Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2017) Volume 7, Issue 2
Dendrobium sonia-28 is an important ornamental orchid in the flower industry. In vitro mutagenesis by means of gamma irradiation can be used to produce economically improved mutants. Protocorm-like bodies (PLBs) of Dendrobium sonia-28 were subjected to gamma irradiation at doses of 10 to 200 Gy gamma rays. Results showed that PLBs survival rate and weight were inversely related to the irradiation doses. In untreated PLBs, survival rate and weight were tenfold and fivefold higher as compared to 200 irradiated PLBs. Results indicated that the radio sensitivity test (LD50) for the PLBs were approximately at 43 Gy. There were appreciable variations in plantlet structure and morphology relative to increase irradiation doses. Plantlets infected with low doses of gamma radiation had better development of the shoot and root as well as the foliage. SEM and TEM analysis disclosed severe surface and cell organelles damage and stomatal closure in protocorm-like bodies (PLBs) infected with the high irradiation dose. Therefore, the findings of this study have proven an efficient method of in vitro mutagenesis through gamma radiation. The results obtained in this study suggested that PLBs survival rate, growth rate percentage, morphological characteristics as well as techniques such as SEM and TEM may be used in early assessment of effectiveness and superiority of radiation dose to induce mutations. Moreover, Some RAPD markers clearly discriminated between selected and non-selected variants of Dendrobium sonia-28, showing different banding patterns for each gamma irradiation doses.
<Keywords: Dendrobium sonia-28; Gamma irradiation; Mutation; PLBs
Gy: Gray; MS: Murashige and Skoog; PLBs: Protocorm-Like Bodies; RAPD: Randomly Amplified Polymorphic DNA; SEM: Scanning Electron Microscope; TEM: Transmission Electron Microscope
The flowering plant family of Orchidaceae is the most species-rich of plant families with more than 25,000 species in an estimated 800 genera [1]. The orchid genus Dendrobium increasing popularity is owed to its floriferous flower sprays, year round availability, long flowering life; as well as the genus’s wide spectrum of colours, sizes and shapes [2-4]. Similarly, Dendrobium Sonia-28 orchid hybrid which can be created by crossing the two hybrids Dendrobium Caesar and Dendrobium Tomie Drake is cherished for its pink-coloured blossoms as well as the quality of the cut florae [5-7].
Dendrobium orchids are sensitive to pests and diseases. Latiffah et al. [8] stated that root discolourification and a yellowing of the stem are strong indications of mold, chiefly associated with species from the genus Fusarium such as Fusarium proliferatum . Currently, there are no economically feasible biological, chemical, or cultural methods of controlling Fusarium wilt in an infected field of crop [9,10]. In a controlled environment not limited by seasonal and environmental constraints, the culture of tissue may provide the opportunity of a reduction in the selected period [11].
Furthermore, mutation breeding joined with tissue culture has made a substantial impact to the breeding of plants by presenting new processes in relation to the propagation and induction of genetic variants [12]. Ahmad et al. [5] stated that one way for expanding the genetic variety and alternative breeding strategy within in vitro conditions is the use of chemical as well as physical mutagens; such examples are ethyl methane sulfonate (EMS), ethyl ethane sulfonate (EES) and ionized radiation such as gamma radiation and neutron rays. The application of mutagens during an in vitro culture may intensify the genetic diversity; which will expand the process of selecting favorable genotypes, therefore, speeding up the duration of breeding programs [13]. Recently, mutation breeding has been used for some important ornamental plants such as: chrysanthemums [14], orchids [15] roses [16], pelargoniums [17] and canna [18]. Tokuhara et al. [19] reported that the breeding of orchids has been conducted for more than 100 years, and recent in vitro cultured organisms inducing mutation have been increased. The use of the above mentioned mutagens is mentioned in regards to the tolerance to cold and salt of rice [20], the acceptance of cold temperature of canola [21], the resistance to drought of wheat [22] and the tolerance to cold and saline conditions in cauliflower species [23]. Until now, some orchids have been successfully generated via mutagenesis, which among them are four new varieties of Dendrobium Sonia have been developed in Malaysia [5].
Hameed et al. [24] stated that physiological and biochemical processes processed by plants are considerably affected by gammairradiation stress; the extent of the mutations to the morphological, structural and functional features is determined by the strength as well as the extent of the gamma-irradiation stress. The results of moderate stress in the form of gamma radiation will not hinder the adaptive capacity of the individual specimen. Bae et al. [12] reported that heavy-ion beam irradiation produce abnormal chlorophyll containing leaves from irradiated rhizomes in two species of Cymbidium orchid.
In order to accomplish the goal of acceleration of the progress within orchid breeding programs regarding resistance to Fusarium wilt, it is crucial to improve bioassay that can distinguish between resistant and susceptible cultivars competently as well as accurately. Consequently, the current study was undertaken to probe and investigate into the use of gamma radiation being utilized as a mutagen for Dendrobium Sonia-28 PLBs with the future goal of the selecting Fusarium wilt disease tolerant orchids.
