Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Review Article - (2017) Volume 5, Issue 4
Cytochrome P450s belong to a particular class of enzymes (Oxygenases) which are extensively distributed in all classes of organisms that attract the interest of scientists worldwide. It has been proved multi-functioned super gene family that sophisticates the biosynthesis of several endogenous molecules and metabolism of discrete pharmacologically important oxymolecules such as Antibiotics, essential secondary metabolites, fatty acid conjugates, signaling molecules, lipid degradation, hormones and many more. In this article, we briefly overviewed the heterogeneous role of the superfamily of Cytochrome P450 based on recent advances in molecular biology and genetic engineering. The inevitable role of Cytochrome P450s proteins in the biosynthesis of secondary metabolites likewise Hormones, Flavonoids, signaling molecules and other important pigments in plants such as anthocyanin, terpenoids and their pharmacological significance are specifically focused. We have predicted the distinctive metabolic networks and molecular characteristics of Safflower genome based on extensive transcriptome analysis from various developmental stages of floral tissues. The presence of repeated sequences, high copy number coding and noncoding RNA sequences and high expression level in the petals provide a gateway to enable the development of all-inclusive gene networks for economic and clinical features of Cytochrome P450 family. The implementation of metabolic engineering in floral pigments and alteration in their biosynthetic pathways can be exploited for a comprehensive understanding of several other pathways which invites new avenues for novel therapeutic blueprints and drug development
Keywords: Cytochrome P450; Expression; Flavonoid; Application; Plants
Enzymes are generally proteins and often provide a favorable environment for discrete biochemical reactions occurred in cellular pathways which plays a significant role in transforming substrates under controlled reaction conditions (room temperature, pH minimum activation energy) unlike general insensitive biochemical reactions [1]. Oxygenases are a particular class of enzymes that hold a prominent role in direct hydroxylation of aromatic or aliphatic carbonhydrogen bonds. The major classes of oxygenase that catalyze hydroxylation reactions include flavin-dependent monooxygenases, peroxidases, and Cytochrome P450s, which include heme as a cofactor and therefore known as hemoproteins.
Cytochromes P450 are found in all forms of life from prokaryotes to eukaryotes including humans, plants, insects, fungi and bacteria, and have even been observed in viruses [2]. To date, approximately 15,000 Cytochrome P450 genes have been recognized in various kingdoms, among which a third of common sequences are contributed by kingdom Plantae . The contribution of Cytochrome P450 system in flowering plants (Angiospermae ) genome is approximated as >1%. On the other hand, prokaryotic genome also accounts the assembly of these unique Cytochrome P450 systems with few exceptions in case of Gram-negative Escherichia coli , which are deficient in Cytochrome P450 system.
Miller and their disciple Mueller founded in vitro investigation of rat liver and thus understand the involvement of these enzymes for the irst time speci ically during the degradation of the carcinogen 4- dimethylaminoazobenzene in 1949 [3]. Ryan and Engel for the irst time discovered P450 in rat liver microsomes and observed the reversible inhibition of CO by light in 1958 which lead to the later discovery of the function of Cytochrome P450s, carrying a pivotal role in the form of oxidase which utilizes molecular O2 and NADPH during biochemical reshaping that occurs in various signi icant pathways [4]. Advances in steroid hormone biosynthesis conjointly subsidized towards the origin of Cytochrome P450s complex, it was found that these group of complex enzymes also intricate in the hydroxylation of C21 of progesterone from the bovine adrenal cortex in microsomes [5]. A major breakthrough made by Martin Klingenberg in 1958 in the Cytochrome P450 groundwork was the ferreting out of a shi t in the spectra of rat liver microsome from 413 nm to 450 nm during the spectral investigation of a novel pigment bound with CO (carbon monoxide) [6].
Cytochrome P450 enzymes exhibit broad diversity with respect to their biological occurrence and biochemical functions and properties, such as location in cells or tissues, solubility, types of redox partners, catalyzed substrates or chemical transformations. However, all of them share similar structural features and functional properties. All Cytochromes P450 apparently originate from a single ancestor CYP51 [7] and thus exhibit a common pattern of protein fold. Furthermore, all P450s contain the same heme prosthetic group (Fe-protoporphyrin IX) with cysteine ligating heme iron [8] and all of them utilize a similar catalytic cycle that activates the molecular oxygen or hydrogen peroxide. These analogies in structure and catalysis provide an opportunity for extrapolation of information obtained from some isozymes to the studies of others, as well as for making certain predictions about functional properties of previously unknown P450 enzymes.
It has been observed so far that the majority of the members of Cytochrome P450 enzymes are covered by plant kingdom classified into different gene families, subfamilies, and isoforms [9]. Individual P450s are uniquely identified by the last number in the CYP name [10]. These Cytochrome P450s can facilitate distinctive enzymatic reactions such as metabolism of aromatic and aliphatic molecules which undergo hydroxylation, N-oxide reduction dealkylation, desulphuration, sulfoxidation, deamination, epoxidation and dehalogenation and variety of reactions [11].
