Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2012) Volume 3, Issue 1
The catch rate, diet, size, and reproduction of 565 specimen of Labeo coubie were studied over 24 months (January 2005–January 2007) in three reaches [upstream, midstream (waterfalls region) and downstream] along the 6817.7 m length of Agbokim waterfalls during wet and dry seasons. Data were processed using fecundity (F), allometric coefficient (b), Fultons condition factor, and diet breadth. Fecundity values were 10411.0 ± 121, 15102.0 ± 303.5 and 24143.3 ± 662 eggs for fish from upstream, midstream, and downstream, respectively, with overall mean value of 19811.3 ± 223 eggs. The relationships of fecundity with total weight, ovary weight, ovary length, body breadth were positively linear. Mean allometric coefficient (b) of the length–weight relationship of fish in the three reaches were 2.662 ± 0.291 (upstream), 2.592 ± 0.368 (midstream), and 2.622 ± 0.315 (downstream) and all showed significant departure from cubic value (negative allometry). Detritus, phytoplanktons and plant materials were the dominant food items in the diet, contributing 36.8%, 32.1%, and 17.2% of stomach content, respectively. Food Richness of 7 and Gut Repletion Index, 100% were recorded. The low mean diet breadth exhibited by fish from the three sampling sites showed that the species has low trophic flexibility and cannot easily change feeding habit. Mean condition factor varied from 0.442 ± 0.127 upstream to 0.648 ± 0.146 downstream. Downstream therefore provided better spawning conditions for Labeo coubie in the tropical waterfalls ecosystems.
Keywords: Diet, length?weight, waterfalls, Labeo coubie, reproductive biology
The African carp Labeo coubie (L. coubie) occurs in most of the major rivers in Africa including Nigeria, Senegal, Gambia, Ivory Coast, Liberia, Zaire, and Gabon. L. coubie has protractile mouth equipped with well-developed lips forming suckers with jaws devoid of teeth. Abdominally situated pelvic fin has forked tail with rounded lobes. The fish is silvery in color [1] and is commonly cultured in ponds. It is known for its sweet tastes with rich source of protein for the common people of Africa [2]. Of the four species of African carp, L. senegalensis, L. pseudocoubie, L. rhohita, and L. coubie; L. coubie is the most common species [3]. There is therefore need to study some aspects of the biology of the fish species for sustainable management and conservation of the wetland species. Length–weight relationship studies have been done in different water bodies and on different fishes in African regions. Notably among these are some studies on Nigerian fresh water fishes [4–8] and others include those for Chrysichthys spp. [9] from the southern part of the river Nile, Egypt; for mullet in New Calabar River [10], for Nigerian coastal waters [11], and for Mugilcephalus in Bonny Estuary, Nigeria [12]. The dimensional variations exhibited by the fishes are governed by the various ecological parameters in various locations of the freshwater. Knowledge about fecundity of a fish is essential for evaluating the commercial potentialities of its stock, life history, practical culture, and actual management of the fishery [9,10] Many works had been done on the fecundity of different fishes [13–19]. Dietary analysis of organisms in their natural habitats also enhances the understanding of the growth, productivity, abundance, and distribution of such organisms, as well as knowledge of their trophic relationships [20–23]. Very little work has been done on the diet, size, and reproductive biology of L. coubie in waterfalls ecosystem. This paper therefore presents some aspects of the biology of the species. Such data will provide useful information for the culture of L. coubie in different environmental conditions.
Study area
The study area is Agbokim Waterfalls in Cross River State, Nigeria (Figure 1). Agbokim Waterfalls is located in latitude 5º59’ North and longitude 8º45’ East. It is bounded in the West by the Cross River and in the North by the Cameroon high forests. The climate of Agbokim Waterfalls is the tropical hinter-land type, with wet (May–November) and dry (December–April) seasons. Mean annual temperature ranged between 20°C and 32°C and annual total average rainfall, from 1450 to 3015 mm. The vegetation is the rainforest type with soil consisting of deep laterite and dark fertile, clayey, and loamy soils. Agbokim Waterfalls is drained by two small rivers, Ekue and Bakue, which are tributaries of the Cross River system. Of ecological importance are numerous small pools and swamps which are found along the length of the waterfalls. The high annual discharge and rainfall of the Agbokim Waterfalls provide excellent buffers against natural ecological stresses such as drought [18], which can lead to the loss of important fish taxa. The total length of the waterfalls is 6817.73 m. For the purpose of this study, the waterfalls is divided into upstream, midstream (region of water fall), and downstream reaches. Upstream is close to cocoa farms with high forest cover along its shoreline and moderately fast water currents during the dry season and very fast water current during the wet season. Midstream has heavy water turbulence during the wet season and moderate water turbulence during the dry season with shoreline sparsely shaded with vegetation while downstream water current is low with an extensive wide area. The length upstream is 2003.13 m, midstream 807.42 m and downstream 4007.18 m. Mean width upstream, midstream, and downstream are 26.34 ± 2.2, 32.84, and 40.52 ± 10.9 m, respectively. Upstream mean height is 31.87 ± 3.4 m while the depths upstream and downstream are dependent on variation of the volume of water from rainfall and water discharge.
