Pancreatic Disorders & Therapy
Open Access
ISSN: 2165-7092
ISSN: 2165-7092
Review Article - (2014) Volume 4, Issue 3
All-trans-Retinoic Acid (atRA) is a biologically active derivative of vitamin A that regulates numerous physiological processes through its interaction with nuclear retinoid receptor proteins, termed as Retinoid Acid Receptors (RARs) and Retinoid X Receptors (RXR). Retinoid signaling is diverse and its role in embryonic development, adult growth and development, maintenance of immunity and epithelial barriers, and vision has been elucidated. An increased body of evidence suggests that altered metabolism of retinoic acid under experimental type-1 diabetes conditions induced with streptozotocin (STZ) is related to insulin deficiency. In several experimental approaches the role of treatment in STZ-induced diabetes has been tested. This review summarizes current knowledge on the role of retinoids and atRA in the improvement of pathological alterations in STZ-induced experimental type-1 diabetes in kidney, retina, skin and nervous system.
<Keywords: All-trans-retinoic acid; Diabetes, Streptozotocin; β cell injury; Kidney damage
Vitamin A or retinol is a fat-soluble compound derived from [beta]-carotene found in plants and retinyl esters from animal sources. The human body obtains vitamin A from two sources: preformed vitamin A (retinol and retinyl esters) and provitamin A carotenoids (β-carotene, α-carotene and β-cryptoxanthin) [1,2].
All-trans-retinoic acid (atRA) is a derivative of vitamin A and it is required for almost all essential physiological processes and functions because of its involvement in transcriptional regulation of over 530 different genes [3,4]. atRA exerts its actions by serving as an activating ligand of nuclear atRA receptors [retinoid acid receptor (RAR) α, RARβ, and RARγ] and peroxisome proliferator-activated receptor (PPAR) β/δ which form heterodimers with retinoid X receptors (RXR) [5,6]. atRA plays a relevant role in tissue development and differentiation [7]. This review focuses on the role of atRA in the alterations secondary to β cell damage induced by streptozotocin.
Vitamin A and its metabolites (retinoids) are a group of potent natural or synthetic molecules which exert a number of biological activities, including embryonic development, adult growth and development, maintenance of immunity, maintenance of epithelial barries, and vision [8]. Dietary retinyl esters are hydrolyzed in the intestine, and retinol taken into the enterocyte is reesterified. Retinyl ester is further secreted into the circulation and transported as retinol bound to retinol-binding protein (RBP4) to its target cells [9]. Studies have shown that retinol enters by diffusion [10]. However, in 2007, a cell surface receptor for RBP4 termed STRA6 (stimulated by retinoic acid 6) was identified, STRA6 is a widely expressed multi-transmembrane domain protein, it binds to RBP with high affinity and has robust vitamin A uptake activity from the vitamin A–RBP complex. It is widely expressed in embryonic development and in adult organ systems [11]. A human genetic study found that mutations in the human STRA6 gene are associated with widespread birth defects in multiple organ systems [12]. This is consistent with the expression of STRA6 and the diverse functions of vitamin A in embryonic development
Most of Vitamin A actions depend on its active metabolites, mainly atRA and 9-cis-RA [13,14], formed in the target tissues mostly through the intracellular oxidative metabolism [15]. Intracellularly, retinoic acid (RA) is subsequently converted to atRA, which can be isomerized through non-enzymatic process to form 9-cis-RA isomer. atRA is produced from retinol in two oxidative steps: first, retinol is oxidized to retinaldehyde, and then retinaldehyde is oxidized to atRA. The first step, the oxidation of retinol to retinaldehyde is catalyzed by two enzyme families, the cytosolic alcohol dehydrogenases (ADHs) and microsomal retinol dehydrogenases (RDHs) and is generally considered rate-limiting [16]. Retinaldehyde can be converted back to retinol, but the oxidation of retinaldehyde to atRA is irreversible, this latter reaction is catalyzed by three retinaldehyde dehydrogenases (RALDH1, RALDH2 and RALDH3)
Two forms of retinoic acid, atRA and 9-cis retinoic acid (9-RA), serve as ligands for two families of nuclear receptors: RAR (RARα, β, and γ) and RXR (RXRα, β, and γ). In vitro binding studies have demonstrated that RARs bind to atRA with high affinity, whereas 9-RA is a bifunctional ligand, which can bind to and activate both RARs and RXRs. However, it has not been demonstrated a relationship between the structure of atRA and 9-cis-RA with the affinity for RARs and RXRs. Following ligand binding, these compounds interact with cis acting DNA sequences called retinoic acid responsive elements in the promoter regions of target genes, thereby regulating gene expression (Figure 1) [17].