Natural metabolites for example Flavonoids, unsaturated fatty acids and hormones are commonly recognized at very low concentrations in combine plant extracts. Moreover, these substances are synthesized normally at various stages of particular tissues and more often they were founded as lineage-specific in various plants which falls under narrow taxonomic groups. These factors greatly influence the extraction and purification of these remarkably beneficial substances on a wider front. Carthamus tinctorius is exceptionally acknowledged for its seed oil consists of high amount of unsaturated fatty acids which is extremely rich in linoleic and oleic acid [12,13]. Various candidate genes from Cytochrome P450 enzymes family in C. tinctorius have been revealed by our esteemed group which is solely responsible for Flavonoids accumulation. However, there are many branch points still need to be unpacked with the help of metabolic engineering and high throughput technology in order to fully understand the metabolic flux, regulation and hereditary characteristics of essential flavonoids and polysaccharides for significant therapeutic targets. This can be followthrough by introducing artificial switch in the metabolic network of floral pigments in various flowering plants.
The Supremacy and Functional Diversity of Cytochrome P450 System
Cytochrome super gene family consisting of more than 4000 identified members carrying below 20% sequence similarity, despite this fact, their anatomic organization holds a strongly conserved topography and 3D folding. In the eukaryotic cellular system, Cytochrome P450 enzymes are present in membrane-bound organelles such as endoplasmic reticulum, plastids, Golgi complexes and mitochondria [14]. However, some P450s have been reported in the external chloroplastic membrane, for example, Cytochrome P450 enzymes in Arabidopsis [15] while in prokaryotes they are soluble in the cytoplasm.
P450 systems across all kingdoms catalyze a huge diversity of different region and stereospecific irreversible biochemical reactions during their natural metabolic routes, the general function of Cytochrome P450s can be identified from their interaction with various kinds of substrates, which are mostly hydrophobic organic compounds. In view of their substrates, Cytochrome P450s have been proved multi-functioned super gene family that sophisticates the biosynthesis of several endogenous molecules and metabolism of discrete pharmacologically important drugs, heterogeneous xenobiotics, cosmetics and polymer industries. Cytochrome P450s in prokaryotes participates in antibiotics metabolism, the degradation of particular carbon-containing compounds and during biosynthesis of fatty acids molecules [16]. Eukaryotic Cytochrome P450s adversely accounts for the metabolism of membrane sterol. For example, in animals, these enzymes are involved in the formation of essential signal molecules along with the metabolism of steroid hormones in the steroidogenic organs and drug metabolism in the liver as well as vitamin D3 accumulation. Cytochrome P450 system in kingdom fungi also effectuates essential their part in mycotoxins and lipid metabolism [17]. In kingdom Plantae , they participate in the catabolism and synthesis of hormones and other medically important secondary metabolites.
The most important biological attribute of Cytochrome P450 enzymes is to render the necessity of NADPH cofactor with the help of heme prosthetic group during several metabolic pathways metabolism. The ability to use host cell’s cofactor restoration system can avoid the excessive amounts of cofactor require for a biochemical reaction, which would be an ultra-economic drawback in any other way. They can produce different kinds of products in the result of hydroxylation and oxidation, following by alkylation, dealkylation, epoxidation, demethylation, aryl migration and so on. These specific events are frequently attributed to P450 systems [18]. Various plant Cytochrome P450s play a key role in the synthesis of metabolons in association with additional proteins. A metabolon is a non-covalent complex of various enzymes which can convert direct substrate to desired product unlike other enzymes requiring a huge amount of intermediates [19]. The Cytochrome P450 superfamily has remarkable capability in terms of useful applications in various fields such as bioconversion processes (CYP105A3), Bioremediation (CYP1 family) and Biosensors (CYP152A1) because Cytochrome P450s necessitate neither reduced nicotinamide adenine dinucleotide phosphate nor electron donor proteins, and effortlessly catalyze active metabolic events using H2O2 (hydrogen peroxide) [20]. The explicit competency of Cytochrome P450s to catalyze the oxidation of complex set of regio, chemo and stereospecific substrates lift up their profile at the molecular level and thus make them essential competitors for the upcoming biotechnological applications.
Plant metabolic systems use a range of Cytochrome P450 monooxygenases in their biosynthetic and detoxification pathways in different tissues. These enzymes are typically heme-dependent, having mixed functions as oxidases which manipulate NADPH or NADH and molecular O2 to produce functionally active organic compounds. Other catabolic P450s metabolize lethal herbicides and insecticides into non-fatal products or, conversely, biologically activate non-toxic compounds into toxic products.
A variety of cloning and transient expression strategies have already been marked useful for Cytochrome P450s research in plants [21]. This has encouraged plant scientists to isolate and characterize a number of cDNAs and gene sequences of Cytochrome P450s because many of their genes have been cloned recently. Earlier studies revealed current advancement in targeted re-sequencing which explains rare mutations and genetic polymorphism in Cytochrome P450 family showing enrich practical modulations. However, the substrate specificity of P450- dependent reactions in plants and the mechanisms by which these genes are expressed and regulated still need to dig into more depth for potential significant products. In recent years, research on the Cytochrome 450 family has extended from basic research towards the engineering of biocatalysts for a wide range of functional variations [22].
