Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Review Article - (2015) Volume 3, Issue 4
Weeds contribute to severe loss of agricultural crop productivity. Herbicides are efficient, cheap and effective method for their control, but their multiple uses at higher concentration have resulted in induced toxicity and stress in non-target crops. The herbicideinduced toxicity affects growth, photosynthesis, physiological and biochemical characteristics of plants directly impacting yield of plants. It is therefore, necessary to find ways to mitigate the herbicidal effects by increasing the tolerance of crops to their application. Plant growth regulators are known to promote growth and development of plants under optimal and stressful environment. They induce various physiological and biochemical responses and also confer tolerance of plants to abiotic stresses. The present review covers the aspects of herbicidal response of plants and evaluates the contribution of plant growth regulators in mitigating herbicidal stress effects and increasing the tolerance of plants. Further, future research in this direction to enhance our understanding on the relation between plant growth regulators and herbicides to improve tolerance of crop plants is discussed. The study suggests the use of plant growth regulators as a tool in mitigating effects of herbicidal stress together with improved growth and development.
Keywords: Agriculture, Crop loss, Herbicides, Stress, Plant growth regulators, Tolerance
Weeds are undesirable plants that grow along with the cultivated crops that compete with the crop for water, nutrients and light, make harvesting difficult and cause a considerable yield loss [1,2]. The yield losses of field crops (cereals, legumes, Brassica and vegetables) due to weeds are considerable. In Indian context, it has been reported that the loss is 10-60% in Triticum aestivum, 10-100% in Oryza sativa, 30- 40% in Zea mays, 16-65% in Pennisetum glaucum and 50% in Eleusine coracana, 10-45% in Vigna radiata, 10-50% in Cicer arietinum, 30- 35% in Lens culinaris, 10-100% in Glycine max., 10-50% in Pisum sativum and 20-30% in Cajanus cajan, 20-30% in Brassica spp and 20-30% in vegetables [3]. The measures for management of weeds and to overcome the reduction in yield are necessary for sustainable agricultural production. There are different methods available for the control of weeds which include traditional method and advanced technological method, both of which have their own limitation. For example, hand weeding is effective but is cost expensive and difficult owing to labour scarcity and often results in high yield loss due to late weeding. Harrowing is another method for controlling intra-row weeds but is also less effective. Hoeing-ridging method effectively controls both inter and intra row weeds and reduces competitive ability and seed production of uncontrollable weeds [4]. The adoption of intercropping agriculture, a practice to grow two or more crops simultaneously in the same field, reduces the chances of growth of weeds. Another option available is the cover crops which includes suppression of weeds grown at same time of plant growth [5]. These available options require knowledge of soil crop management system, interaction between plants and weeds, and environmental influences associated with the weed control operations [6]. Moreover, there is also growing concern on the involvement of heavy cost in the use of advanced technology methods, such as sensing/guidance technology and robotic method for the control of weeds. Presently, herbicides are commonly used for the control of weeds in the agricultural practices. The herbicides are considered as chemical agents that provide an efficient and cost effective method for the control of weeds. The efficiency of herbicides in the control of weeds has increased their use to protect yield loss of crop plants and sustainability of agriculture, but their prolonged use has prompted the shift in weed population behavior [7] by which they become distinct in morphological, physiological and biochemical nature and may develop resistance to a number of herbicides [8]. Moreover, as the concentration of herbicides is increased it causes inhibition in growth and physiological functions of plants [9]. It is therefore, essential to evaluate the effects of herbicides on non-target plants and impact on soil health, and develop methods that can have positive effects on development of crop plants in the presence of herbicides. Plant growth regulators have been recognized to stimulate growth and development of plants under optimal conditions. They are also involved in tolerance of plants to biotic and abiotic stresses [10-12]. They induce expression of stress related genes for maintaining photosynthetic activity, antioxidant enzymes activation and osmoprotectants accumulation against oxidative stress [13-15]. They can also prove beneficial for reducing the risks of herbicides exposure through modulating plant response to oxidative stress induced by herbicides. However, the importance of plant growth regulators in the control of herbicides needs more attention and collected information is necessary for understanding their role in herbicidal resistance. The present review evaluates the physiological, growth and yield responses of plants to herbicides treatment, and the impact of plant growth regulators in counteracting the adverse effects of herbicides. It also identifies the gap and delineates future prospects of research.