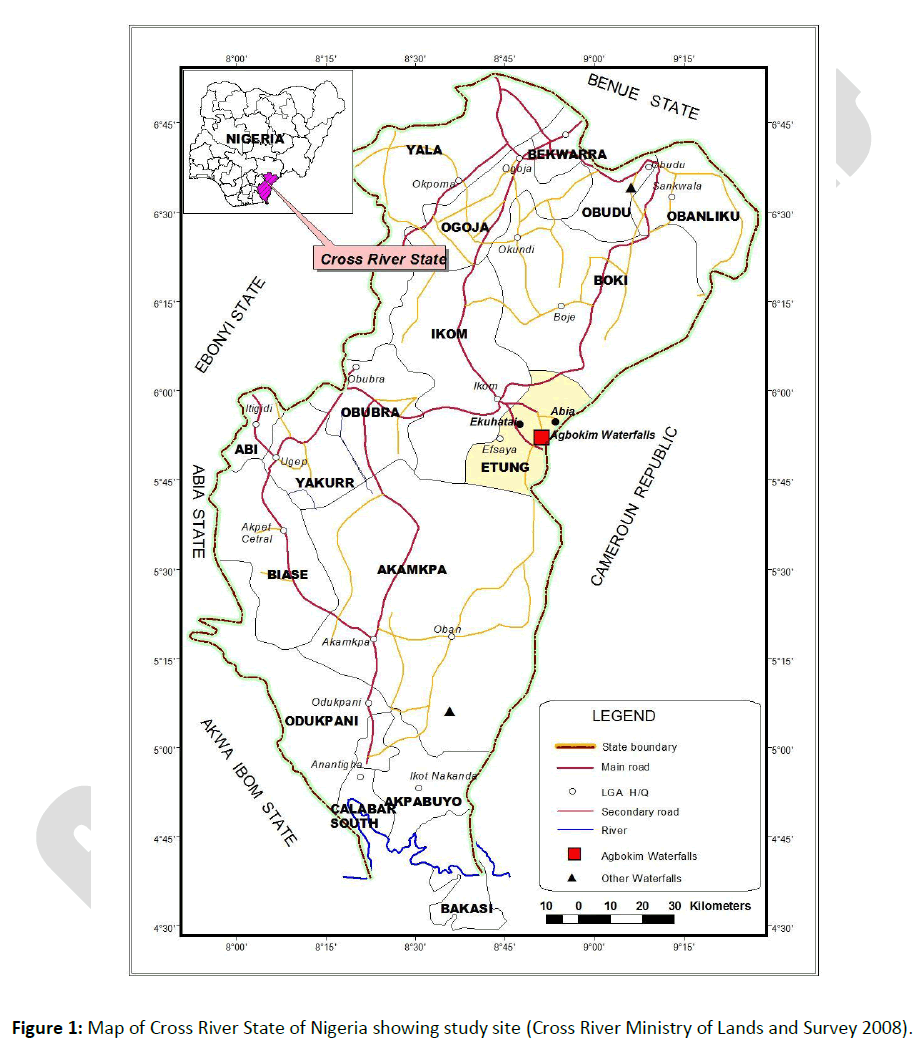
Figure 1: Map of Cross River State of Nigeria showing study site (Cross River Ministry of Lands and Survey 2008).
Sampling description
An average of twenty fish was sampled from each of the three reaches. Fish samples were collected once every month for 24 months (Jan 2007–Jan 2009), from 10 randomly selected fishers in each reach. The fishing gears comprised mainly seine net (10–34 mm, stretched mesh size) and gill net (22–76 mm stretched mesh size). Distribution pattern of the fish in the study area was investigated in the field. Fish samples were preserved in 10% formalin prior to laboratory examination.
Morphological measurements
In the laboratory, data obtained from each fish included; length, weight, sex, fecundity, and food records. Standard length (SL) and total length (TL) were measured to the nearest 0.1 cm and weighed (wt) to the nearest 0.1 g. Samples were identified using FAO Species Identification Sheet and sexed by visual observation of the gonads [24–27].
Stomach content
A total of 150 live specimens for diet studies were dissected and their guts removed immediately after capture and stored in formaldehyde solution (4%) until the contents were analyzed [28]. Gut analysis was later carried out, food items identified to the lowest possible taxon and analyzed quantitatively for percentage composition by number (N) and frequency of occurrence (FO) according to [29,30]. Percent composition by number is the percentage of the number of food items examined accounted for by selected taxa. The FO describes the percentage of fish with non-empty gut that contained at least one of a selected food item.