Propagation of plant materials
Semi-solid, half- strength MS medium [25] supplemented with 2% sucrose, 2.75 g L-1, Gelrite TM (Duchefa, the Netherlands) and 1 mg L-1 6-benzylaminopurine (BAP; Duchefa) was prepared. The PH value of media was adjusted to 5.7 to 5.8 prior to autoclaving (STURDY SA-300VFA-F-A505, Sturdy Industrial Co. Ltd., Taiwan). In vitro culture of protocorm-like bodies (PLBs) of Dendrobium Sonia-28 was initiated by aseptically culturing shoot tips of the plant in the above mentioned media. The cultures were incubated at 25 ± 2ºC under 16 h photoperiod using cool white fluorescent light of 30 μmol m-2 s-1 per day. Newly developed healthy PLBs in size of 3-4 mm were used as explants for the following experiments.
Gamma radiation on Dendrobium Sonia-28 PLBs
In order to determine PLB’s gamma radiation sensitivity, four weeks old of 3-4 mm PLBs were irradiated with acute gamma irradiation. The source of gamma rays was Cobalt 60. Dendrobium Sonia-28’s PLBs were acutely irradiated with different doses of 10, 20, 30, 40, 50, 60, 80, 100, 150 and 200 Gy (10 replicate containing 10 PLBs for each dose). The process of irradiation was carried out at Agro technology and Bioscience Division, Malaysian Nuclear Agency, Bangi, Selangor. PLBs were immediately transferred individually to fresh half strength MS medium and were maintained at 25 ± 2°C with the photoperiod of 16 hours light and 8 hours dark for growth regeneration purpose. The first vegetative generation in which treatment was performed was referred to as M1V1. The study continued until the fourth generation (M1V4) to see the stability of the induced traits before selecting as mutated plants. Brunner [16] indicated that selection of mutants usually starts in the second or later generation for multiplication of the vegetative plants.
Radiation sensitivity test
For initiation and selection of tolerance to radiation, survival PLBs after four weeks, which were green in appearance were selected. The survival rate and growth rate of growing PLBs were scored at the end of the experiment. LD 50 was calculated based on the number of surviving PLBs at four weeks after irradiation. The latter was multiplied on the multiplication medium.
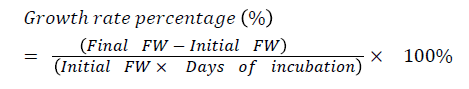
Scanning electron microscope (SEM) observation of PLBs
Comparison of PLBs surface and stomatal behavior between irradiated and control PLBs were made using scanning electron microscope observation through the freeze drying method. Subsequently, after four weeks of irradiation, the specimens were subjected to SEM analysis. Stomata width of 30 stomata was recorded in two PLBs. Consequently, the fresh experiment samples were removed from MS medium, PLBs then were dabbed dry with a piece of filter paper, finally affixed on double-sided strips of tape; were later placed upon the sample stub. The VP SEM was initiated with a saturated humidity of 4°C; the apparatus consisted of a Peltier cooling stage possessing a gas pressure of 500-700 Pa. The tension value was assessed at an extra high value of 15 kV. The producing PLBs were later viewed using a Leo Supra 50 VP Field Emission scanning electron microscope (Carl Zeiss SMT, Germany).
Transmission electron microscope (TEM) observation of PLBs
To begin the process, the PLBs were situated within a McDowell- Trump fixative, which was prepared in a 0.1 M phosphate buffer at a temperature of 40ºC for 24 h. The next day, PLBs were washed three times with the 0.1M phosphate buffer. Then, PLBs were then treated with a 1% osmium tetraoxide solution at room temperature for 2 h. The product samples were washed in distilled water. Following stage, the PLBs were dehydrated by a graded series of ethanol. A resin mixture was permeated into the samples via a rotator. Eventually, the produced PLBs were further combined in a new ratio of the Spurr’s mix in the rotator. The above mentioned samples were also inserted in resin beads which were formed in the mold and finally, the samples were cured at a temperature of 60ºC for 12 to 48 days. With an ultramicrotome cutter, PLBs were then dissected and gathered on carbon planchette. Finally, the samples were stained for viewing the specimens under the transmission electron microscope.
DNA extraction
DNA was extracted from gamma irradiated and controls Dendrobium Sonia-28 PLBs using a Promega Wizard® Genomic DNA Purification Kit according to manufacturer’s instructions.
