Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2013) Volume 3, Issue 2
Keywords: Banana, Fungal diversity, Fusarium wilt
China is the second largest producer of bananas in the world after India, and the biggest producer of Cavendish (AAA) bananas globally. In the past two decades, the Chinese banana industry has grown from 118.253×103 ha in 1990 to approximately 413.853×103 ha in 2010. Cavendish bananas makes up 90% of production, with the rest made up of Pisang Awak (ABB) and Dajiao (AAB) bananas [1]. The total annual turnover amounts to approximately RNB 1611.5 million (US$ 230 million). Except Taiwan, production is limited to five provinces in the south: Guangdong (39.28%), Hainan (18.02%), Guangxi (19.50%), Fujian (9.23%) and Yunnan (13.97%), and all bananas are consumed locally. Since 1996, banana production in China has been significantly affected by Fusarium wilt (Panama disease), a destructive disease caused by the soil-borne fungus Fusarium oxysporum f. sp. cubense (Foc) [2].
Foc is considered the most important pathogen of banana in the world. The pathogen gained prominence when it almost destroyed the banana export trade, based on Gros Michel bananas, in Central America in the first half of the 1900s [3]. Only the introduction of Cavendish bananas in the early 1960s prevented the industry from complete collapse. Since then Fusarium wilt has been reported in all production areas except those bordering the Mediterranean, Melanesia, Somalia and some islands in the South Pacific [4]. It is, however, outbreaks of Fusarium wilt on Cavendish bananas in Asia, caused by a damaging strain of Foc, which is of particular concern to scientists and banana producers. The strain is referred to as Foc ‘Tropical’ Race (TR) 4, and can be distinguished from Foc race 1 and 2 strains which attack Gros Michel and Bluggoe cultivars, respectively, and those attacking Cavendish bananas in the subtropics following cold stress (referred to as Foc ‘Subtropical’ Race (STR) 4) [5]. To simplify the identification of strains affecting banana, Foc has been divided into 24 Vegetative Compatibility Groups (VCGs) [6]. Strains that attack Cavendish bananas in the subtropics primarily belong to VCG 0120, and those attacking Cavendish bananas in the tropics mainly belong to the VCG complex 01213/16 [4]. Phenotypic and genotypic analyses of worldwide collections of Foc suggest that Southeast Asia is the centre of origin of Foc [6,7] and that the pathogen has been introduced into new regions from there [3].
Banana plants with Fusarium wilt symptoms were first observed in Panyu, Guangzhou city by Dr. I. Buddenhagen, a plant pathologist employed by FHIA, in 1992 (Li, personal communication). The disease, however, was never confirmed and no measures were introduced to monitor outbreaks of Fusarium wilt until 1998, when approximately 14 ha of Cavendish banana plantations were affected in Guangdong province. By 2002, Fusarium wilt has been discovered in new regions and almost 1.4×104 ha of bananas was affected. The disease now occurs in all banana production areas in China, and the total area affected by Fusarium wilt has grown to 4.0×104 ha [8]. This includes plantations planted with Cavendish, Pisang Awak and Dajiao bananas.
The development of appropriate disease management strategies for Fusarium wilt of banana, and the selection of Foc-resistant banana cultivars, requires knowledge of the diversity and evolutionary history of the pathogen. VCG analysis and a variety of DNA-based tools can be used to accurately characterize Foc isolates [9]. When isolates belong to the same VCG, their hyphae anastomose and form a stable heterokaryon because of identical alleles at all of their vegetative incompatibility (vic) loci [10]. However, when isolates have different alleles at one or more of their vic loci, stable heterokaryons cannot form and such isolates will then belong to different VCGs. DNA-based techniques, such as RAPDs, AFLPs, PCR-RFLPs and multi-gene sequencing further separate Foc into two clades and a number of clonal lineages that include closely related VCGs [6,7,11,12]. In this investigation, the diversity of Foc present in the five banana-growing provinces of mainland China has been clarified by VCG analysis, molecular methods and pathogenicity testing.