From the time of their occurrence, Cytochrome P450s captivated researchers from all over the world including biochemists, microbiologists, plant biologists, biotechnologists, and pharmacologists because of its substantial part in the biosynthesis of plant secondary metabolites, unique chemical mechanism of the Cytochrome P450 enzymes and their outstanding capability in drug degradation. Arabidopsis Cytochrome P450 genes have been identified carrying well-known functions including biosynthesis of secondary metabolites such as color pigments, hormones, defense compounds against various pathogens, flavors and complex macromolecules (lignin, suberin, and cutin) [15]. The investigation of Cytochrome P450 enzymes CYP710A1 and CYP710A11 from Arabidopsis and tomato, respectively, both in vitro and in planta simplified that they catalyzed a C-22 desaturation reaction with ß-sitosterol (substrate) yielding stigmasterol (product) while CYP710A2 is essential for the production of brassicasterol and stigmasterol from 24-epi-campesterol and ß-sitosterol, respectively. T-DNA mutagenesis analysis verified that CYP710A2 was necessary for brassicasterol formation in Arabidopsis , signifying that unknown desaturases were not capable of performing the C-22 desaturase reaction catalyzed by CYP710A2 [23]. It is possible that the desaturation reactions in other plant species could be catalyzed by other CYP710A family members because of the conserved amino acid sequences coming from single ancestral gene family hence providing a remarkable opportunity to discover a greater number of different biocatalysts [24].
A further extensive research in the field of novel anticancer drugs remains the point of interest for many toxicologists all over the world. It is postulated that most of the anticancer drugs emanate from plants sources and therefore their open accessibility is very limited, new strategies need to be adopted under the principle of recombinant expression system of candidate enzymes. In recent past, comprehensive studies were conducted using Yeast-based co-expression system in which CYP725A1 (taxoid 10b-hydroxylase) fused with another corresponding Cytochrome P450. It has been described that both of the Cytochrome P450s were isolated from Taxus cuspidate . The in vivo system of both this fusion system catalyzes the biosynthesis of anticancer drug taxol, which is recommended in the UK and many other countries for a number of types of cancer including prostate, melanoma, ovarian, breast, and pancreatic cancer [25]. Another study revealed biocatalytic production of perillyl alcohol which has also been proven an efficient anticancer drug. This whole process uses a candidate enzyme of the CYP153 family (cytochrome P450 alkane Hydroxylase) from Mycobacterium which is expressed in Pseudomonas putida yielding a sufficient amount of perillyl recovered via whole-cell process [26].
The plant Cytochrome P450 CYP71AV1 has also been proved as dynamic contributor to produce the precursor of an antimalarial drug (artemisinic acid) using recombinant expression system of fungi (Saccharomyces cerevisiae) [27]. Artemisinic acid was isolated initially from Artemisia annua spp. Artemisinin is an epoxy sesquiterpene lactone that is produced in many steps: synthesis of farnesyl pyrophosphate (FPP) originates from acetyl CoA via the MVA pathway and amorpha-4,11-diene synthase (ADS) catalyzes the conversion of FPP to amorpha-4,11-diene. Amorpha-4,11-diene is then hydroxylated to artemisinic acid by a Cytochrome P450 hydroxylase and further catalytic reactions occur to generate the final product [28]. Cytochrome P450s covers a potential class of enzymes with unique diversity to produce effective and useful products both in synthetic and pharmaceutical industries. Their complex substrate spectrum, vast substrate-binding pockets, and ability to catalyze a variety of reactions has made them amenable towards industrial applications. However, for the comprehensive industrial uses, the stability of most Cytochrome P450s are inadequate and more extensive efforts need to be made to epitomize these limitations that can amplify all directions of industrial application of Cytochrome P450s biocatalysts.
Cytochrome P450 enzymes system is an exceptional transition to an active functional class of monooxygenases participating assiduously in the biosynthesis of essential compounds such as hormones, terpenoids, xenobiotics, signaling molecules, alkaloids, glycosides, flavonoids and other natural substances [29]. Plant secondary metabolites are the most abundant organic compounds that do not directly participate in the usual growth, development, and reproduction of an organism, unlike primary metabolites although secondary metabolites are exclusively involved in the defense mechanism of various plant species against a number of pathogens that can effectively invoke harmful agents which are dangerous to human health. Plant Cytochrome P450 enzymes follow ubiquitous pathways and associates wherein resulting in the synthesis of pharmacologically distinguished compounds.