For RAPD analysis, 40 oligonucleotide primers were tested with only 5 primers produced bands in gamma treated PLBs (Table 1). The primers were obtained from The Custom Biology CompanyTM (Integrated DNA Technologies). The PCR consumables and buffers used in the RAPD analysis were obtained from Fermentas Life Sciences (Vilnius, Lithuania). The quantity and purity of the isolated DNA was checked using a UV spectrophotometer (Quawell UV spectrophotometer Q3000, Korea). The ratio of absorbance at two wavelengths (A 260 and A 280) was compared with the standard ratio of pure DNA. The quantities of the DNA isolated were found to be optimal for further PCR amplification.
| Primers | Sequence (5’-3’) | G+C Content (%) | TM (°C) |
|---|---|---|---|
| OPA04 | AATCGGGCTG | 60 | 39.5 |
| OPAW13 | CTACGATGCC | 60 | 39.5 |
| OPAW17 | TGCTGCTGCC | 70 | 43.6 |
| OPB02 | TGATCCCTGG | 60 | 39.5 |
| OPB05 | TGCGCCCTTC | 70 | 43.6 |
| OPB06 | TGCTCTGCCC | 70 | 43.6 |
| OPB08 | GTCCACACGG | 70 | 43.6 |
| OPB11 | GTAGACCCGT | 60 | 39.5 |
| OPB12 | CCTTGACGCA | 60 | 39.5 |
| OPB13 | TCCCCCCGCT | 70 | 43.6 |
| OPB14 | TCCGCTCTGG | 70 | 43.6 |
| OPB17 | AGGGAACGAG | 60 | 39.5 |
| OPB18 | CCACAGCAGT | 60 | 39.5 |
| OPD01 | ACCGCGAGGG | 70 | 43.6 |
| OPG03 | GAGCCCTCCA | 70 | 43.6 |
| OPG13 | CTCTCCGCCA | 70 | 43.6 |
| OPG14 | GGATGAGACC | 70 | 43.6 |
| OPG15 | ACTGGGAACTC | 60 | 39.5 |
| OPZ04 | AGGCTGTGCT | 70 | 39.6 |
| OPZ09 | CACCCCAGTC | 70 | 43.6 |
Table 1: List of RAPD primers.
RAPD-PCR
The total volume of each RAPD-PCR reaction was 20 μL. The mixture consisted of 20 ng DNA template, one unit of Taq DNA polymerase (DreamTaq™ DNA Polymerase, 5 U μL-1), 0.5 μL of 10 mM deoxyribonucleotide triphosphate (dNTP; dNTP Mix), 2 μL of 10x PCR buffer (10x DreamTaq™ Buffer) containing 100 mM Tris- Hydrochloride (Tris-HCl) at pH 8.8, 500 mM potassium hydrochloride (KCl), 0.8% (v/v) Nonidet P40 and 20 mM magnesium chloride (MgCl2), 1 μL from 10 μM of a single primer and autoclaved distilled deionised water was used to top up the PCR volume to 20 μL. A MyCycler™ Thermal Cycler (Bio-Rad Laboratories, Inc., USA) was used to amplify the DNA samples. The PCR conditions were set with an initial denaturation at 95°C for 3 min. The PCR was performed for 35 cycles with denaturation at 95°C for 30 s, annealing at 5°C below each primer’s melting temperature (Tm) for 30 s and extension at 72°C for 1 min, followed by a final extension cycle at 72°C for 10 min. The PCR product was kept at 4°C and finally stored at -40°C prior to electrophoresis.
DNA electrophoresis
The DNA fragments produced were visualized in a 1.5% agarose gel under UV light and photographed using a Gel Documentation System (Biostep DH-20, Germany). The evaluation of the DNA fragment patterns of Dendrobium Sonia-28 PLBs was performed by calculating the Similarity Indices of the PLBs obtained from gamma-treated PLBs compared to controls. Reproducible bands were scored manually as 1 or 0 for presence or absence. The coefficients of similarity between the treatment and control samples were calculated using the formula: Similarity Index=2Nxy/(Nx+Ny), where Nxy=number of monomorphic bands between the control and treatment groups, Nx = total number of bands in the control group and Ny=total number of bands in the treatment group [26-28].
Morphological studies
Both control and irradiated Dendrobium Sonia-28 plantlets were subjected to morphological studies (except 200 Gy), whereby, the morphological appearance of the plantlets such as height and number of shoots, the number and length of fully opened leaves and the number and length of roots per each plantlets were determined. Meanwhile, all morphological characteristics were observed and recorded on month six of the inoculation. The length of shoot, leaf and root were assessed using a ruler and all characters were based on the mean of the shoot, leaf and root length of each selected plantlets. Three plantlets per replicate were subjected to the experiment with four replicates for each treatment.
Statistical analysis
Statistical analysis was performed using predictive analytic software SPSS (SPSS 16.0, IBM, US). Data collected were analyzed for the variant using one-way ANOVA and the differences were contrasted using Tukey’s multiple range tests at 5% significance level.
Effect of different gamma radiation doses on PLBs of Dendrobium Sonia-28
Results obtained from this study showed highly significant differences (P ≤ 0.01) were observed among the different doses of irradiation. In the present study, gradual reduction in survival rate and subsequent growth weight of treated population were observed with a corresponding increase in dose of radiation.
The survival rate of PLBs treated with gamma radiation decreased compared to the control ranged from 9.8 with the untreated control to 0.6 at 200 Gy. There was a significant decrease in survival rate of PLBs while a dose of gamma radiation increase (except 50 Gy) whereas PLBs irradiated with lower doses of gamma radiation (10 and 20 Gy) showed a negligible decrease of survival rates (Figure 1). Furthermore, LD 50 value of treating PLBs estimated at the end of four weeks was recorded at 43 Gy.