Collection of isolates
Discolored vascular tissue in pseudostems of banana plants with Fusarium wilt symptoms were collected in Guangdong, Hainan, Guangxi, Fujian and Yunnan provinces of mainland China (Table 1). Dried vascular strands of each infected banana plant were then cut into 10-mm pieces, placed individually onto half-strength potato dextrose agar (½ PDA) modified with 0.1 g/l streptomycin, and incubated for 5-7 days at 25°C. Developing colonies reminiscent of Fusarium were selected, one from each diseased banana plant, and single-spore isolates prepared for identification purposes [10]. All isolates are stored at the facilities of the Department of Plant Pathology, Stellenbosch University in South Africa.
| Province | Banana variety | Total | ||
| Brazilian (AAA) | Guangfen #1 (ABB) | Dajiao (AAB) | ||
| Guangdong | 19 | 18 | 3 | 40 |
| Hainan | 2 | 12 | 0 | 14 |
| Guangxi | 6 | 1 | 0 | 7 |
| Fujian | 10 | 6 | 0 | 16 |
| Yunnan | 0 | 3 | 0 | 3 |
| Total | 37 | 40 | 3 | 80 |
Table 1: Number of Fusarium oxysporum f. sp. cubense isolates collected from three banana varieties in five Chinese provinces.
Morphological identification of fusarium isolates
All Fusarium isolates were cultured on PDA and Carnation Leaf Agar (CLA) for identification using cultural and morphological characteristics [10,13]. Cultures morphologically identified as Fusarium species other than F. oxysporum and those F. oxysporum isolates of which morphological identities were doubtful, were subjected to sequencing of the Translation Elongation Factor 1α (TEF) gene and compared to sequences in GenBank using NCBI BLAST.
DNA isolation, pcr amplification and sequencing
DNA was obtained from all Fusarium isolates after culturing on ½ PDA for 10-14 days at 25°C. Mycelia was harvested and placed in Eppendorf tubes, where after DNA was extracted by dissolving the fungal cell walls in 400 μl lysis buffer containing glass beads and shaking in a Retch MM 301 shaker (Düsseldorf, Germany) for 5 min. The suspension was incubated in a water bath (Polyscience, Niles, USA) at 65°C for 10 min. Following incubation, the Eppendorf tubes were centrifuged (Spectrafuge 24 D, Labnet International, Edison, USA) at 14000 rpm for 4 min to separate the DNA from cell remains. Extraction and purification of DNA was executed using the Wizard SV Genomic DNA Purification System Kit (Promega, Madison, USA) according to the manufacturer’s instructions. After extraction, the DNA was quantified using a Nanodrop spectrophotometer (NanoDrop, Wilmington, USA), and stored at -20°C until use.
The 650-bp region of the TEF gene and 1500-bp Intergenic Spacer (IGS) region of each Fusarium isolates were amplified on an Eppendorf Mastercycler Gradient PCR machine (Eppendorf Scientific, Hamburg, Germany) using primer sets EF1 and EF2 [11] and PNFO and PN22 [14] respectively. The TEF PCR reaction volumes was conducted in a total volume of 40 μl containing 1 x Buffer, 3.5 mM MgCl2 (Bioline, USA), 1.5U Taq DNA polymerase, 0.4 mM dNTPs (Bioline), 1 mg.ml-1 BSA and 0.4 μM of each primer (EF1 and EF2) (Whitehead Scientific, Cape Town, South Africa). For the IGS PCR, a total volume of 50 μl was used containing 0.8 x Buffer, 2.8 mM MgCl2, 3U Taq DNA polymerase, 0.26 mM dNTPs, 0.8 mg.ml-1 BSA and 0.2 μM of each primer (PNFO and PN22). PCR cycling conditions for TEF amplification included 40 cycles at 94°C for 45 sec, 53°C for 45 sec and 72°C for 60 sec. PCR conditions for the IGS gene region were set at 35 cycles at 94°C for 45 sec, 53°C for 45 sec, and 72°C for 90 sec. PCR cycles were preceded by a denaturation step at 94°C for 5 min and concluded by an extension step at 72°C for 5 min. The PCR products were purified using High Pure PCR Product purification Kit (Invetec, Hayward, USA) and sequenced in both directions with the TEF and IGS primers on an automated sequencer (Applied Biosystems, Foster City, USA) using the Big Dye™ Terminator version 3.1 cycle sequencing kit (Applied Biosystems). An additional internal reverse primer, IGS2, was included for sequencing of the entire 1 500-bp IGS fragment [7]. Raw sequences were visualized, corrected and compared to NCBI Genbank and Fusarium databases.