Responses of plants to herbicides
Herbicides control weed plants by disrupting or altering their metabolic processes but also induce stressful conditions and cause damage to the non-target plants. Moreover, they leave residues in soil that are harmful ecologically [16,17]. The plants may or may not recover from the damage caused by herbicide depending on its dose and the severity of the damage. The chances that both crop plants and weeds may develop resistance to the herbicides are also prominent [18]. The interaction between herbicides, weeds, crops and soil explaining the effects of herbicides on weeds, crop plants and soil, and chances of development of resistance in weeds and toxicity in cultivated plants is presented in Figure 1. The nature of herbicides and the dose of application are the important factors determining damage to crop plants as the herbicides may have different capacity to remove weeds without affecting crop. The herbicides with common chemistry are grouped in families and shown in Figure 2. Two or more families may have the same mode of action and thus express same herbicide activity and injury symptoms. Herbicide family and mode of action of some gradient herbicides are presented in Table 1. The commonly growing weeds along with the crop plants and herbicides used are shown in Table 2. The effects of herbicides on various characteristics of plants have been evaluated in the following pages.
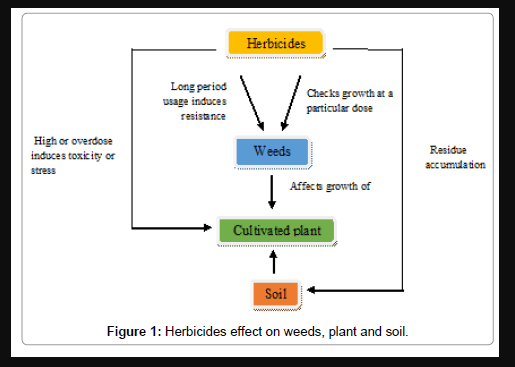
Figure 1: Herbicides effect on weeds, plant and soil.
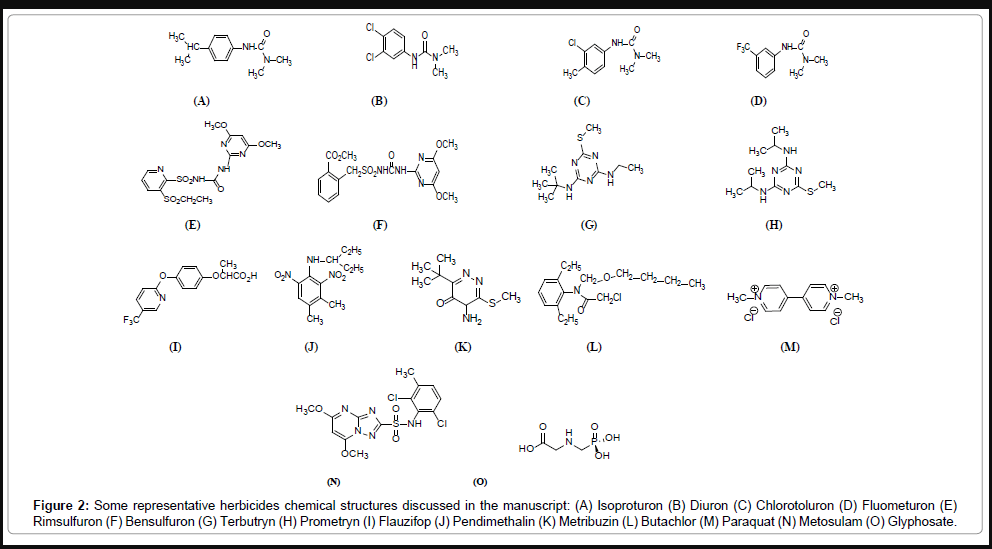
Figure 2: Some representative herbicides chemical structures discussed in the manuscript: (A) Isoproturon (B) Diuron (C) Chlorotoluron (D) Fluometuron (E) Rimsulfuron (F) Bensulfuron (G) Terbutryn (H) Prometryn (I) Flauzifop (J) Pendimethalin (K) Metribuzin (L) Butachlor (M) Paraquat (N) Metosulam (O) Glyphosate.