Figure 1: Effect of various doses of gamma irradiation on the survival rate of Dendrobium Sonia-28 orchid PLBs within four weeks. Letters a-f represent Significant differences at P<0.05.
Significant differences were observed between the growth rates of PLBs treated with various gamma radiation doses compared to control treatment after four weeks. Gamma radiation at the higher dose appeared to have a greater influence on PLB’s growth rate compared to control and treated PLBs at the dose of 10 and 20 Gy (Figure 2).

Figure 2: Effect of various doses of gamma radiation on the final fresh weight of Dendrobium Sonia-28 orchid PLBs (3-4 mm). Results were analysed using one-way ANOVA.
Radio-sensitivity test of Dendrobium Sonia-28 PLBs was determined by measuring survival rate and lethal dose (LD50) of the PLBs, four weeks after irradiation. The experimental design was a Completely Randomized Design and a significant treatment effect was determined by using the Analysis Of Variance (ANOVA).
LD50 graph was plotted in order to determine the optimal doses for mutagenesis. The decreasing trend of survival rate was clearly shown in Figure 3.

Figure 3: Effect of various doses of gamma radiation on the survival rate of Dendrobium Sonia-28 orchid PLBs (size 3-4 mm) and the LD50 level.
Surface morphological observations of PLBs
Figures 4 and 5 show the surface morphology of Dendrobium Sonia-28 PLBs under scanning electron microscope (SEM). SEM observation was initiated under light condition after one month of irradiation.

Figure 4: SEM of Dendrobium Sonia-28 PLB. (a) Untreated PLBs
surface; (b) the surface of PLBs treated with 10 Gy gamma
irradiation; (c) surface of PLBs treated with 20 Gy gamma
irradiation; (d) surface of PLBs treated with 30 Gy gamma
irradiation; (e) surface of PLBs treated with 40 Gy gamma
irradiation; (f) surface of PLBs treated with 50 Gy gamma
irradiation; (g) surface of PLBs treated with 60 Gy gamma
irradiation; (h) surface of PLBs treated with 80 Gy gamma
irradiation; (i) surface of PLBs treated with 100 Gy gamma
irradiation; (j) surface of PLBs treated with 150 Gy gamma
irradiation; (k) surface of PLBs treated with 200 Gy gamma
irradiation. Scale bar=100X.
The PLB surface rich in stomata could be seen clearly under SEM. Damage of the irradiated PLBs surface was intensified with increasing doses of radiation (Figure 4).

Figure 5: SEM of Dendrobium Sonia-28 PLB. (a) Untreated PLBs
stomata; (b) stomata of PLBs treated with 10 Gy gamma
irradiation ; (c) stomata of PLBs treated with 20 Gy gamma
irradiation; (d) stomata of PLBs treated with 30 Gy gamma
irradiation; € stomata of PLBs treated with 40 Gy gamma
irradiation; (f) stomata of PLBs treated with 50 Gy gamma
irradiation; (g) stomata of PLBs treated with 60 Gy gamma
irradiation; (h) stomata of PLBs treated with 80 Gy gamma
irradiation; (i) stomata of PLBs treated with 100 Gy gamma
irradiation; (j) stomata of PLBs treated with 150 Gy gamma
irradiation; (k) stomata of PLBs treated with 200 Gy gamma
irradiation Scale bar=1KX.
There was no significant effect of lower doses on stomata size. However, stomata in higher doses irradiated PLBs were significantly reduced in size (Table 2).
| Gamma irradiation doses (Gy) | Mean of stomatal aperture (µm) |
|---|---|
| 0 | 7.38 ± 0.56 a |
| 10 | 7.12 ± 0.98 a |
| 20 | 6.94 ± 0.82 ab |
| 30 | 7.01 ± 0.56 a |
| 40 | 6.12 ± 1.04 b |
| 50 | 5.67 ± 0.97 bc |
| 60 | 4.95 ± 1.23 c |
| 80 | 3.62 ± 0.78 d |
| 100 | 2.45 ± 1.39 e |
| 150 | 2.14 ± 0.51 ef |
| 200 | 1.84 ± 0.80 f |
Table 2: Effect of various doses of gamma radiation on stomatal mean aperture (µm) of Dendrobium Sonia-28 orchid PLBs after 30 days under light (Values within the column followed by different letter(s) are significantly different at p = 0.05 by Tukey’s multiple range test).
Gamma rays irradiation had a significant effect on pore width of stomata. Maximum mean stomatal width (7.38 ± 0.56 μm) was recorded in untreated plants, whereas the minimum mean stomatal width (1.84 ± 0.80 μm) was recorded at 200 Gy.
At lower gamma ray treatment, stomatal width was statistically at par with control stomatal width. Moreover, reducing of stomatal size in the present experiment is consistent with the general trend of the deleterious consequences of gamma irradiation on related growth parameters like leaf size.