Vegetative compatibility group analyses
PCR-RFLP analysis of the IGS region was performed to reduce the number of pairings with tester strains during VCG analysis [7]. The IGS-PCR product was digested with five restriction enzymes (Eco88I, BceAI, Csp6I, BsrDI and BbvI) to separate them into one of eight different lineages. The enzyme Eco88I (Fermentas, Nunningen, Switzerland) separated isolates into one of two clades, called Clade A and Clade B. Clade A contains Foc race 4 isolates, and Clade B contains Foc race 1 and 2 isolates. Clade A is further subdivided into Lineages 1-5 using restriction enzymes BceAI (New England BioLabs, Hitchin, England) Csp6I (Fermentas) and BsrDI (Fermentas), and Clade B into lineages 6-8 using the restriction enzyme BbvI (New England BioLabs). In addition, the primer set R117 (GTC AAC CAG GAG CAG ACT G) and U9 (GTA ACC TCT GAC TCA CCG) was used to target a repeated region on the Foc mitochondrion (MtR) that distinguish between Lineage VII and VIII isolates within Clade B [7].
Fusarium oxysporum isolates were subjected to VCG testing according to the method described by Leslie and Summerell [10], with minor modifications. Briefly, all Chinese cultures were grown on ½ PDA at 25°C for 1 week, where after aerial hyphae were collected with a sterile needle and placed on a 90-mm-diameter Petri dish containing 15% Chlorate Medium (CM), at four hyphael collections per plate. The cultures were incubated under lights at 25°C and inspected daily for the development of fast-growing sectors. These sectors were then transferred to Minimal Medium (MM) slants and incubated at room temperature for 14 days. All cultures that produced sparse-growing hyphae were considered nit-mutants, while those that reverted to wild type growth were discarded. The nit-mutants were then typed on a selective nitrogen-modified medium as either nit-1, nit-3, nit-M or crn type mutants [10].
Nit-1 and/or nit-3 mutants representing each of the Chinese isolates were crossed with nit-M VCG tester strains that represent the known VCGs of Foc. These testers were kindly provided by Drs S. Bentley and W. O’Neal from QDPI, Australia, and Prof. R.C Ploetz from the University of Florida, USA. Nit-testers of VCGs 01221, 01222, 01223 and 01224 were generated at Stellenbosch University after receiving cultures representing these isolates from the USDA-ARS in Peoria, USA. During the pairing of unknown nit-1 mutants with nit-M testers, a known nit-1 tester was included as control. The Chinese mutants were only paired with tester strains belonging to the lineage that they were grouped into previously, as described above. If the Chinese mutant did not pair with any of the known VCG testers, they were paired with a nit-M mutant generated from the same isolate to exclude the possibility of vegetative self-incompatibility, and with all the other nit-M testers to ensure that they were not incorrectly typed to a specific lineage. All isolates were tested at least twice for VCG identity.