| S No | Herbicide | IUPAC name | Herbicide Family | Mode or site of action | Reference |
|---|---|---|---|---|---|
| 1. | Quizalofop-p-ethyl FluazifopHaloxyfop | ethyl (2R)-2-[4-(6-chloroquinoxalin-2-yl)oxyphenoxy]propanoate (RS)-2-{4-[5-(trifluoromethyl)-2-pyridyloxy]phenoxy}propionic acid (RS)-2-[4-[(3-chloro-5-trifluoromethyl-2-pyridyloxy)phenoxy]propionic acid |
Aryloxy phenoxy proprionate | Lipid synthesis inhibitors at acetyl CoA carboxylase enzyme (ACCase) | [67] [23] [56] |
| 2. | Imazethapyr | 5-ethyl-2-[(RS)-4-isopropyl-4-methyl-5-oxo-2-imidazolin-2-yl]nicotinic acid | Imidazolinones | Branched chain amino acid synthesis inhibitors at acetolactate synthase (ALS) | [44] |
| 3. | Rimsulfuron Bensulfuron-methyl | 1-(4,6-dimethoxypyrimidin-2-yl)-3-(3-ethylsulfonyl-2-pyridylsulfonyl) urea methyl 2-[(4,6-dimethoxypyrimidin-2-yl)carbamoylsulfamoylmethyl]benzoate |
Sulfonylurea | Branched chain amino acid synthesis inhibitors at acetolactate synthase (ALS) | [28] [66] |
| 4. | Terbutryn | N2-tert-butyl-N4-ethyl-6-methylthio-1,3,5-triazine-2,4-diamine | Triazine | Photosynthesis inhibitors at PS II | [31] |
| 5. | Prometryn | N2,N4-diisopropyl-6-methylthio-1,3,5-triazine-2,4-diamine | [26] | ||
| Metribuzin | 4-amino-6-tert-butyl-3-methylsulfanyl-1,2,4-triazin-5-one | Triazinones | Photosynthesis inhibitor at PS II | [41] | |
| 6. | Diuron Isoproturon |
3-(3,4-Dichlorophenyl)-1,1dimethylurea 3-(4-isopropylphenyl)-1,1-dimethylurea |
Phenylurea | Photosynthesis inhibitor at PS II | [72] [25] |
| Chlorotoluron Fluometuron |
3-(3-chloro-p-tolyl)-1,1-dimethylurea 1,1-Dimethyl-3-[3-(trifluoromethyl)phenyl]urea |
[38] [28] |
|||
| 7. | Paraquat | 1,1'-Dimethyl-4,4'-bipyridinium dichloride | Bipyriduliums | Photosynthesis inhibition at PS I | [55] |
| 8. | Glyphosate | N-(phosphonomethyl)glycine | Glycine | Aromatic amino acid inhibitor at EPSP synthase | [62] |
| 9. | Pendimethalin | 3,4-Dimethyl-2,6-dinitro-N-pentan-3-yl-aniline | Dinitroanilines | Mitotic disruptors or microtubule assembly inhibitors | [19] |
| 10. | Butachlor Acetachlor |
N-(Butoxymethyl)-2-chloro-N-(2,6-diethylphenyl)acetamide 2-Chloro-N-(ethoxymethyl)-N-(2-ethyl-6-methylphenyl)acetamide |
Acetanilide | Long chain fatty acid inhibitor | [41] [66] |
| 11. | Metosulam | 2',6'-dichloro-5,7-dimethoxy-3'-methyl[1,2,4]triazolo[1,5-a]pyrimidine-2-sulfonanilide | Triazolopyrimidine (TP) sulfonanilide | Acetolactate synthase ALS or (acetohydroxyacid synthase AHAS) inhibitor | [21] |
Table 1: Family, mode of action and site of action of gradient herbicides used for crops discussed in the review.
| S No | Crop | Weeds | Herbicides | References |
|---|---|---|---|---|
| 1. | Wheat (Triticum aestivum) | Phalaris minor Avena fatua Argemone Mexicana Rumex dentatus |
Clodinafop Fenoxaprop Sulfosulfuron Isoproturon |
[2] |
| 2. | Maize (Zea mays) | Amaranthus spinosus Cynodon dactylon Commelina benghalensis Bidens pilosa |
Metribuzin Butachlor Chlorimuron ethyl |
[86] [41] |
| 3. | Rice (Oryza sativa) | Fimbristylis miliacea Cyperus iria Cyperus difformis |
Acetachlor Bensulfuron-methyl |
[87] [66] |
| 4. | Black gram (Vigna mungo) | Parthenium Tridax Hyptis Cortaderia selloana |
Quizalofop-p-ethyl | [88] [67] |
| 5. | Mung bean (Vigna radiata) | Cynodon dactylon Echinochloa colona Chenopodium album Amaranthus viridis |
Pendimethalin Oxyfluorfen Quizalofop-p-ethyl |
[89] |
| 6. | Soyabean (Glycine max) | Trianthema portulacastrum Celosia Echinochloa colona |
Flauzifop Pendimethalin Trifluralin |
[90] |
| 7. | Potato (Solanum tuberosum ) | Amaranthus retroflexus Sinapsis arvensis |
Metribuzin | [91] |
| 8. | Onion (Allium cepa) | Anthemis palastina Amaranthus spp. Sinapsis arvensis Trigonella arabica |
Oxyfluorfen Trifluralin Pendimethalin Metribuzin |
[92] |
| 9. | Pea (Pisum sativum) | Chenopodium album Echinocloa crus-galli Sonchus arvensis Fallopia convolvulus |
Imazethapyr Metribuzin Pendimethalin |
[93] [94] |
| 10. | Cucumber (Cucumis sativus) | Capsella bursa-pastoris Stellaria media Chenopodium album Brassica kaber |
Triflularin Sethoxydim Clomazone |
[95] |
Table 2: Some common weeds present in crop plants and herbicides used to control the weeds.