TEM observation of Dendrobium Sonia-28 PLBs after gamma irradiation treatment
The effects of gamma radiation on PLBs cell were analyzed using transmission electron microscopy (TEM). The TEM analysis displayed that large central vacuole, undamaged chloroplast and mitochondria as well as intact plasma membrane were more observed in un-irradiated PLBs cells. The chloroplasts were more round and bold in untreated plants, whereas at higher dose (200 Gy), they were not in proper shape and were broken or fused. A significant difference was recorded in cell organelles after gamma irradiation. At higher dose gamma-treated PLBs, chloroplasts were more damaged, the cell walls were folded, the plasma membrane was broken and even the structure of the mitochondria and vacuoles were destroyed (Figure 6).

Figure 6: Transmission electron micrographs of Dendrobium Sonia-28 PLBs treated with 0, 20, 50 and 150 Gy of gamma radiation. (a) Normal cells showing typical arrangement of the organelles; (b) cell organelles in 20 Gy-treated PLBs; c cell organelles in 50 Gy-treated PLBs; d cell organelles in 150 Gy-treated PLBs. Scale bar=1, 2 µm. Arrows indicate various shapes of chloroplast, mitochondria and vacuoles. cw: cell wall, v: vacuole, m: mitochondria, c: chloroplast, cc: collapsed chloroplast.
Genetic diversity of irradiated plantlets
Five (5) different primers produced various mono and polymorphic bands and were used to assess and compare the genetic fidelity of PLBs obtained from irradiated PLBs compared with non-irradiated PLBs. The 5 primers produced 48, 46, 46, 45, 43, 46, 38, 39, 36, 32 and 31 bands in the control, 10, 20, 30, 40, 50, 60, 80, 100, 150 and 200 Gy irradiated PLBs, respectively (Figure 7).

Figure 7: Results for RAPD analysis of DNA samples obtained from untreated control PLBs (C), 10 Gy-irradiated PLBs, 20 Gy-irradiated PLBs, 30 Gy-irradiated PLBs, 40 Gy-irradiated PLBs, 50 Gy-irradiated PLBs, 60 Gy-irradiated PLBs, 80-Gy irradiated PLBs, 100 Gy-irradiated PLBs, 150 Gy-irradiated PLBs and 200 Gy-irradiated PLBs using the primers OPA04 (A), OPG15(B), OPAW13 (C), OPB02 (D) and OPB11(E). M=1 kb DNA ladder.
The sizes of these amplification products ranged between 100bp and 3000 bp. There were 2.1, 2.1, 12.5, 8.3, 12.5, 10.4, 4.2 and 6.3% polymorphic bands compared to controls (n=48) for the 30, 40, 50, 60, 80, 100, 150 and 200 Gy-irradiated PLBs. The similarity index (SI) value ranged from 0 to 1.0 (Table 3).
| Primers | SI for control and 10 Gy irradiated PLBs | SI for control and 20 Gy irradiated PLBs | SI for control and 30 Gy irradiated PLBs | SI for control and 40 Gy irradiated PLBs | SI for control and 50 Gy irradiated PLBs | SI for control and 60 Gy irradiated PLBs | SI for control and 80 Gy irradiated PLBs | SI for control and 100 Gy irradiated PLBs | SI for control and 150 Gy irradiated PLBs | SI for control and 200 Gy irradiated PLBs |
|---|---|---|---|---|---|---|---|---|---|---|
| OPA04 | 1 | 1 | 1 | 0.95 | 0.84 | 0.46 | 0.46 | 0.46 | 0.46 | 0.4 |
| OPG15 | 0.94 | 0.94 | 0.94 | 0.94 | 0.89 | 0.94 | 0.94 | 0.94 | 0.82 | 0.9 |
| OPAW13 | 1 | 1 | 0.96 | 0.96 | 0.89 | 0.96 | 0.86 | 0.85 | 0.92 | 0.9 |
| OPB02 | 1 | 1 | 1 | 0.94 | 0.94 | 0.89 | 0.94 | 0.94 | 0.88 | 0.8 |
| OPB11 | 0.93 | 0.93 | 0.8 | 0.77 | 0.62 | 0.46 | 0.5 | 0.55 | 0.4 | 0 |
Table 3: Similarity indices from RAPD analyses of DNA samples obtained from PLBs of Dendrobium Sonia-28 subjected to the various doses of gamma irradiation.