The second and most important feature of Cytochrome P450 is to render the metabolic detoxification and biotransformation of challenging exogenous compounds. Keeping in view this fact it has been remaining a great confrontation of particular importance among scientists to distinguish the sources and to probe the factors that genuinely contributes to the extraordinary pharmacological and pharmaco-dynamic variability of Cytochrome P450s enzymes within and intraspecies for novel therapeutic drugs [30]. Majority of Cytochrome P450s have been found applicable for clinical pharmacology which can catalyze the oxidative biotransformation of many drugs and lipophilic xenobiotics by switching certain metabolic rearrangements in their pathways [31]. Human Cytochrome P450s were also reported with specific intrinsic functions like the production of steroid hormones, bile acids, prostaglandins and many others [32].
The mechanism of (CYP2C19) enzyme was previously shown for the deactivation of proton pumps inhibitors (PPI) metabolism such as pantoprazole and omeprazole. It is also considered a key enzyme for the metabolic stimulation of anticoagulant clopidogrel into 2-oxo metabolites [33]. This enzyme also plays a prominent role in the biosynthesis of many antidepressants (both first and second generation) [34]. Recent advances in the field of biochemistry and plant molecular biology has to lead us towards the understanding of complex pathways carrying terpenoid biosynthesis and it is concluded that Cytochrome P450s have the potential in commencing a key role in the production of terpenoids. TIAs (Terpeneindole alkaloids) are the well-known class of secondary metabolites withholding the terpene ring and indole chain alkaloid substances.
Catharanthus roseus has been extensively studied for terpenoid indole alkaloids and their metabolite engineering research revealed the existence of anti-cancer or anti-arrhythmic effects [35]. Approximately 130 types of terpenoid indole alkaloids have been identified so far. The iridoid pathway in Catharanthus roseus starts with the activation of 10- hydroxy-geraniol which is proved lately as CYP76B6 via catalytic dehydrogenation of geraniol-10-dehydrogenase, which then modified into 10-oxogeranial with the help of redox of 10-hydroxy-geraniol and undergoes further into methyl marcfortine vomica glycosides and finally the production of secologanin through secologanin synthase. secologanin is an important antitumeric agent [36]. The authors tend to assume that this CYP76B6 itself or likely sequences might be implicated for the biosynthesis of indole alkaloids in other plant species based on the fact of complex proteins interaction with this unique class of monooxygenases. The potency of these molecular cassettes not only limited to Catharanthus roseus , but it may also be determined and investigate in several other plants species which can bring about the unaccustomed pharmacological importance in near future.
Chicory plant (Cichorium intybus L. ) is a robust herbaceous plant belongs to Asteraceae family. Chicory is considered a major source of polysaccharide (Inulin) and has been proved to acquire a broad variety of pharmacological properties containing anti-inflammatory, antidiabetic and anti-malarial effects [37]. Moreover, a direct study has been undertaken in order to investigate the consequences of specific plant secondary metabolites along with a global extract of the roots of Chicory plant upon the expression of various Cytochrome P450s (CYP1A1, CYP2A19, CYP2C33, CYP2D25, CYP2E1, and CYP3A29) in the pig hepatocytes in vitro [38]. Previous studies conducted on pigs metabolizing enzymes established the extensive involvement of CYP cassettes in detoxification of certain xenobiotics (antibiotics and drugs). It has been proved that the expression as well as the activity of these metabolizing enzymes in pigs notably enhanced with the use of chicory plant in their feed [39]. Plenty of secondary metabolites are produced by Chicory root including esculetin, artemisinin, sesquiterpene lactone lactucin and coumarins esculin [40]. Out of all tested plant secondary metabolites esculetin, scoparone, artemisinin, and lactucin all simultaneously increased the expression of aforementioned CYPs in comparison with standard activators of the receptors, however, esculin indicates no effect in the expression of these selected CYPs. These results indicate a roadmap towards the understanding of roles and challenges of plant secondary metabolites by inducing the expression of Cytochrome P450s enzymes with pharmacokinetic relevance in other species which could mimic the detoxification of xenobiotics generally (Figure 1).