Photosynthesis
Herbicidesnegatively impact photosynthetic characteristics of plants. Their use has shown to decrease photosynthetic pigments, carbon assimilation and efficiency of pigment system II (PSII). Application of pendimethalin on Foeniculum vulgare leaves [19]; chevalier in Triticum aestivum cultivars [20] and metosulam in Vicia faba plants [21] has shown to decrease photosynthetic pigments. In a study on Zea mays, application of glyphosate at 10 mM resulted in decreased photosynthesis-related proteins [22]. The inhibition in carbon assimilation with 0.20 g L-1 flauzifop and 0.14 g L-1 haloxyfop in Cucumis sativus has been found [23]. Studies have shown that chlorophyll synthesis and content were decreased in plants with herbicides application. The decreased content of chlorophyll was found in Triticum aestivum with chlorimuron-ethyl at 300 μgkg-1, isoproturon at 2 mg kg-1, prometryne at 4 mg kg-1, sekator at 150 mL ha-1 and zoom at 120 gha-1 [24-27]; in Vicia faba with fluometuron at 2.98 kg ha-1 [28], in Glycine max. cultivars with diuron at 1, 2 and 5 ppm [29]. The decrease in photosynthetic characteristics with herbicides might have involved its effects on chlorophyll biosynthetic pathway and enzymes of carbon assimilation. The details on the key points in the biosynthetic pathway of photosynthetic pigments and activity of carbon assimilation enzymes are still lacking and needs attention. Chlorophyll fluorescence is a simple and widely used method to study the mechanism of photosynthesis and is used as marker to detect stress in plants. The herbicides application may block photosynthesis related intermediate metabolites and affect fluorescence emission. The herbicides that inhibit glutamine synthase, protoporphyrinogen oxidase and carotenoid biosynthesis affect chlorophyll fluorescence [30], and even lipid synthesis inhibitor herbicides induce chlorophyll fluorescence. Application of 1.96 kg ha-1 Terbutryn to Vicia faba plants strongly decreased Fv/Fm ratio and CO2 assimilation capacity [31]. Recently, Ref. [32] have shown that application of imidazolinone group of herbicides to Oryza sativa changed the photosynthetic metabolism of plants and chlorophyll a fluorescence.
Osmolytes
The production of osmolytes is an adaptation against stress in plants [33]. These osmolytes include polyols (sugars, fructans, mannitol, pinitol and inositol), amino acids and their derivatives (proline, isoleucine, valine, betaine, glycinebetaine). Proline is widely studied osmolyte that acts as a cellular protector in several plant species by scavenging ROS produced in response to abiotic stress [34-36]. It provides resistance against stress either by uniting oxygen and free radicals generated in stress [37] or by efficiently scavenging OH- and O2 -and inhibit lipid peroxidation, and acts as a source of carbon and nitrogen during stress. Proline may be accumulated in plants in large amounts during stress either due to enhanced synthesis or reduced degradation. Herbicide treatment results in accumulation of proline as a measure of resistance to stress. Application of 5-25 mg kg-1 chlorotoluron to Triticum aestivum [38] and 10-5, 10-6, 10-7% metosulam to Vicia faba plants [21] resulted in accumulation of proline. Soluble carbohydrates increased in Glycine max. by increasing diuron application from 1<2<5 ppm [29]. The increase in soluble sugars content in plants is an adaptive response to support basal metabolism under stress. Soluble amino acids have been found increased more in leaves of susceptible cultivars of Glycine max. plants than resistant cultivars in response to glyphosate application at 2:100 ratio with water [39], but the content of soluble amino acids increased in roots in all the cultivars. The content of myoinositol increased in roots of Brassica napus seedlings after glyphosate application in a concentration dependent manner from 40 -2000 μM [40]. In contrast, application of chlorimuron-ethyl at 20 g ha-1 decreased the amino acids; valine, leucine and isoleucine in Triticum aestivum and Zea mays plants [41]. These studies are however, limited to the quantification of osmolytes, and the key points influenced by the herbicides are not known. Also, studies are limited to few osmolytes and leaves scope for determination of array of osmolytes that plants require for stress tolerance.