Morphological characteristics of in vitro plantlets upon gamma irradiation treatment
The association between the growth of irradiated PLBs and dose of gamma radiation has been demonstrated by investigating the morphological alteration and growth of the treated and untreated plantlets. The plantlet's morphological characteristics kept decreasing with increasing irradiation dosage for six months after irradiation, as illustrated in Table 4.
| Gamma doses (Gy) | No of shoots | Shoot length (cm) | No. of roots | Root length (cm) | No. of leaves | Leaf length (cm) |
|---|---|---|---|---|---|---|
| 0 | 2 ± 0.18ab | 0.7 ± 0.32a | 4 ± 0.56a | 1.3 ± 0.18a | 4 ± 0.94bcd | 2.4 ± 0.38a |
| 10 | 2 ± 0.40abc | 0.7 ± 0.20a | 4 ± 1.14ab | 1.2 ± 0.43ab | 4 ± 0.69bc | 1.8 ± 0.47b |
| 20 | 2 ± 0.32a | 0.8 ± 0.19a | 3 ± 1.13bc | 0.9±0.46bc | 5 ± 0.62a | 1.6 ± 0.42bc |
| 30 | 2 ± 0.211ab | 0.7 ± 0.153a | 3 ± 0.971bc | 0.8±0.384bc | 4 ± 0.597ab | 1.5 ± 0.388bc |
| 40 | 2 ± 0.189abcd | 0.7 ± 0.141ab | 3 ± 0.801cd | 0.8±0.357bc | 4 ± 0.52bcd | 1.4 ± 0.359c |
| 50 | 1 ± 0.31bcd | 0.5 ± 0.12bc | 3 ± 1.08bc | 0.9±0.51bc | 3 ± 0.40ef | 0.7 ± 0.21d |
| 60 | 1± 0.256cd | 0.4±0.063c | 3 ± 0.917cd | 0.8±0.419bc | 3± 0.358ef | 0.7 ± 0.196d |
| 80 | 1 ± 0.389d | 0.5 ± 0.26c | 3 ± 0.43cd | 0.8±0.62c | 3±0.49f | 0.8 ± 0.15d |
| 100 | 1 ± 0.42d | 0.3 ± 0.13c | 4 ± 0.33ab | 0.6±0.20c | 4±0.74cde | 0.7 ± 0.59d |
| 150 | 1 ± 0.21cd | 0.4 ±0.22c | 1 ± 0.85d | 0.1 ±0.10d | 3±0.82de | 0.5 ± 0.30d |
| 200 | 1 ± 0.24cd | 0.5 ± 0.9c | 3 ± 0.39cd | 0.2± 0.12d | 3 ±0.78de | 0.7 ± 0.23d |
Table 4: Effect of various doses of gamma radiation on the no. of shoots, shoot length (cm), no. of roots, root lengths (cm), no. of leaf and leaf length (cm) of Dendrobium Sonia-28 orchid plantlets after 6 months
Length and number of offshoots were decreased in plantlets after higher doses of gamma irradiation of PLBs. The highest number of shoots was obtained from the untreated and lower dose treated PLBs with around two off shoots while irradiation at higher doses produced only one shoot. Gamma irradiation postpone the plantlet shoot emergence, moreover, higher gamma doses reduce the mean number of shoots produced per plantlets. Generally shoot length of seedling was decreased gradually with increasing dose. Maximum shoot length was observed at untreated plantlets (0.8 cm) whereas highest shoot length decrease was observed after irradiation dose of 100 Gy (0.3 cm).
In this study, increasing doses of gamma radiation caused a decrease in the average number of plantlets leaves produced from Dendrobium Sonia-28 PLBs. There were only 3 leaves recorded for higher dose irradiated plantlets while for both control and treatments at lower dose of gamma irradiation, mean number of leaves are 4 and 5 leaves per plantlets. Furthermore, some dose specific leaf abnormalities such as change in leaf size were observed in the treated plantlets.
Root length also decreased after the higher doses of irradiation as compared to non-irradiated control in six month plantlets. The percentage of these stunted roots increased with increasing gamma ray doses, while untreated plantlets (control) showed better foliage appearance. Maximum decrease in root number was observed after 150 Gy dose of gamma radiation. Increasing dosage of irradiation caused root development to be postponed giving poor root size and less number of roots per plantlets.
While higher doses of gamma radiation inhibited plantlets growth, no significant morphological abnormalities were observed using lower doses of gamma rays. The growth of PLBs exposed to low-dose gamma irradiation was faintly increased compared with that of the control, while the seedling growth was noticeably decreased by a high-dose irradiation of 30 Gy. Lower dose treated plantlets showed the longest and highest mean number of shoot and leaf per plantlets within the same time period. Moreover, gamma ray treated PLBs produced the highest number and highest height of roots per plantlets (Figure 8).

Figure 8: Effect of gamma irradiation at different doses on Dendrobium Sonia-28 plantlets after 6 months of irradiation. (a) Untreated plantlet; (b) plantlet treated with 10 Gy gamma irradiation; (c) plantlet treated with 20 Gy gamma irradiation; (d) plantlet treated with 30 Gy gamma irradiation; (e) plantlet treated with 40 Gy gamma irradiation; (f) plantlet treated with 50 Gy gamma irradiation; (g) plantlet treated with 60 Gy gamma irradiation; (h) plantlet treated with 80 Gy gamma irradiation; (i) plantlet treated with 100 Gy gamma irradiation; (j) plantlet treated with 150 Gy gamma irradiation (Scale bar=1 cm).