Identification of Foc TR4
To rapidly detect isolates of Foc TR4 (VCG 01213/16) in the Chinese population of Foc, a VCG 01213/16-specific primer set was developed. Sequences of both IGS and TEF-1a regions were manually edited using Geneious® bioinformatics software version 6.0.6 (Biomatters Ltd, Auckland New Zealand) and aligned to datasets, including representatives of all known VCGs and closely related formae speciales of F. oxysporum, using MAFFT version 7, with the L-INS-i option effective and visualized in Se-Al version 2 (Oxford, England). Sequences of the IGS region representing different VCGs were obtained from NCBI Genbank. Single Nucleotide Polymorphisms (SNPs) were identified within the IGS region and used for primer design. One forward (VCG01213/16 F1) and one reverse primer (VCG01213/16 R2) were designed to generate a unique amplicon of 455 bp (Table 3). The PCR assay was conducted using 20 ng of fungal DNA in a total volume of 25 μl containing 2 mM MgCl2, 1 U Taq DNA polymerase, 0.32 mM dNTPs, 0.8 mg.ml-1 BSA and 0.2 μM of each primer. Cycling conditions consisted of 35 cycles at 94°C for 45 sec, 64°C for 45 sec and 72°C for 60 sec. Each PCR was preceded by initial denaturation step of 94°C for 5 min and concluded with a final extension step of 72°C for 5 min. Specificity was confirmed by testing the primer set against all Chinese isolates included in the VCG analysis, a collection of 87 isolates that included representatives of all VCGs of Foc, other formae speciales of F. oxysporum (chrysanthemi, conglutinans, dianthi, elaeidis, gladioli, lini, lycopersici, melonis, nicotianae, niveum, phaseoli, passiflorae, perniciousum, pisi, radicis-lycopersici, raphani and tulipae), and non-pathogenic isolates of F. oxysporum.
Pathogenicity testing
To determine cultivar specificity and compare the virulence of individual isolates, three banana varieties were inoculated with the Chinese F. oxysporum isolates under greenhouse conditions. In addition, Fusarium species other than F. oxysporum that were isolated from Chinese banana pseudostems were also included in the pathogenicity tests. The cultures were first grown in Armstrong medium [16] at 25°C for 4 days to ensure ample production of microconidia. A spore suspension of 1×106 spores/ml was prepared after the spores were washed with sterile distilled water, followed by filtration through cheese cloth and counting with a haemacytometer. The fungal isolates were then added to a planting medium in 2 liter pots that consisted of three parts vermiculite, one part peat, and 0.5 parts coconut coir, in order to give a final concentration of 5000 conidia/g soil. Sixty plants each of the Cavendish cv. Brazilian, the Pisang Awak cv. Guangfen #1, and the Dongguan cv. Dajiao were used to evaluate each Fusarium isolate. Disease incidence was determined 4 weeks after inoculation as a percentage of plants that developed internal and external Fusarium wilt symptoms [17].
Identification of isolates
Fusarium oxysporum isolates were distinguished from other Fusarium species by the ample production of microconidia on short monophialides, and by the production of chlamydospores that were single or in pairs, on CLA. Of the Chinese isolates, 80 were identified as F. oxysporum, four as F. proliferatum and two as F. commune. Molecular analysis confirmed the morphological identification. Fusarium commune was isolated from Dajiao bananas in Guangdong province. The Dajiao bananas in Guangdong province also contained an isolate of F. proliferatum, while the same species was further isolated from Guangfen #1 bananas in Guangdong and Yunnan provinces.
Fifty percent of Foc isolates collected in China was from Guangdong province (Figure 1). These isolates were isolated from Brazilian, Guangfen #1 and Dajiao bananas (Table 1). In Hainan and Fujian provinces, 14 and 16 isolates, respectively, were collected from diseased banana pseudostems of Brazilian and Guangfen#1 bananas only (Table 1). Foc isolates were collected from Guangfen #1 in all five provinces and from Brazilian bananas in four provinces. Only three Foc isolates were obtained from Dajiao bananas, in addition to F. proliferatum and F. commune (data not presented), and these all come from Guangdong province.
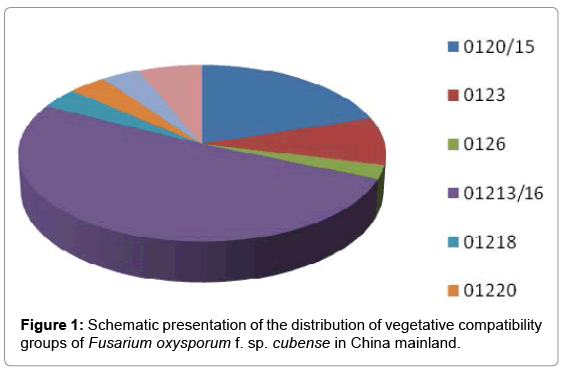
Figure 1: Schematic presentation of the distribution of vegetative compatibility groups of Fusarium oxysporum f. sp. cubense in China mainland.