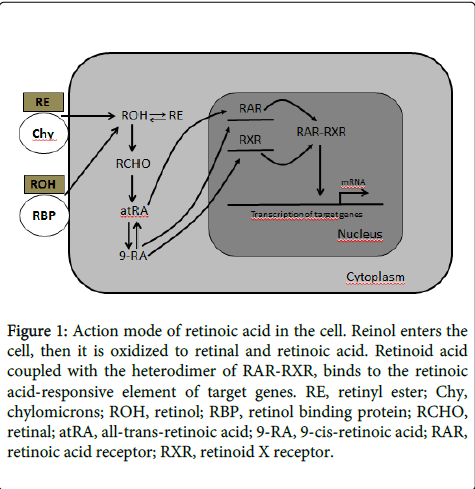
Figure 1: Action mode of retinoic acid in the cell. Reinol enters the cell, then it is oxidized to retinal and retinoic acid. Retinoid acid coupled with the heterodimer of RAR-RXR, binds to the retinoic acid-responsive element of target genes. RE, retinyl ester; Chy, chylomicrons; ROH, retinol; RBP, retinol binding protein; RCHO, retinal; atRA, all-trans-retinoic acid; 9-RA, 9-cis-retinoic acid; RAR, retinoic acid receptor; RXR, retinoid X receptor.
Streptozotocin (STZ) (2-deoxy-2-(3-methyl-3-nitrosourea)-1-D-glucopyranose) is a naturally occurring compound, produced by the soil bacterium streptomyces achromogenes, that exhibits broad spectrum of antibacterial properties [18]. STZ is a cytotoxic glucose analogue. After its discovery, it was used as a chemotherapeutic alkylating agent in the treatment of metastasizing pancreatic islet cell tumors and other malignancies such as: small cell lung cancer, lymphomas, mycosis fungoides, multiple myeloma, glioma and malignant melanoma [19]. Rakieten and colleagues reported the diabetogenic properties of STZ in 1963 [20]. From that time of discovery till date, STZ has been one of the chemical agents for the induction of diabetes in experimental animals. STZ induces diabetes in rats, mice, monkeys, hamsters, rabbits and guinea pigs [18]. STZ is cytotoxic to pancreatic β-cells and its effects are present within seventy two hours after administration depending on the dose used [21]. STZ toxic action involves its uptake into cells
The selective pancreatic beta cell toxicity and the diabetic condition, resulting from STZ induction, are related to the glucose moiety in its chemical structure, which enables STZ to enter the beta cell via the low affinity glucose 2 transporter (GLUT2) in the plasma membrane. In contrast, in insulin-producing cells not expressing the GLUT2, the cellular uptake of STZ is very slow. Correspondingly low is the toxicity [22] because the β-cells of the pancreas are more active than other cells in taking up glucose and so are more sensitive than other cells to STZ challenge.
STZ is a structural analogue of glucose (Glu) and N-acetyl Glucosamine (GlcNAc). STZ causes β-cell death by DNA fragmentation due to the nitrosourea moiety. Three major pathways associated with cell death are: (i) DNA methylation resulting in the activation of the nuclear enzyme poly ADP-ribose synthetase as part of the cell repair mechanism and consequently, NAD+ depletion; (ii) Nitric oxide production, and (iii) Free radical generation such as hydrogen peroxide [23,24]. The American Diabetes Association established an etiologic classification of Diabetes mellitus and based on their classification, four groups were proposed: 1) Type 1 (5–10%); 2) Type 2 (90–95%); 3) Other specific types and 4) Gestational. Thus, STZ-induced diabetes belongs to the category of other specific types or drug (chemical) induced diabetes.