Interaction of irradiation with other cellular molecules, like water cause producing of free radicals, which these free radicals could combine and form some toxic substances (H2O2), which contribute to the destruction of cells and cause plant death. This indirect effect is more important in vegetative cells where the cytoplasm contains about 80% water [29]. Generally, it was accepted that the survival rate andregeneration availability decreases while frequency of mutation increases [28]. Taheri et al. [29] reported that the mortality rate of three varieties of Curcuma alismatifolia increased with increasing gamma radiation dosage and mortality reached the highest rate at 100 Gy of radiation. After irradiation with 10 and 20 Gy of gamma irradiation, a stimulatory effect on growth and weight of in vitro Sugarcane callus was noticed, conversely, more necrotic callus were observed at higher doses of gamma radiation [30].
According to [31], regeneration rate of Rosa hybrid decreased from 47% to 0% when X-ray doses increases between 25 to 100 Gy. Tokuhara et al. [19] reported that the frequency of PLBs mortality increased with increasing absorbed dose of ion beam irradiation. In some Orchid species, all PLBs showed necrosis after irradiation with a dose higher than 50 Gy and more than 62% of the PLBs exhibited necrosis after a dose higher than 5 Gy. Ahmad et al. [5] reported that at lower doses of irradiation, PLBs fresh weights increase while PLBs fresh weight decreased after the higher dose of irradiation.
Afrasiab and Iqbal [2] reported that higher doses of gamma radiation reduce the amount of endogenous growth regulators like cytokines. Al-Safadi and Simon [9] reported similar findings regarding the increase of the callus weight compare to the control at the lower doses of gamma irradiation in carrot (Daucus carota L .). Biochemical and physiological changes can be induced by gamma irradiation which resulting in the faster vegetative growth of onion plants [7]. Hence, this could be due to the post effect mutation treatment of PLB’s final fresh weight increase at lower doses of gamma radiation in this study.
In general, ionizing radiation influences plant growth and development in living plants. Low doses of ionizing radiation are known to have stimulatory effects on growth hormones, cell division and plant growth, whereas high radiation levels induce increasingly harmful effects on vegetative growth, as well as pronounced reductions in reproductive fitness and yields [32,33].
Moghaddam et al. [34] reported that atoms and molecules interacted with gamma radiation can create free radicals in cells that are able to alter plant cellular structure and metabolism depending on the dose of irradiation. Stomatal conductance is an indicator for stomatal opening [35]. Willmer and Fricker [36] stated that abscisic acid (ABA) as a plant hormone is responsible for regulating stomatal behaviour, inhibiting the stomatal opening and inducing stomatal closure. There is some defense related phenomena that can possibly induce the initial stomatal closure [14,20].
First layer of epiderm produce epidermal structures like trichomes and stomata. No epidermal cell progeny is pushed into the core of the plant tissue as a result of divisions. Hence, any change as a result of mutations are preserved and passed down the cell lineage that forms the outer skin of the plant. Study of the size of stomata could be a suitable indicator of the magnitude of mutation as a result of exposure to physical mutagens like gamma rays. Therefore, changing of stomata aperture can potentially be used to fix gamma radiation dose levels for reliable survival of mutagenic progeny in in vitro and field systems [37].
Stomata responded quickly to environmental changes by reducing their dimensions and areas [18,38]. However, some studies have shown that abiotic stress treatments have no effect on stomatal density [18,39].
Nogués et al. [40] also observed that ultraviolet-B radiation resulted to stomatal closure in three different plants of pea (Pisum sativum L.), commelina (Commelina communis L.) and oilseed rape (Brassica napus L.). Moreover, study done by Ahuja et al. [3] reported that higher gamma irradiation doses resulted to inhibition of stomatal conductance. Stomatal movements are regulated by many abiotic and biotic factors, including radiation and the plant hormone ethylene [1,41].
Kumari et al. [42] reported that number of chloroplast decreased and shape of chloroplast showed drastically damage in gamma irradiated chrysanthemum plants. Similarly, stated that no ultrastructural changes in cell organelles occurred at gamma irradiation doses of 0 to 5 Gy. While, relatively high-dose gamma treatment (50 Gy) resulted to the alteration of chloroplast structures and disruption of organelle membranes. They also reported that some portions of the mitochondria were structurally altered.
Gamma rays caused chloroplasts to lose structural integrity. The orderly pattern of grana and stroma thylakoids was disrupted, and some of the thylakoids appeared to be slightly dilated [43]. Most of the stroma displayed crystalline appearance, which may give rise to the water stress imparted by gamma irradiation treatment [44]. Teramura et al. [45] also reported that gamma-irradiated plants suffered water stress. Application of irradiation resulted to appearance of starch granules and plastoglobuli in the stroma [10,15]. Ultrastructural configurations of chloroplasts represented the ideal system to maximize photosynthesis [15]. Excessive production of starch granules in the stroma is a reflection of depressed translocation of photo assimilates, arising from a source-sink imbalance between the capsule wall and the lint [15].