Vegetative compatibility groups
The Chinese Foc isolates were divided into five Lineages and 11 VCGs (Table 2). These include Lineages II, IV, V, VI and VII, those VCGs belonging to VCGs 0123, 0126, 01218, 01220 and 01221, and the VCG complexes 0120/15, 0124/22 and 01213/16. VCG 01213/16 (Lineage V) contained 51.25% of isolates, whereas VCG 0120/15 (Lineages IV) contained 20% of the isolates (Figure 1). Both VCG 01213/16 and VCG 0120/15 are members of Foc race 4, which causes disease to Cavendish bananas [7]. VCGs 0123 and 01218 are members of Lineage VII, which primarily infect non-Cavendish varieties, and are therefore designated Foc race 1. VCG 01213/16 isolates were found in all provinces except Yunnan province, while VCG 0120/15 isolates were found in Guangdong, Yunnan and Fujian provinces (Table 2). Lineage VII isolates (VCGs 0123 and 01218) were found in Guangdong, Hainan and Guangxi provinces and Lineage VI isolates (VCGs 0124/22 and 01220) occurred in Guangdong and Hainan provinces. VCG 0126 (Lineage II) was found in Guangdong and Yunnan provinces only (Figure 2).
| VCG | Lineage designation | Number of isolates | Isolated from | Disease incidence (%) | ||
| Brazilian (AAA) | Guangfen #1 (ABB) | Dajiao (AAB) | ||||
| 0126 | II | 2 | Guangdong | 8.5 (5-12) |
66 (63-69) |
0.0 |
| 0120/15 | IV | 14 | Fujian; Guangdong; Yunnan | 46.7 (13-72) |
63.1 (37-85) |
0.0 |
| 01213/16 | V | 40 | Fujian; Guangdong; Guangxi; Hainan | 66.2 (39-78) |
68.8 (49-87) |
0.0 |
| 0124/22 | VI | 3 | Guangdong; Hainan | 6.3 (3-11) |
74 (59-82) |
0.0 |
| 01220 | VI | 3 | Guangdong; Fujian | 12.3 (6-18) |
60.7 (59-62) |
0.0 |
| 0123 | VII | 7 | Guangdong; Guangxi; Hainan | 17.9 (6-56) |
75 (62-88) |
0.0 |
| 01218 | VII | 3 | Guangxi | 14.0 (11-18) |
82.3 (79-86) |
93.6 (92-96) |
| 01221 | Unknown | 5 | Guangxi; Hainan; Yunnan | 12.4 (6-22) |
70.2 (53-85) |
0.0 |
Table 2: The average incidence of Fusarium wilt developing in three banana varieties grown in China following greenhouse inoculation with Fusarium oxysporum f. sp. cubense.
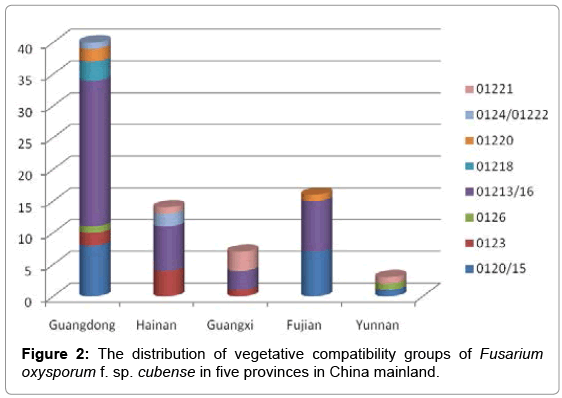
Figure 2: The distribution of vegetative compatibility groups of Fusarium oxysporum f. sp. cubense in five provinces in China mainland.
