondroitin sulphate proteoglycans | RNases binds with heparan sulphate proteoglycan due to acidic or coulombic interaction. | Endocytosis protein helps to initiated the endocytosis pathways | [72-74] |
| TAg (Virus T antigen) |
TAg act as receptor which is covalently bound with RNA via a phosphodiester bond between the β-hydroxyl group of a serine residue and the 5'-phosphate. | Cytotoxicity exerts on cancerous cell without internalization | [72,74] |
| Hyaluronic acid modified RNase A (RNase A-HA) | Lysine residues mounting the negative charge on protein and facilitating its electrostatic complexation with cationic lipid nanoparticles,due to RNase A-HA complex can exclusively bind to the CD44 receptor. | RNase A-HA binds with CD44 receptor which activates the encocytosis pathways for cellular uptake of RNaseA-HA. | [69,75] |
Table 2: RNase-receptor interaction on cancerous cell surface.
Globo H is a tumor-associated neutral hexasaccharide glycosphingolipid, which acts as receptor/ligand for RNase A and RNase 1. Globo H is located endogenously on the outer membrane of epithelial cells of mammary, uterine, gut, kidney tissues and pancreas (Figure 2). RNase 1 is a major secretory ribonuclease from human which strongly interacts with a human cell-surface Globo H glycan [72]. Globo H, sialic acid and heparin sulphate play vital role in the cellular uptake of RNases due to increasing anionic nature of cancerous cell surface. RNases cationic side-chain interacts with glycan or sialic acid or lipids of cancerous cell-surface with nonspecific Coulombic interactions. Globo H has been reported as a more specific ligand for RNase 1 than RNase A. RNase 1 and RNase A have 82% identity in their amino acid sequence, however Val52 and Asn 76 are different in RNase A. Tumorigenesis changes the cell-surface components and also amplifies the toxicity that is based on differential exposure of Globo H and sulphated glycosaminoglycans on the cell surface. The Globo H and sulphate glycosaminoglycans interact distal from the enzymic active site of RNases. Globo H and other cell components are displayed on the interior of endosomes following endocytosis. The previous studies have suggested that the RNase 1 involves binding of lumenal Globo H, and this complex is internalized by endocytosis pathway into the cytosol, which eventually leads to degradation of cellular RNA. RNase A interacts tightly with abundant cell-surface proteoglycans containing glycosaminoglycans, such as heparan sulfate and chondroitin sulfate, as well as with sialic acid-containing glycoproteins [71,74]. Cancer cells, however, undertake constitutive endocytosis more hastily than noncancerous cells. Virus T antigen (TAg) also known as an onco- protein is exposed on the surface of MLE12 pneumocytes. TAg is covalently bound with RNA via a phosphodiester bond between 5’-phosphate and the β-hydroxyl group of a serine residue, and this complex acts as a receptor for RNase [74]. The RNase degraded the RNA which was associated with TAg onco-protein of transformed mouse cells into ribonucleoside phosphates. Binase binds with TAg onco-protein which is present in MLE12 pneumocytes membrane as receptor and hydrolyzes the TAg-associated RNA. In MLE12 pneumocytes, Binase exerts a cytotoxic effect even without its internalization, while Binase was insensitive to non-transformed type II pneumocyte cell lines [74].
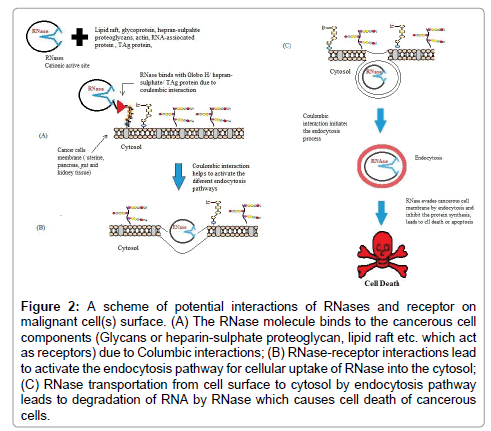
Figure 2: A scheme of potential interactions of RNases and receptor on malignant cell(s) surface. (A) The RNase molecule binds to the cancerous cell components (Glycans or heparin-sulphate proteoglycan, lipid raft etc. which act as receptors) due to Columbic interactions; (B) RNase-receptor interactions lead to activate the endocytosis pathway for cellular uptake of RNase into the cytosol; (C) RNase transportation from cell surface to cytosol by endocytosis pathway leads to degradation of RNA by RNase which causes cell death of cancerous cells.
Hyaluronic acid (HA)-RNase A is a nano-complex with cationic lipid molecules which inhibits the cancer proliferation. RNase A binds with HA due to supra-molecular interaction with carrier lipoids which promotes protein encapsulation efficacy, and facilitates cancer cell targeting via interaction with overexpressed CD44. HA-RNase A can specifically bind to the CD44 receptor through Coulombic interactions, and initiates the endocytosis pathways for cellular uptake of RNase. These nano-complexes are also used for drug delivery to the cancer cells. In previous studies, HA-RNase A was efficiently delivered to the CD44-overexpressing A549 cells which extensively inhibited cancer cells proliferation, indicating an efficient method of targeted cancer therapy [69,75].
The RNases as antitumour drugs or therapeutic agents have the ability to eliminate the tumour cells. RNase cytotoxic effects are based on selective hydrolysis of RNA, selective intracellular routing and membrane-specific recognition processes. Onconase is the only RNase that has been evaluated in clinical trial but yet it has obvious limitations. The cytotoxic pathways of RNases serve as working platform(s) for the creation of new anticancer drugs in future. By engineering process and chemical modification in ligand/receptor an increase in the cytotoxic properties of RNase molecules may be achieved to selectively allow the cytotoxic pathway to work in the malignant cells. The greater insight in the working mechanisms involved in the efficient uptake and subsequent cytotoxicity of α-sarcin and RIPs in the tumor/ malignant cells have suggested a potent role to these microbial RNases in the anticancer therapy [76]. There is a strong belief that further development in RNase studies will be the prerequisite for development of effective drug(s) against malignant cells and workable anticancer therapeutics.