Nitrogen and sulfur assimilation
Nitrogen is primary nutrient element and forms backbone for proteins and nitrogenous compounds required for growth and development of plants. The energy needed for N assimilation comes from the photosynthetic process. There exists a strong correlation between N uptake and chlorophyll fluorescence parameters as shown in Triticum aestivum [42]. Herbicides may influence N assimilation directly or through its effects on photosynthesis. The activity of nitrate reductase (NR) and nitrite reductase (NiR) was found inhibited on application of isoproturon at 1 kg ha-1, accompanied with the inhibition of electron transfer [43]. The reduction in carbon dioxide fixation also reduced the activity of NR, as energy and carbon skeleton required for N assimilation are provided by photosynthesis. Ref. [44] reported that the decrease in nitrate uptake due to 52 μM imazethapyr was partly related to a reduced stomatal conductance in Glycine max. The decrease in NR activity with 3 L ha-1 butachlor and 20 g ha-1 chlorimuron-ethyl was found in Triticum aestivum and Zea mays [41]. In contrast, recently Ref. [45] found increased N uptake with 1.50 kg ha-1 isoproturon application to Triticum aestivum. Sulfur makes a part of vitamins, co-enzymes, phytohormones and reduced sulfur compounds involved in the regulation of metabolism of plants under optimal and stressed conditions [46]. Reduced glutathione (GSH) is formed from its assimilation and reduction [47] and may have role in herbicide detoxification [48] since GSH has well accepted role in protection of plants from oxidative damage by keeping ROS under control [49]. In spite of the recognized role of S and S-compounds in abiotic stress tolerance, only few studies have been conducted to show the influence of herbicides on S assimilation. Application of 10 mM glyphosate increased glutathione-S-transferase (GST) activity in Pisum sativum [50] and Zea mays [22]. Ref. [25] and Jiang and Yang [26] also found increase in GST activity in Triticum aestivum on treatment with 5 and 10 mg kg-1 isoproturon and low concentration of prometryne at 4, 8 and 12 mg kg-1. The detailed studies on S assimilation and S-compounds could provide information on better tolerance of plants to herbicide treatment.
Antioxidant system
The enhanced production of reactive oxygen species (ROS; superoxide radicals, O2-; hydroxyl radicals, OH-; perhydroxyl radicals, HO2-; alkoxy radicals, RO-; hydrogen peroxide, H2O2; singlet oxygen, 1O2) takes place under abiotic stress conditions which damage plants primary metabolites; protein, lipid, carbohydrate, DNA and halt metabolic activities of cell [51]. Plants require activation of antioxidative system, especially ROS-scavenging systems within the cells that help in stress tolerance and contribute to continued plant growth under stress conditions [52]. The effects of herbicides on ROS and antioxidant system have been shown in plants. Application of isoproturon increased malondialdehyde production, while decreased the content of GSH and ascorbic acid and reduced the activity of SOD, CAT and APX in Triticum aestivum seedlings [53]. In another study, application of isoproturon resulted in increased CAT activity and decreased peroxidase (POD) activity [54]. Lipid peroxidation increased in Triticum aestivum cultivars by application of chevalier at 0.9, 1.2, 1.5 mg pot-1 [20]. The activity of antioxidant enzymes; SOD and APX increased by application of 50 μM haloxyfop-ethoxyethyl and 15 μM paraquat to Triticum aestivum [55,56]. A concentration dependent change in antioxidant activity has been reported with prometryne in Triticum aestivum; low concentration of prometryne at 4, 8, 12 mg kg-1 increased the activity of SOD, POD, CAT, APX activity in leaves, while higher concentration 16, 20, 24 mg kg-1 decreased the activity [26]. The induced accumulation of superoxide anion (O2 -) and H2O2 in leaves and peroxidation of plasma membrane lipids were found with 5-25 mg kg-1 chlorotoluron to Triticum aestivum in a concentration dependent manner. Moreover, the total activity of POD in roots was enhanced by 5 mg kg-1, but CAT activity generally suppressed under the chlorotoluron exposure at 20-25 mg kg-1 [38]. In contrast, glyphosate application at 10 μM increased CAT activity in Zea mays [22]. The treatment of Zea mays with herbicides 2.98 kg ha-1 fluometuron and 0.015 kg ha-1 rimsulfuron increased POD activity [28]. However, the application of increasing concentration of metosulam herbicide from 10-7 to 10-5% in plants resulted in decrease of POD activity and increase of CAT and APX in Vicia faba plants [21]. The activity of SOD and CAT declined in proportion to concentration of paraquat (0.1, 1, 2 μM) applied to Lactuca sativa, Phaseolus coccineus and Pisum sativum [57].