Guangdong Province had the greatest number of VCGs (9) in China, followed by Hainan (6), Fujian (5), Guangxi (4) and Yunnan (4) provinces (Figure 2). More isolates were collected in Guangdong than in any of the other provinces (Table 1) as this province represents the largest production area in China. The diversity of the fungus in Hainan was particularly high, with six VCGs present in the 14 isolates collected. Isolates belonging to the VCG 01213/16 complex were collected in all provinces, with the exception of Yunnan, and those belonging to the VCG 0120/15 complex, VCG 0123 and VCG 01221 were all collected from three provinces. VCG 0126 was unique to Guangdong province.
Six VCGs were isolated from Brazilian bananas, a Cavendish variety grown widely in all banana-producing provinces in China (Table 1) ( Figure 3). These include VCGs 0120/15, 0123, 01213/16 and 01221. Most VCGs, however, were collected from Guangfen #1, a Pisang Awak banana which is susceptible to all Foc races, namely VCG 0120/15, 0123, 0124/22, 0126, 01213/16, 01220 and 01221.
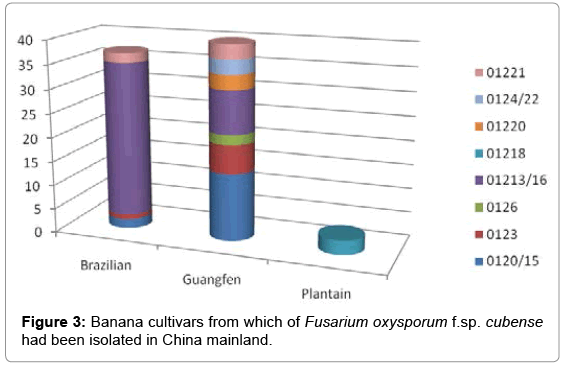
Figure 3: Banana cultivars from which of Fusarium oxysporum f.sp. cubense had been isolated in China mainland.
Specificity of Foc TR4 primers
The primers set designed in this study (VCG01213/16 F1 and VCG01213/16 R2) were specific for VCGs 01213, 01216 and 01213/16, and produced amplification products of 455 bp (Figure 4). Electrophoresis of PCR products did not show bands of any other known Foc VCG, non-pathogenic F. oxysporum, or formae specialis of F. oxysporum. It successfully identified only isolates characterized by VCGs as members of the VCG 01213/16 complex in the Chinese Foc population.
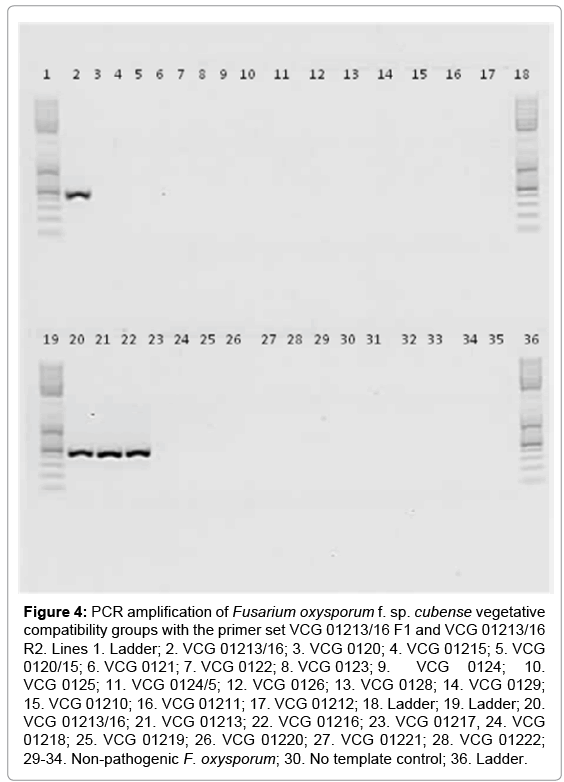
Figure 4: PCR amplification of Fusarium oxysporum f. sp. cubense vegetative compatibility groups with the primer set VCG 01213/16 F1 and VCG 01213/16 R2. Lines 1. Ladder; 2. VCG 01213/16; 3. VCG 0120; 4. VCG 01215; 5. VCG 0120/15; 6. VCG 0121; 7. VCG 0122; 8. VCG 0123; 9. VCG 0124; 10. VCG 0125; 11. VCG 0124/5; 12. VCG 0126; 13. VCG 0128; 14. VCG 0129; 15. VCG 01210; 16. VCG 01211; 17. VCG 01212; 18. Ladder; 19. Ladder; 20. VCG 01213/16; 21. VCG 01213; 22. VCG 01216; 23. VCG 01217, 24. VCG 01218; 25. VCG 01219; 26. VCG 01220; 27. VCG 01221; 28. VCG 01222; 29-34. Non-pathogenic F. oxysporum; 30. No template control; 36. Ladder.