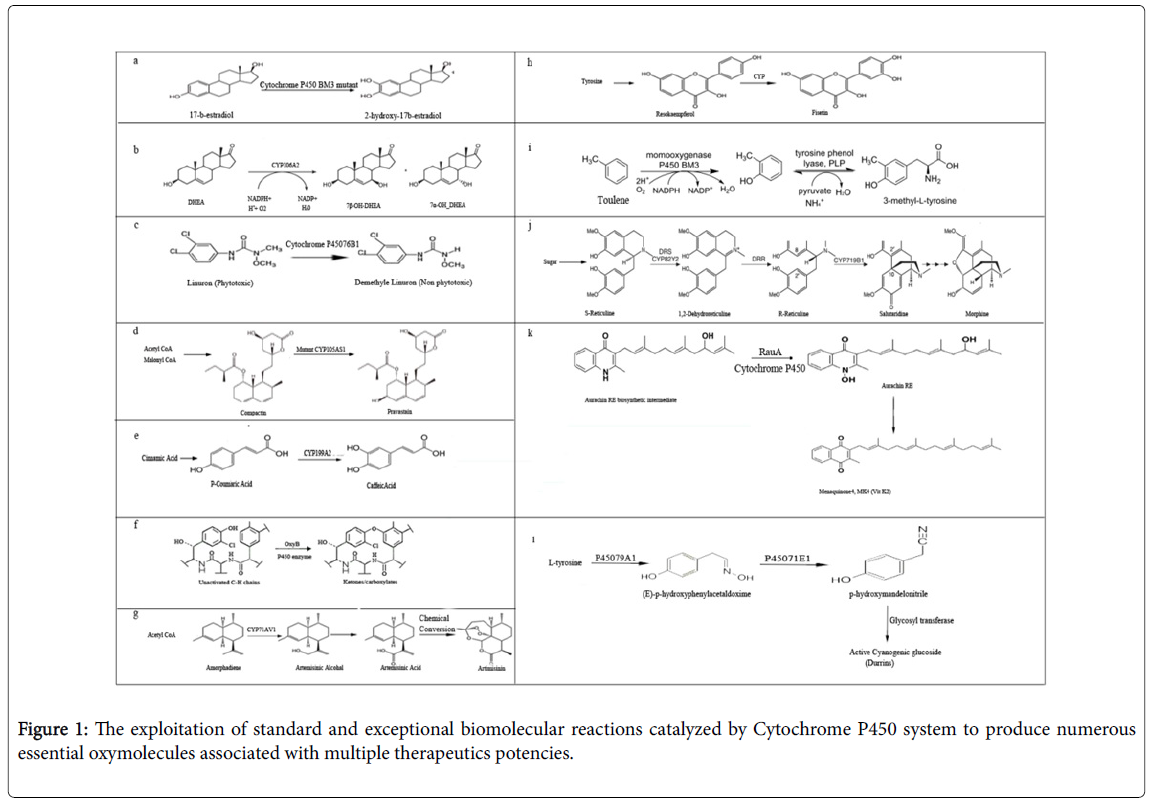
Figure 1: The exploitation of standard and exceptional biomolecular reactions catalyzed by Cytochrome P450 system to produce numerous essential oxymolecules associated with multiple therapeutics potencies.
a. Diverse role of a microbial Cytochrome P450 BM3 has been exploited for the production of unusual oxymolecules because this type of P450 is reported with the maximal activity of monooxygenase out of all P450s [41]. The figure shows the hydroxylation of 2-hydroxy-17-bestradiol (C18H24O3) from 17-b-estradiol by manipulating the catalytic activity of Cytochrome BM3 P450 via direct evolution. 17-b-estradiol (C18H24O3 or 2-OH-Estradiol is an important human metabolite. Human Cytochrome P4501A1 and P4501B1 have also been reported previously for the synthesis of 2-hydroxy-b-estradiol [42]. b. An efficient in vivo whole cell conversion of an engineered Bacillus megaterium for considerable scale production of 7ß- Hydroxydehydroepiandrosterone has shown here. 7beta-OH-DHEA is an essential human metabolite which takes part as immune modulators and neuroprotective agents. The affinity of regio selective hydroxylation of 3-hydroxy-delta5 steroids is attributed to CYP106A2 (B. megaterium MS941 strain) after screening the substrate specificity of steroid library. Increased and volumetric conversion of DHEA into 7ß-Hydroxy-dehydroepiandrosterone and 7a-Hydroxy-dehydroepiandrosterone was achieved by introducing CYP106A2 overexpression in whole cell system [43]. c. The demonstration of increased herbicide degradation and tolerance induced by Cytochrome P45076B1in tobacco (Nicotiana tabacum ) and Arabidopsis . Successful efforts concluded the sole role of Jerusalem artichoke (Helianthus tuberosus Cytochrome in a significant transformation of multiple phenylurea herbicides into nonphytotoxic compounds. Cytochrome P45076B1which catalyzes the N-demethylation of linuron ring which is phytotoxic in nature and hence converted to nonphytotoxic state [44]. Improved herbicide metabolism was observed with the expression of Cytochrome P45076B1 both in tobacco and Arabidopsis carrying a 20-fold increase in tolerance to linuron as well as some other selected phenylureas [45]. d. Penicillium chrysogenum a member of Trichocomaceae family was efficiently engineered for the production of pravastatin: a drug for lowering the cholesterol level. During this pathway, the stereo-selective hydroxylation of usual compactin can be achieved from by introducing an engineered P450 gene (CYP105AS1) which induces the production of active pravastatin instead of epimer epi-pravastatin which is the inactive mode of this essential natural product [46]. e. The elucidation of the caffeic acid pathway in which Cytochrome P450 (CYP199A2) actively catalyze in the conversion of the p-coumaric acid to caffeic acid. The source of the monooxygenase exploited in this pathway is from Rhodo-pseudomonas palustris. Caffeic acid belongs to a natural class of essential metabolite coupled with antidiabetic, anticancer, antioxidants and more of the same pharmacodynamic properties [47]. This engineered pathway is recruited using the expression system of E. coli replacing the role of C3H (4-coumarate 3-hydroxylase) which needed more solvents extract, high temperature and intensive energy to isolate the product yield. Hence it was proved that CYP199A2 shows higher catalytic activity during the conversion of p-coumaric acid into caffeic acid than 4-coumarate 3-hydroxylase. The study reveals the possibility of producing more complex plant molecules with beneficial health effects such as flavonoids and curcuminoids [48]. f. Sequential hydroxylation of a Cytochrome P450 dependant conversion during xiamycin pathway. Cytochrome P450 drives the conversion of unactivated carbon-hydrogen linkages to ketones/carboxylates [49]. A variety of natural products such as alkaloids and biocatalysts have been previously described for Cytochrome P450 catalyzed decarboxylation and oxidation [50]. Alkaloids are associated with multiple drug metabolism strategies for