Growth and yield
The use of herbicides negatively influences growth and yield of crop plants. The application of 8.5 mL L-1 pendimethalin reduced growth characteristics; shoot length, fresh and dry weight of Foeniculum vulgare plants [19]. Application of phenoxy herbicide, MCPA (4 -chloro-2- methyl-phenoxy acid) at 1.5 L ha-1 has been reported to reduce shoot length in Triticum aestivum [58], while the use of metosulam at 10-7 to 10-5% induced cytotoxic effects in Vicia faba and decreased fresh and dry weight of plants [21]. Glyphosate treatment at 2 to 10 mM concentration in two varieties of Vigna radiata produced differential effects. The germination percentage was similar to control at 6 mM, but was increased at 8 mM in one variety. On the other hand, fresh weight declined by two-fold on increasing concentration to 10 mM in both the varieties [59]. In another study, glyphosate application at 0.62 and 0.84 kg ha-1 to Triticum aestivum [60], 100, 250 and 500 μg L-1 to Allium cepa [61] and 1, 2, 4, 6, 8, and 10 mM to six Vigna mungo genotypes [62] was found to inhibit germination in a concentration dependent manner, and decreased leaf area and shoot dry biomass of Glycine max. at 800, 1200, and 2400 g ha-1 [63]. Higher concentration of glyphosate at 500 μg L-1 was found inhibitory on root growth in Allium cepa [61] and at 10 mM for growth of six Vigna mungo genotypes [62]. Haloxyfopethoxyethyl at 50 μM inhibited growth of Triticum leaves [56], while increasing dose of chevalier at 0.9, 1.2 and 1.5 mg pot-1 resulted in a greater decrease of roots than shoots of Triticum aestivum [20]. Root length and shoot length [25], shoot fresh and dry weights of Triticum aestivum were decreased after treatment with isoproturon [53] at 2-20 mg kg-1 and 2.5 L ha-1, respectively. Application of 1 kg ha-1 metribuzin, 3 L ha-1 butachlor and 20 g ha-1 chlorimuron-ethyl to Triticum aestivum and Zea mays seedlings resulted in decrease of shoot fresh weight [41]. Pendimethalin at 0.5-10 ppm reduced the germination percentage of Zea mays [64], while isoproturon at 2.5 kg ha-1 significantly reduced the fresh and dry weight of shoot and root of Zea mays seedlings [65]. Fresh weight of Oryza sativa shoot and root was reduced by the application of acetachlor at 3.2 μmol L-1 and bensulfuron-methyl at 0.96 μmol L-1 [66]. Higher concentration of quizalafop-p-ethyl at 1, 1.5 and 2% decreased the yield of Vigna mungo [67]. Reduction in number of tillers per plant and grain yield in Triticum aestivum was recorded with 1 kg ha-1 isoproturon [43], while bromoxynil decreased the number of spikes, main spike length, grains weight and straw per plant [68]. Pendimethalin decreased yield components of Foeniculum vulgare [19]. Table 3 presents the summary of studies on the effects of different doses of herbicides on various characteristics of plants.
| S No | Herbicides | Plants name | Concentration of herbicides | Response | Affected parameters | References |
|---|---|---|---|---|---|---|
| 1. | Isoproturon | Triticum aestivum | 2, 3.5, 5, 10, 20 mg kg-1 | - | Growth, photosynthesis, chlorophyll content | [25] |
| 2. | Isoproturon | Triticum aestivum | 2, 3.5, 5, 10 and 20 mg kg-1 | + | SOD, APX, POD and GST | [25] |
| 3. | Pendimethalin | Zea mays | 0.5, 1.0, 2.5, 5.0 and 10.0 ppm | - | Seed germination | [64] |
| 4. | Chlorotoluron | Triticum aestivum | 5, 10, 15, 20 and 25 mg kg-1 | - | Biomass | [38] |
| 5. | Chlorotoluron | Triticum aestivum | 5, 10, 15, 20 and 25 mg kg-1 | + | Enhanced total activity of POD and APX, and proline content | [38] |
| 6. | Chlorotoluron | Triticum aestivum | 5, 10, 15, 20 and 25 mg kg-1 | - | CAT activity | [38] |
| 7. | Metribuzin | Triticum aestivum, Zea mays | 1 kg ha-1 | - | Shoot fresh and dry mass, GS and GOGAT activity | [41] |
| 8. | Butachlor | Triticum aestivum, Zea mays | 3 L ha-1 | - | Shoot fresh and dry mass, GS and GOGAT activity | [41] |
| 9. | Chlorimuron-ethyl | Triticum aestivum, Zea mays | 20 g ha-1 | - | Shoot fresh and dry mass, GS and GOGAT activity | [41] |
| 10. | Acetachlor | Oryza sativa | 3.2 µmol L-1 | - | Root and shoot biomass, NR activity | [66] |