Pathogenicity testing
Isolates from Dajiao bananas caused Fusarium wilt to both Dajiao and Guangfen #1 banana plantlets during greenhouse pathogenicity testing (Table 2). Disease development on Dajiao, however, was twice as fast on Guangfen #1 bananas. In Dajiao, leaves turned yellow approximately 20 days after inoculation, while symptoms became visible on Guangfen #1 in 40 days after inoculation. Dajiao bananas were immune to all isolates collected from Cavendish and Guangfen #1 bananas, which were identified as known Foc VCGs. Foc VCG 01213/16 and 0120/15 caused disease to both Cavendish and Guangfen #1 bananas, whereas isolates in VCG 0124/22 caused disease mainly in Guangfen #1 bananas.
Since its discovery in Indonesia and Malaysia in the early 1990s, Foc TR4 has been reported from and caused destruction of Cavendish plantations in Indonesia, Malaysia, Taiwan, the Northern Territory of Australia, southern China and the island of Mindanao in the Philippines [4]. In these areas, Foc races and VCGs responsible for such disease outbreaks have to be rapidly and accurately identified, and banana varieties tested and improved for resistance to all forms of the pathogen. The rapid and accurate detection of plant pathogens is also of great importance for implementing quarantine measures between and within countries, and to manage banana Fusarium wilt outbreaks using appropriate banana varieties and hybrids. The current study represents the first report of the diversity and distribution of Foc in China.
Foc appears to be well established in China. The population is highly diverse, with the greatest diversity occurring in Guangdong province. Guangdong is in the southern subtropics, and has the largest area planted to bananas in China. The main VCG associated with bananas in the province was VCG 01213/16, known as tropical form of Foc race 4. However, it is the widespread occurrence of VCG 0120/15 in the province, often referred to as the ‘subtropical’ form of Foc race 4, which was of particular interest. This VCG is commonly found in sub-tropical banana-producing countries such as Australia, South Africa and the Canary Islands, but seldom in the tropics. VCG 0120 has also been found in Fujian and Yunnan provinces, but not in Hainan province. The reason might perhaps be due to the climate of Hainan province, which is tropical whereas the climate of the other provinces is subtropical. The proper mapping of Foc VCGs occurring in China is important to control the movement of plant material, for implementing quarantine measures, and for the employment of Fusarium wilt-resistant banana varieties.
The rapid spread of Foc in China has decreased production of Cavendish and Pisang Awak bananas in many areas. In these areas, the cultivation of Dajiao became more frequent, despite its relative poor quality compared to Cavendish and Pisang Awak. The disease in Dajiao bananas was first observed in two banana-growing areas in Southern China, Dongguan and Panyu, in 2007 and in 2012, respectively. Now, an estimated 5% of all losses of banana in local area were attributed to Fusarium wilt of Dajiao. All three Foc isolates from Dajiao belong to VCG 01218; a VCG that was previously associated with both AAB and ABB bananas in Indonesia, Malaysia and Thailand [4]. In China, however, it was neither isolated nor caused disease to Guangfen #1, a Pisang Awak cultivar.