These products are known to physically prevent light from reaching the thylakoids and impede photosynthesis [46]. Hence, it appears that starch accumulation triggers events that are inhibitory to photosynthesis [15]. The accumulation of starch within the chloroplasts accompanied by damage and disorientation of grana and thylakoids indicate an inhibition of carbohydrate transport [15]. Similar results have been reported to arise from other environmental stress factors such as UV light, heavy metal contamination, acid rain and excessive light exposure [47-49] reported that their TEM analysis showed that the structure of chloroplasts was obviously altered and the membranes of cell organelles were swollen and disrupted under highdose gamma irradiation. However, low dose irradiation did not cause changes to the ultrastructural of chloroplasts. From the ultrastructural observations of the irradiated plant cells, the prominent structural changes of chloroplasts revealed that chloroplasts were more sensitive to a high dose of gamma rays than other cell organelles [50].
Afrasiab and Iqbal [2] stated that RAPD markers were efficient in discriminating somaclonal variants and induced gamma-irradiated mutants of potato. They conclude that the presence of polymorphic bands in variants and mutants suggest that genetic variation occurred in all the treatments as compared to the controls. Gamma ray irradiation resulted to DNA structural changes (breaks, transpositions, deletions) which produced band polymorphism [51].
Sianipar and Maarisit [52] reported that gamma irradiation resulted to the genetic variation in rodent tuber (Typhonium flagelliforme Lodd.) and theses genetic changes were detected by RAPD molecular markers. El Sherif et al. [30] stated about the presence of variation in DNA of the irradiated seeds of Hibiscus sabdariffa in comparison to the control were successfully assessed using random amplified polymorphic DNA-polymerase chain reaction (RAPD-PCR). Ganapathi et al. [44] highlighted the effect of gamma irradiation on banana using RAPD-DNA analysis. Each primer generated a unique set of amplification products and some revealed higher polymorphisms than others. Similarly, Minisi et al. [53] stated that RAPD analysis showed polymorphic bands when seeds of Moluccella laevis irradiated with gamma irradiation.
The results obtained in this study are similar to Wendt et al. and Hegazi and Hamideldin [48,54] who used the RAPD markers to study the effect of gamma radiation on potato and okra (Abelmoschus esculentus ) plants and concluded that treatment with highest gamma irradiation promoted the highest variation in genetic distances. Guozhong et al. [46] also pointed out that RAPD-DNA analysis showed that the offspring’s of the 5 irradiated Chamaecrista cultivars were significantly genetically different from the control treatment.
Ahmad et al. [5] reported that lower dose of ion beam irradiation resulted to the highest average PLBs shoot regeneration, while the PLBs shoots regeneration decreased at higher doses of irradiation. They concluded that PLBs irradiated at higher dose did not produce complete shoots. Similarly, Hung and Johnson [50] have observed that mutation treatment with gamma and X-ray irradiation in Wasabia japonica resulted in an alteration of the explant growth whereas treatment of both gamma and X-ray irradiation at 10 and 20 Gy had no significant differences in shoot growth while higher doses produced necrotic and non-regenerable explants.
Ahmad et al [5] indicated that higher ion beam doses increase the frequency of narrow and pointed leaves of Oncidium lanceanum plantlets producing from PLBs. Datta [23] reported that physiological disturbances and chromosome aberrations are able to increase the number of such abnormal leaves in the treated populations. Results of this study was also in accordance with [55,56] reports on the root growth of rice (Oryza sativa L. ) plant was reduced in a dose-dependent manner of irradiation. Qin et al. [57] confirmed that gamma irradiation inhibit root development among regenerated Brassica oleraceae plants.
Hewawasam et al. [49] reported that higher doses of gamma radiation resulted decrease in the average number of shoots, leaves and roots produced in Crossandra infundibuliformis var. danica after two months of treatment. Similar results have also been reported in other ornamental plants such as Chrysanthemum, Rose [23] and Tulip [55]. Vazquez-Tello et al. [55] reported that irradiation affects cell division, which is the most sensitive part to irradiation and is able to inhibit plantlets growth. According to Preuss and Britt [58], high doses of irradiation damage the genome leading to Arabidopsis growth inhibition [59-72].
Kim et al. [55] reported that gamma-irradiated plant materials show the stimulation of plant growth at low irradiation doses. The results obtained by Kiong et al. [59] indicated that survival of plants to maturity depends on the nature and extent of chromosomal damage. Increasing frequency of chromosomal damage with increasing radiation dose may be responsible for less germinability and reduction in plant growth and survival [73-80].
The results of this observation can be concluded that PLBs of Dendrobium Sonia-28 were very sensitive to gamma rays. The optimum dose of PLBs and plantlets of Dendrobium Sonia-28 was lower doses of gamma radiation (10 and 20 Gy) since the number of PLBs survival and growth rate at higher doses of gamma irradiation decreased and surface and cell organells damaged compared to control. Furthermore, our results indicated that gamma irradiation maybe affected molecular characteristics which transferred to plantlets regeneration.