example Quinidine and secologanin act as antiarrhythmic and neurotransmitters respectively [51]. g. The conversion of artemisinin from Acetyl CoA. Artemisinin is an antimalarial drug which can be produced chemically with the help of CYP71AV1 during the metabolic engineering of Saccharomyces cerevisiae pathway. CYP71AV1 facilitate the hydroxylation of Acetyl CoA to amorphadiene and convert further in to artemisinic alcohol then aldehyde and finally to artemisinic acid which is then chemically converted to artemisinin [52]. h. A nine-step biosynthetic cascade process involving the synthesis of fisetin using the expression system of E. coli . Fisetin is an essential plant secondary metabolite carrying anticancer, anti-tumeric, anti-viral and many other important pharmacological effects. The cascade mechanism of the synthesis of fisetin begins with tyrosine initially, which is further proceeded by a nine-step process metabolizing diverse group of intermediates. The final step of this pathway demonstrated the hydroxylation of resokaempferol to fisetin through an unusual Cytochrome P450. The source of this enzyme is reported in Petunia hybrida [53]. i. The illustration of a novel biocatalysis of 3-methyl-L-tyrosine coming from toluene in a cascade manner wherein Cytochrome P450 (BM3) acts as a lyase in combination with another enzyme tyrosine phenol-lyase. PLP: Pyridoxal phosphate. This pathway further elaborates chemoenzymatic paths as well as recent advances and strategies towards the production of pharmaceutically important molecules and intermediates on large scale. The derivatives of 3-methyl-L-tyrosine are important precursor molecules for a variety of effective drugs including saframycin (anticancer drug) [54,55] j. The role of two Cytochrome P450 genes during the biosynthetic pathway in fungi leads to the production of Morphine from sugar [56]. This pathway comprises the conversion of S-reticuline from the feedstock of sugar with the help of reticuline epimerase which is a complex protein consisting various modules i.e., P450 CYP82Y2 and AKR (aldo-keto reductase). S-reticuline catalyzed by DRS (1,2-dehydroreticuline synthase) forming 1,2-dehydroreticuline, while the R-reticuline product is catalyzed by DRR (1,2-dehydroreticuline reductase) coming from AKR domain of epimerase [44]. During this pathway, another P450 CYP719B1facilitate the oxidative C–C bond coupling and the rearrangement of R-reticuline during the conversion of R-reticuline into salutaridine and finally into Morphine [57]. k. A Rhodococcus erythropolis Cytochrome have been shown successfully during aurachin alkaloid pathway. Aurachin alkaloids possess sensitive antibiotic properties when the nitrogen atom on quinolone ring (intermediate of aurachin RE) is hydroxylized by cytochrome P450 RauA [58]. It was also debated in recent past that the homology and topology of aurachin 3-D structure is similar to vitamin K2 (Menaquinone) which is an important unit of respiratory complex in most of the bacterial species, thereby enterprising these natural alkaloids as potential inhibitors for the production of novel drugs by manipulating the mitochondrial respiration [59,60]. l. The involvement of two Cytochrome P450 enzymes during an engineered dhurrin pathway in Sorghum bicolor . In this study, they exploited the expression system of model species of cyanobacteria (PCC6803) for the first time. The first step of this biosynthetic pathway is catalyzed by Cytochrome P45079A1 by the conversion of L-tyrosine into (E)-phydroxyphenylacetaldoxime. Another Cytochrome P45071E1 enzyme likely metabolize their substrate (E-p-hydroxyphenylacetaldoxime) and produce p-hydroxymandelonitrile which is finally glycosylated by UGT85B1 (Glycosyl transferase) to synthesize bioactive durrin Durrins are the important class of plant secondary metabolites (Cyanogenic glucosides). These cyanogenic glycosides are also found in the seed of cherries, quinces, apples and plums. Cyanobacterial strains are believed to be potential candidates for metabolic engineering of P450- dependent pathways [61].