The type of diabetes induced by STZ is controversial since STZ-hyperglycemia can be similar to either type I or type II diabetes mellitus [25]. Type I diabetes is an autoimmune disease leading to the destruction of the insulin producing pancreatic beta cells in the islets of Langerhans. Type I diabetes is most commonly diagnosed in children and young adults, and by the time of diagnosis, patients have very little endogenous insulin production, many researchers conclude that STZ produces type I diabetes mellitus [26,27].
On the other hand, the dose of STZ required for inducing diabetes depends on the animal species, age of animal, route of administration, weight of animal, nutritional status and different responses to xenobiotics.
Several studies have pointed out that atRA acts as an important signaling molecule for mesenchymal/epithelial interactions in the development of kidney, lung, central nervous system and gut [28-31]. Also, retinoids have been considered as insulinotropic factors [32,33] or its deficiency is related to the cause of type-1 diabetes [34,35]. It has been shown that at embryonic day (e) 11.5 of mice, atRA is endogenously and exclusively present in pancreatic mesenchyme, made evident by mRNA and protein expression of retinaldehyde dehydrogenase 2 (RALDH2) enzyme. In the presence of exogenous atRA, pancreatic rudiments differentiate into ducts and endocrine cells and inhibit acini. Furthermore, atRA upregulates pancreatic duodenum homeobox (PDX)-1, an important transcription factor in pancreatic development. These data suggest the important roles of atRA in determining the cell fate of pancreatic progenitor cells, leading to the proper formation of endocrine versus exocrine pancreas during organogenesis [36]
Also, it is possible to induce pancreatic differentiation of mouse Embryoid Body-Like Sphere (EBS) by simultaneous stimulation with activin and retinoic acid [37]. In fact, using activin to induce the differentiation of undifferentiated Embryonic Stem (ES) cells into endoderm and induction of pancreatic differentiation with retinoic acid are important elements that are common to almost all of the methods for obtain pancretic β cells from human ES cells and induced Pluripotent Stem Cells (iPS) [38]
There is scant information about the metabolism of atRA in diabetes, including diabetic nephropathy. Starkey and colleagues [39] recently described altered retinoic acid metabolism under diabetic conditions and suggested that a shift in atRA metabolism is a novel feature in type-2 diabetic renal disease. Ingenuity Pathway Analysis identified altered retinoic acid as a key-signaling axis that was altered in the diabetic renal cortical proteome. Western blotting and real-time PCR confirmed diabetes-induced upregulation of RALDH1, which was localized by immunofluorescence predominantly to the proximal tubule in the diabetic renal cortex, while PCR confirmed the down regulation of Alcohol Dehydrogenase (ADH) identified by mass spectrometry. Despite increased renal cortical tissue levels of retinol and RALDH1 in db/db versus control mice, atRA was significantly decreased in association with a significant decrease in PPARβ/δ mRNA [39]. Also, plasma, and kidney concentrations of Retinol Binding Protein (RBP) are significantly lower in STZ-treated rats compared to controls, suggesting that STZ-induced diabetes is associated with a depressed plasma concentration of retinol which may be due, at least in part, to its impaired metabolic transport from the liver [40]. It has long been established that 66–75% of dietary retinoid (chylomicron and chylomicron remnant retinoid) is taken up by the liver where it is stored in Hepatic Stellate Cells (HSCs) [41]. Evidence to date suggests that reduced metabolic availability of vitamin A occurs predominantly as a result of insulin deficiency
The role of atRA treatment on STZ-induced experimental diabetic nephropathy has been tested by Han et al., who described that atRA treatment decreased diabetes-induced renal expression and urinary excretion of monocyte chemoattractant peptide (MCP)-1 and Albumin: Creatinine Ratio (ACR). Also, in cultured podocytes, high glucose stimuli rapidly increased MCP-1 mRNA and protein expression, which was attenuated by atRA, suggesting an anti-inflammatory and renoprotective role of atRA in the early stages of diabetic nephropathy [42]
A protective role of atRA has been shown in diabetic and nondiabetic proteinuric diseases [42] and a link between a cytochrome P450 enzyme known to metabolize atRA and increased mitochondrial oxidative stress in type 1 diabetic rat kidneys has been established, since P450 enzyme increases the elimination of atRA from the body [43]. Recently, atRA has been shown to bind PPARβ/δ and act as a ligand to activate transcription, suggesting that altered retinoic metabolism could provide a potential link to insulin resistance and fatty acid [43]
Glucose-induced Endothelial Nitric Oxide Synthase (eNOS) expression and NO production in mesangial cells may contribute to hyperfiltration in diabetes and RA may exert beneficial effects by downregulation of Stromal Interaction Molecule 1 (STIM1) and store-operated Ca2+ influx (SOC) [44].