| 11. | Bensulfuron-methyl | Oryza sativa | 0.96 µmol L-1 | - | Root and shoot biomass, NR activity | [66] |
| 12. | Glyphosate | Vigna radiata | 2, 4, 6, 8, and 10mM | - | Germination percentage,Root length, Fresh weight | [59] |
| 13. | Glyphosate | Vigna radiata | 2, 4, 6, 8, and 10mM | + | CAT,POD, GST | [59] |
| 14. | Paraquat | Lactuca sativa, Phaseolus coccineus, Pisum sativum |
0.1, 1, and 2 μM | - | Chlorophyll and carotenoid content, SOD, CAT | [57] |
| 15. | Alachlor | Lactuca sativa, Phaseolus coccineus, Pisumsativum |
0.2,1, 2, 20, 100, and 200 μM |
- | Chlorophyll and carotenoid content, SOD, CAT | [57] |
| 16. | Metolachlor | Lactuca sativa, Phaseolus coccineus, Pisum sativum |
0.2,1, 2, 20, 100, and 200 μM |
- | Chlorophyll and carotenoid content, SOD, CAT | [57] |
| 17. | Paraquat | Triticum aestivum | 15, 30 and 60 µM | - | Chlorophyll fluorescence, chlorophyll and carotenoid content | [55] |
| 18. | Paraquat | Triticum aestivum | 15, 30 µM | + | SOD and POD | [55] |
| 19. | TOPIK (active ingredient-clodinafop-propargyl) | Triticum aestivum. Secale cereale, Zea mays |
8, 80 and 800 µg L-1 | + | LPO Intensity; CAT and APX | [9] |
| 20. | Metosulam | Vicia faba | 10-5, 10-6 and 10-7 % | - | Photosynthetic pigment, Fresh and dry weight, Mitotic index; Cell division | [21] |
| 21. | Metosulam | Vicia faba | 10-5, 10-6 and 10-7 % | + | Chromosomal abberations, Proline content | [21] |
| 22. | Prometryn | Triticum aestivum | 4, 8, 12, 16, 20 and 24 mg kg-1 | - | Growth and biomass, Chlorophyll content | [26] |
Table 3: Some representatives herbicides used in agricultural practices and their effect on various parameters studied. Abbreviations: -: Decrease; +: Increase; APX: Ascorbate peroxidase; CAT: Catalase; GOGAT: Glutamine oxoglutarate aminotransferase; GR: Glutathione reductase; GST: Glutathione-S-transferase; LPO: Lipid peroxidation; MDA: Malondialdehyde; NR: Nitrate reductase; POX: Peroxidase; SOD: Superoxide dismutase.
Role of plant growth regulators in herbicidal stress tolerance
Among the several strategies adopted by plants to counteract the adverse effects of abiotic stress, plant growth regulators provide signals to allow plants to survive under stress conditions. The modulation of their levels in plants in environmental stressed conditions modifies the signaling pathways and induces resistance in plants [69,70]. They are one of the key systems integrating metabolic and developmental events in the whole plant, and are essential for many processes throughout the life of a plant, influencing the yield and quality of crops. The compounds that have been considered as plant hormones are described as classical hormones (auxin, cytokinin, gibberellins, ethylene, abscisic acid) and newly added compounds (brassinosteroids, salicylic acid, nitric oxide, jasmonic acid).
Classical hormones
Auxins have been reported to impart resistance to herbicide toxicity. Application of 2, 4-dichlorophenoxyacetic acid at 60-120 g ha-1 was found suitable for protecting cultivated plants against the phytotoxic action of 20-60 g ha-1 clodinafop-propargyl [71]. The protective effect of benzyladenine at 1, 10, 100 μM reduced the decrease in chlorophyll, carotenoid and ascorbic acid contents of Zea mays leaves caused by paraquat. Pretreatment with benzyladenine significantly increased SOD activity after paraquat treatment [72]. Phenylurea cytokinin at 0.1 mM reduced the deleterious effects of 10 mM glyphosate in Zea mays plants. It induced its protective action by hardiness in the antioxidant defense systems [22]. Treatment of Zea mays leaves with exogenous abscisic acid led to significant decrease in the content of malondialdehyde, the percentage of ion leakage and the level of protein oxidation under 50 μM paraquat stress. Abscisic acid induced protection against oxidative damage generated by paraquat through mitogen activated protein kinase cascade in Zea mays leaves [73]. Pretreatment with 5 μM abscisic acid enhanced tolerance of Oryza sativa plants to 10 μM phosphinothricin and reduced ammonium ion accumulation in leaves [74]. The increase in endogenous abscisic acid content in leaves was closely related to phosphinothricin tolerance.