Cognate character of Cytochrome P450s towards floral pigments
It has been shown previously that Cytochrome P450 monooxygenases are extensively involved in the biosynthesis of plant secondary metabolites which directly or otherwise linked with important pharmacological abilities to defend themselves against various kinds of pathogens. Out of all essential plant secondary metabolites three types of floral pigments including flavonoids, carotenoids and betalains are generally associated with the variation in pigmentation of flowers, leaves and fruits in many Plant species [62]. Flavonoids are diverse group of polyphenolic compounds that are present naturally in plants. Flavonoids carry multiple pharmacological roles entailing anti-inflammatory, anti-allergic, vasodilatory, antiviral and antibacterial activities. After subsequent research on flavonoids metabolic profile, their regulation and hypothesizing genetic factors to investigate the floral variations in various plants has lead scientists concerning the role of flavonoids and their subclasses such as flavones, anthocyanin, isoflavones, and flavanol [63].
It is essential and pragmatic to elucidate biochemical scheme and enzymatic network involving regulation of Flavonoids and their affinity towards Cytochrome P450 channels. Besides this, the dissection of all checkpoints, important molecules, intermediates and biocatalysts that are metabolized during this pathway need to explore significantly. Preliminary studies revealed an interesting work on the role of Cytochromes P450s in the biosynthesis of flavonoids and their subclass anthocyanins, which speak about multiple colors phenotypes, for example, orange, red, violet and blue. It was suggested that the presence of variable hydroxyl groups on B-ring of anthocyanidins influence the hydroxylation of this precursor molecule (anthocyanidin) of anthocyanins which is responsible for the fate of anthocyanin (Figure 1). Two Cytochrome P450 monooxygenase subfamilies CYP75B and CYP75A (flavonoid 3'-hydroxylase (F3´H) and flavonoid 3´5´-hydroxylase (F3´5´H) respectively were believed to have an important role in the hydroxylation mechanism of anthocyanidins leading to anthocyanins activation which manifest a fundamental role in floral allotments to the flower color [64].
Flowers with blue color has long been debated by scientists all over the world and some of them has led to the phenomenon of color shift from blue to red and found that this has happened more often than a change from red to blue. A variety of flower color mutations have been studied in many plant species [65]. Most of the plant species observed with a mutation in F3´5´H genes ending up with deficiency of B-ring of delphinidin dependant trihydroxylated-anthocyanin pathway which influence the absence of blue or violet color in flowers. It has been proposed that the genealogical state of many angiosperms beholding active delphinidin dependant trihydroxylated-anthocyanin pattern and hence producing violet or blue flower colors [66]. At the time of their evolutionary study color change from blue to red attributed to a loss of function in F3´5´H genes, keeping in view these facts Cytochrome P450 family has got reasonable attention in the field of plant molecular biology and recombinant DNA technology to manipulate the introduction of blue flowers in angiosperm. Successful switching of these delphinidin dependant trihydroxylated-anthocyanin pattern simply by up-regulating F3´5´H genes resulting in novel blue colors which have been developed commercially. Similarly, the downregulation of F3´5´H genes in normal plants carrying delphinidin dependent trihydroxylated-anthocyanin pathway results in a reduction of the -OH groups on B-ring of anthocyanin which leads to the initiation of pelargonidin dependant monohydroxylated anthocyanins channel producing a shift in the flower color to red or orange.
The regulation and biosynthesis of flavonoids and their different sub-classes including (Isoflavones, anthocyanin, and flavones etc) have also been reported in other Cytochrome P450s in near past. They have successfully indicated that CYP93B (Flavone Synthase FNS) and CYP706C (Flavonoid 3' Monooxygenase) are the enzymes which are essentially responsible to execute the synthesis of various types of flavonoids in coleus forskohlii species [67]. Phylogenetic analysis and homology studies also proved that CYP93B (Flavonoid 3' Monooxygenase) is nearly similar to CYP75A (Flavonoid 3'5' hydroxylase) which is an essential modulator to induce the metabolic regulation during formation of anthocyanins/flavonol from their precursor molecules [68] (Figure 2).