On the other hand, it has been reported that skin disorders in STZ-induced type 1 diabetes might be partially due to the reduced levels of vitamin A, which might exert protective effects against skin changes induced by diabetes. Treatment with vitamin A or RA influences various physiological processes in skin tissues, including enhancement of cell communication, and inhibition of metabolic activation. RA treatment reduced the activity of metalloproteinase and hyaluronidase in the skin tissues of diabetic rats. Also, blood retinol levels in STZ-treated rats were lower than controls, suggesting that type-1 diabetic rats could be vitamin A-deficient [45]. Also, superficial skin wounds in diabetic rats heal more rapidly in animals that have been pretreated with a regimen of topical atRA. At the histological level, recently healed skin from vehicle-treated diabetic rats contains a thin, wispy provisional matrix in which many of the embedded cells were round and some of them were pycnotic. In contrast, a much denser provisional matrix with large numbers of embedded spindle-shaped cells was observed in healed wounds from diabetic skin that had been pretreated with atRA. The atRA-treated diabetic skin was histologically similar to vehicle-treated skin from nondiabetic animals. In light of these findings, prophylactic use of retinoid-containing preparations might be useful in preventing the development of non-healing skin ulcers resultant from minor traumas in at-risk skin [46]
In a model of diabetic mice neuropathy induced by STZ administration, atRA treatment reverted the ultrastructural morphologic changes, as observed by the improvement in sensibility and the reduction in neuropathy by increasing the Neural Growth Factor (NGF) concentrations in nerve terminals [47,48]. Also, in a mouse model of STZ-induced dementia of Alzheimer´s type, atRA attenuated memory deficits by virtue of its neuroprotective, anti-cholinesterase, anti-oxidative, anti-inflammatory and probably amyloid lowering potential [49]. Retinol treatment decreased lipid peroxidation in the retina of STZ-treated animals, and improved the loss of fat-soluble antioxidants determined by the ferric chloride-bipyridyl reaction. It seems permissible to assume that retinoids may be involved in physiological protective mechanisms against lipid peroxidation in the retina in addition to their photo-receptive functions as visual pigment [50]. Also, atRA treatment decreased the number of apoptotic cells in the retina of STZ-treated mice evaluated by TdT-dUTP terminal nick-end labeling assay [51]. It was found that atRA exerts immunomodulatory actions in a number of inflammatory and autoimmune conditions, atRA reduced emergence of primed (autoreactive) CD4+ CD25+ T cells and reduced Th1/Th17 response and nitric oxide production in peripheral lymphoid tissues thus shifting the balance towards the anti-inflammatory cytokines [52]
Several doses have been used to test the protective effect of atRA on STZ-induced alterations, which are in the range of 1-20 mg/kg/day at times between 1-8 weeks [42,46-52] in animal models. However, the results of several classical clinical studies showed that chronic administration of vitamin A in slight excess of 5,000 IU/d is associated with a reduction in bone density and increased risk for osteoporotic fractures in the older individuals [53]. Also, castrated mice injected intraperitoneally with 10 mg/kg daily during 3 weeks with atRA showed significant bone loss, this effect was more pronounced in testosterone-deficient animals. Testosterone deficiency as occurs following castration may sensitize the bone to resorption mediated by atRA. Therefore, chronic vitamin A administration may be a risk factor for osteoporosis in rodents [54].
Retinoids have many important and diverse functions throughout the body, including roles in vision, regulation of cell proliferation and differentiation. The experiments described above provide evidence that atRA supplementation under diabetic conditions exerts protective effects and ameliorates pathological alterations in diabetes by modulating several signaling pathways including anti-inflammatory, antioxidant, immunomodulatory and antiapoptotic properties, thus suggesting a promising role of atRA as a potential chemo-therapeutic or chemo-preventive agent against diverse complications in diabetes.
With partial support from Consejo Nacional de Ciencia y Tecnología (Conacyt). México (Grant 179870 to JLR)