Newly added plant hormones
Besides these classical hormones, salicylic acid is an important signaling molecule that plays a vital role in protecting plants against herbicide toxicity [75]. It regulates plant response to organic contaminants and provides a basis to control herbicide contamination in crop production. Salicylic acid at 500 μmol L-1 mediates 10 μmol L-1 paraquat tolerance in Hordeum vulgare[76] and in Zea mays seedling by enhancing chlorophyll and carotenoid content and leaf total protein. In addition, salicylic acid at 0.5 and 1.0 mM significantly reduced free proline, lipid peroxidation and CAT activity in 10 mM paraquat stressed Zea mays seedlings [77]. It improved growth of napropamide treated Brassica napus and reduced napropamide levels in plants. Its application at 0.1 mM also decreased the abundance of O2- and H2O2 as well as activity of SOD, CAT, APX and increased activities of POD, guaiacol peroxidase (GPX) and GST in 8 mg L-1 napropamide-exposed plants [78]. Nitric oxide has been shown to reduce herbicide toxicity by its protective effects on plants. The reduction in protein content in Oryza sativa leaves by paraquat was prevented by nitric oxide by reducing the level of malondialdehyde and improving the activity of antioxidant enzymes, such as SOD, POD and GR [79]. Nitric oxide given as sodium nitroprusside at 50, 100 and 200 μM protected Glycine max plants by scavenging the ROS generated by 168 g ha-1 lactofen [80]. The protective role of brassinosteroids against biotic and abiotic stress has been documented in literature, but the action of brassinosteroids against damage by herbicides is not well validated. There are few studies that demonstrate that brassinosteroids function to protect metabolism against effects of herbicides. Study of Pinol and Simon [31] has shown protective action of brassinosteroids against photosynthesis-inhibiting herbicides. The study has shown that epibrassinolide at 2 x 10-6 or 2 x 10-5 mM induced the recovery from damage caused by 1.96 kg ha-1 terbutryn in Vicia faba plants, as detected by chlorophyll fluorescence and photosynthetic CO2 assimilation. Polyamines (putrescine, spermidine and spermine) are also known to participate in plants for protection against abiotic stress conditions [81,82]. Spermidine offered protection against paraquat toxicity in Raphanus sativus seedlings [83]. Diethylenetriamine polyamine increased the protection against 2, 4-D herbicide in the triticale plants [84]. Application of polyamines significantly prevented the loss of chlorophyll content, reduced the lipid peroxidation caused by diuron herbicide in leaves of Zea mays and also increased the activity of SOD, GPX and GR [85]. The use of plant growth regulators results in the development of resistance against herbicides through activation of defense mechanisms, such as control of ROS production, enhanced osmolytes biosynthesis, nutrients homeostasis and activation of antioxidant system resulting in protection of cells from damage. These effects together ensured higher photosynthesis, growth and yield. The possible mechanisms involved in the herbicidal stress tolerance through the use of plant growth regulators are shown in Figure 3. However, there is need to study the extent of mechanisms induced by individual plant growth regulators in mitigation of herbicidal stress.
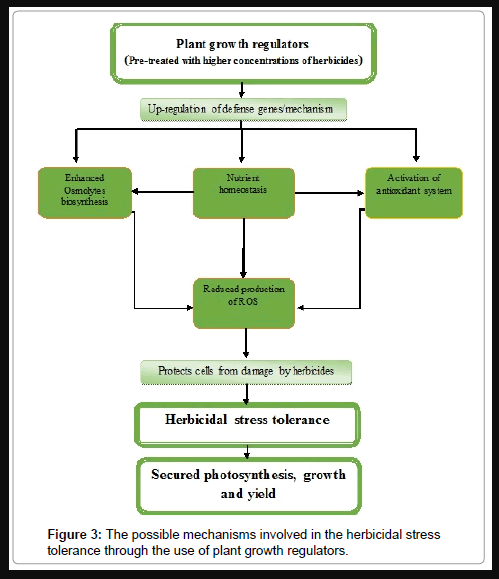
Figure 3: The possible mechanisms involved in the herbicidal stress tolerance through the use of plant growth regulators.
The use of herbicides has become a potential tool in modern agriculture to get rid of weeds menace, but unknowingly the indiscriminate application of herbicides has resulted in adverse effects on the morphological, physiological and biochemical characteristics of crop plants. The production of biomass of crop plants is negatively affected due to impaired photosynthetic activity generated by toxicity induced by herbicides. The traditional as well as modern methods for the control of weeds are not cost effective. Phytohormones have potential in protecting and/or alleviating adverse herbicidal effects in plants as they have significant role in antioxidant metabolism and synthesis of metabolites known for providing stress tolerance. However, the studies related to plant growth regulators action in protection against herbicides are few and in depth experimental studies are required to explain the mechanisms responsible for counteracting herbicidal effects. The developmental stage of plants is important in the use of herbicides to reduce their effects and the dose of applied herbicide needs to be evaluated for crops to reduce their effects. The target points in biosynthetic pathways are to be investigated on which herbicides act, so that these key points can be manipulated for reversing the effects of herbicides. Moreover, the use of molecular tools may help in explaining the proteins involved in providing tolerance to herbicidal effects. In addition, the gap in knowledge and understanding of the effects of herbicides action on various physiological, biochemical and molecular levels are required to explore the regulatory points for combating the effects. The modulation in the endogenous levels of plant growth regulators or signaling for gene expression in plants is required to cope up with the stressful conditions. Further, more studies should be focused on unraveling the cross-talk between different plant hormones and other molecules in regulating the toxicity induced by herbicides.
Authors (SV and FR) gratefully acknowledge the research fellowship by the University Grants Commission, New Delhi, India.