Banana varieties grown in China appear to be differently affected by Foc VCGs. For instance, all Chinese VCGs were found on Guangfen #1, and this variety was also susceptible to all VCGs during pathogenicity testing. Brazilian bananas were more susceptible to VCGs 0120/15 and 01213/16 than to VCGs 0123, 0124/22 01220 in the greenhouse, thereby confirming its Foc race 4 categories. The Foc race 4 VCGs also affected Pisang Awak bananas, thereby indicating its wide host range. Interestingly, the highly virulent tropical strains of Foc race 4 (VCG 01213/16) did not cause disease to Dajiao, a plantain-type banana. This explains why Dajiao can be planted in Foc race 4-affected fields in China. It is also important to test the susceptibility of Cavendish somaclonal variants, such as GCTCV 119 and 218, known to be resistant to VCGs 01213/16 [18], against VCG 01218 under field conditions. Cavendish banana varieties should also be tested against VCGs 0124/22, 01220 and 01221 at infection sites in China in order to employ resistant banana varieties for Fusarium wilt control. Although supposedly resistant to Foc race 1, Cavendish bananas have not been evaluated in the field against members of VCG 01220 and 01222, and have succumbed to strains of VCG 0124 in India [19].
Two other Fusarium species have been isolated from banana pseudostems in China, including F. proliferatum and F. commune. Fusarium proliferatum is a common endophyte of banana roots, and have previously been found associated with banana rhizomes and fruit [20]. However, it is not known to contribute to Fusarium wilt of banana. Fusarium commune is a newly described species that can easily be confused with F. oxysporum based on morphological identification [21]. It differs from the latter in that it produces microconidia in false heads on both mono- and polyphilalides. Neither of the two species proved to be pathogenic to banana seedlings, and is considered nonpathogenic colonizers. It would also be interesting to determine whether F. commune is not more commonly associated with banana, and that previously identified non-pathogenic strains of F. oxysporum did not represent F. commune.
An Foc TR4-specific primer set previously developed by Dita et al. [15] is currently the only one available for PCR diagnostics of VCG 01213/16. Because of the threat of Foc TR4 to banana production internationally, a second VCG 01213/16-specific primer set was developed in the current study for use in combination with the Foc TR4 marker in diagnostic centers and laboratories .The simultaneous use of both markers for Foc TR4 diagnostics would significantly enhance accuracy and ensure that false positive or negative results based on PCR analysis only do not occur (Table 3).
| Primer name | Primer sequence |
|---|---|
| 01213/16 F1 | ACG TTT AAG GTG CCA TGA GAG |
| 01213/16 R2 | CCT CGT GAG CCA CTT TTT AT |
Table 3: The primers specific for 01213/16 showing sequence information and amplicon size.
Fusarium wilt is spreading at a rapid pace in China. Foc is wellknown to be introduced into wilt-free banana fields in infected planting material, from where it is disseminated in water and soil, on tools and equipment, and by the movement of people within and between fields [3]. Banana farming in China consists primarily of small fields of between 0.5 and 2.0 ha. The farmers have a limited knowledge of banana Fusarium wilt, and often use suckers obtained from other farms for replanting instead of disease-free tissue culture plantlets. There is also a significant movement of people and vehicles within and between farms, all potentially contributing to the disease. Once introduced, the fungus will survive in fields for decades, and will severely affect all subsequent plantings of susceptible banana varieties [3]. It is, therefore, of significance that awareness of the threat of Fusarium wilt be raised among producers in China. In areas where the disease has been established, the impact of the disease could potentially be reduced by rotation with leek [2] and, in future, the planting of resistant somaclonal variants of Cavendish and Pisang Awak. The latter is currently being developed through a comprehensive banana improvement programme at GDAAS.
This work was supported by National Natural Science Foundation of China No. U1131004 and 30971991, National High-tech R&D Program (863 Program) No. 2011AA10020602, 948 Project of Ministry of Agriculture No. 2011-G16, National Banana Industry and Technology System Project No. CARS-32-01, International Collaborative Project of Ministry of Science and Technology No. 2013DFB30400 and No 20110501, Commonwealth Industry (Agriculture) Specific Fund No. 200903049-10 and the South African Biodiversity Initiative (SABI). We are grateful for technical assistance provided by Madelein du Plessis.