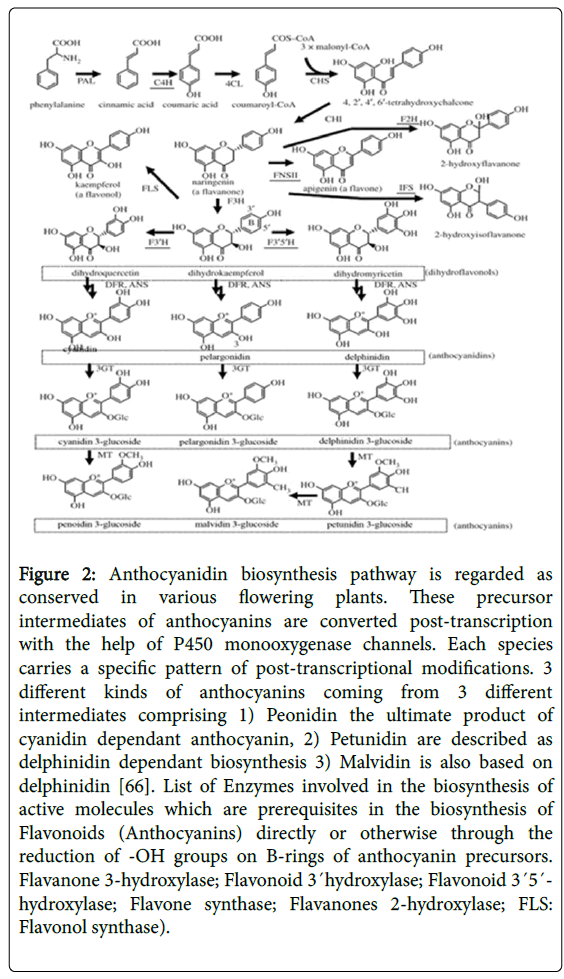
Figure 2: Anthocyanidin biosynthesis pathway is regarded as conserved in various flowering plants. These precursor intermediates of anthocyanins are converted post-transcription with the help of P450 monooxygenase channels. Each species carries a specific pattern of post-transcriptional modifications. 3 different kinds of anthocyanins coming from 3 different intermediates comprising 1) Peonidin the ultimate product of cyanidin dependant anthocyanin, 2) Petunidin are described as delphinidin dependant biosynthesis 3) Malvidin is also based on delphinidin [66]. List of Enzymes involved in the biosynthesis of active molecules which are prerequisites in the biosynthesis of Flavonoids (Anthocyanins) directly or otherwise through the reduction of -OH groups on B-rings of anthocyanin precursors. Flavanone 3-hydroxylase; Flavonoid 3´hydroxylase; Flavonoid 3´5´- hydroxylase; Flavone synthase; Flavanones 2-hydroxylase; FLS: Flavonol synthase).
Plant Cytochrome P450s exhibit remarkable part in various detoxification pathways under the principle of flavonoid biosynthesis respecting post-transcriptional modifications, regulatory motifs, transcriptional factors and defensive compounds. Transgenic lines of Coleus forskohlii which is one of the important plants with multiple pharmacological effects by producing genkwanin (A subclass of flavones) which brings anti-inflammatory effects [69]. Genkwanin inherit these anti-inflammatory consequences in herbs by regulating miR- 101/MKP-1/MAPK pathway through switching off the proinflammatory interceders in macrophages [70]. Cytochrome P450 enzymes catalyze NADPH dependent hydroxylation of a variety of substrates and hence actively participating in complex metabolic cycles such as Flavonoids accumulation. A member of the CYP93B family is found actively involved in the production of Flavonoids [71]. CYP93B gene family in Gerbera hybrida code for flavones synthase II, an enzyme which catalyzes the direct conversion of flavones from flavanones intermediates [72].
Researchers have also indicated that CYP93 families including (CYP93A, CYP93B, CYP93C, CYP93E, and CYP93G) are directly or likely involved in the biosynthesis of flavonoids and isoflavonoids. Metabolic rearrangement in rice has been already debated the assimilation of CYP93 gene family which encodes flavone synthase II (FNS II) enzymes that catalyzes the direct translation of flavanones to flavones [73]. In soybean, CYP93C synthesizes isoflavonoids and CYP93A participates in pterocarpanoid phytoalexin biosynthesis [74]. These branch points are proved to be recommended objectives for reshaping artificial switch in the metabolic flux of definitive metabolites via genetic modification and metabolic engineering. The metabolic engineering of these floral pigments and alteration in their biosynthetic pathways has accomplished a comprehensive understanding of other pathways as well as the structural and functional properties of these enzymes which can influence the novel floral phenotype with multiple health effects in various plant species.
A new area of research on unusual Cytochrome P450s has exploited new edges of various secondary metabolite pathways with the help of 1000 genome sequencing. Functional perspectives of these enzyme associated networks act as hallmarks in plants to produce vaccines, biosensors, derivatives of intricate natural products, drug development and many more of the same [75]. Combining the metabolic engineering and synthetic biology with the discovery of novel Cytochrome P450s via genome mining will introduce us to a wide range of important substances carrying prospective pharmacodynamic properties.
Moreover, the developments of flower in plants with novel floral phenotype coupled with active drug detoxifying compounds are expected to be an exciting step in plant biology. However, it is far essential to understand the efficacy, risks associated with the clinical outcomes and the transmission of information into practical aspects which has not been debated on a wider front so far. To accomplish this landmark future directions should be followed to cognize and understand various checkpoints and components that confer unexpected pharmacodynamic properties for example protein-protein interactions in flavonoid pathways, alteration of delphinidin-based anthocyanins with aromatic acyl groups, variable transgenic expression and non-specific expression, anthocyanin transfer mechanisms, flowering periods, and assembly of flavone C-glucoside. The detailed understanding of these extensive metabolizers in term of pharmacodynamic variability might be apprehended for considerable clinical outcomes in future.
This project was funded by grants the National Natural Science Foundation of China (No. 31771868, 31501366).
The authors declare that they have no conflict of interest.