Fecundity
Analysis of fecundity was limited to the peak spawning period (May–July) and only sexually matured female fish (494) were used for the estimation. Ovaries were removed from body cavity of each fish, weighed and preserved in Gilson fluid [31]. This method of preservation hardened the eggs, break down ovarian tissues and liberate the eggs. Ovaries of each fish were preserved for 7 days before the eggs were counted. The preserved ovaries were washed to remove the preservative. Eggs were separated from tissue and placed on filter paper to remove excess water before being weighed using Metler P 1210 chemical balance. Only the largest matured eggs (2.0–3.0mm), gonadal ripe stage (VI) in each sample were used for fecundity estimation. Eggs in a 1-gram sub-sample were counted. Counting was done for five similar sub-samples. The mean number of eggs in the five sub-samples gave the number of eggs per gram of weight. Fecundity was calculated by multiplying the total weight of eggs by the number of eggs per gram weight. Relative fecundity (RF) = No. of eggs per unit length (cm).
Data treatment
Fecundity-length and fecundity–weight relationships were determined using the expression by King [51].
F = axb
F = absolute fecundity, x = independent variables (body weight, total length). a = scaling constant representing the intercept, b = allometric coefficient both of which were evaluated by least squares regression analysis using
log transformed data; log F = log a + b log xi [32].
Fulton’s condition factor (CF) was determined using the expression [33],
K= W 100
L3
K= condition factor, W = total weight (g) and L =total length (cm).
Diet breath estimates the diet spectrum and was calculated using the diversity index of Shannon-Wiener (H); H = -Σ pi × ln pi, where, pi is the proportion by the number of food type i, Food richness; expressed using Margalef’s index: d= (S-1)/logN), where d = Margalef’s index, S is the number of species and N is the number of individuals [34] and Gut Repletion Index (GRI) which is the percentage of non-empty stomach was estimated for samples from each reach according to Hynes [35].
Statistical analysis
Data collected were collated and analyzed using descriptive statistics (mean, standard deviation, and percentage). Statistical comparison of data between and within reaches were carried out using analysis of variance (ANOVA) and line graphs using MS Excel statistical package (2007). Linear regression was employed to determine the type of relationship between any given pairs of variables. Correlation analysis was used to ascertain the significance of these relationships. The variability in data were evaluated using the coefficient of variation (CV) and the F-ratio test [36].
Distribution and relative abundance
Analysis of species distribution and catch rate in Agbokim Waterfalls showed that L. coubie was more abundant than other fish species in the Cyprinidae family and contributed 27.3% in the overall fish catch during the study (Table 1). The distribution between the sampling sites showed that downstream had the highest representation by number of 38.34% followed by 12.1% midstream and3.1% upstream. The relative seasonal abundance of L. coubie showed that more adult species were common in the catch between the months of August–October representing 72.8% and least (3.1%) were recorded in the months of March, April, and May. The months of August–October revealed more juveniles and fry with 65.6% while the months of February, March, May, and June recorded least (4.3%) (Table 2).
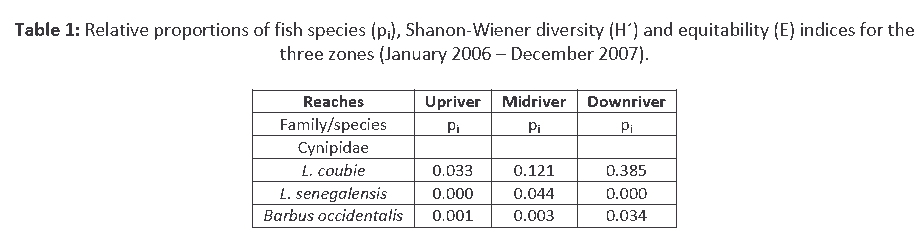
Table 1: Relative proportions of fish species (pi), Shanon-Wiener diversity (H´) and equitability (E) indices for the three zones (January 2006 – December 2007).
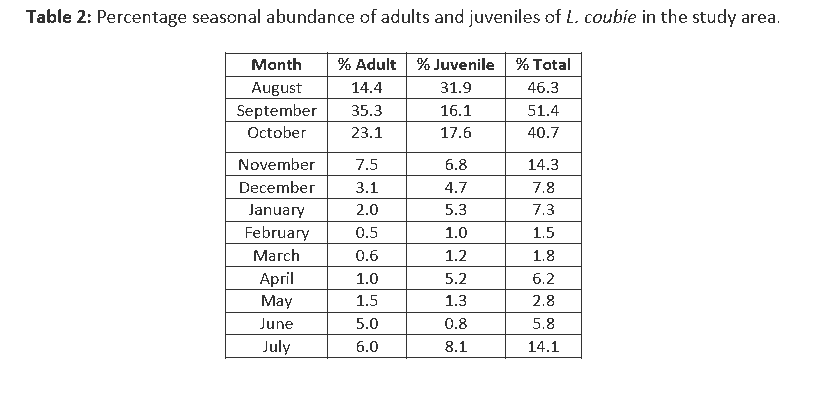
Table 1: Relative proportions of fish species (pi), Shanon-Wiener diversity (H´) and equitability (E) indices for the three zones (January 2006 – December 2007).
Table 2: Percentage seasonal abundance of adults and juveniles of L. coubie in the study area
Sex ratio
There was no significant difference (P > 0.05) in the male: female sex ratio between study sites, and between seasons for L. coubie. Sex ratio as pooled from the study area was 1:0.84. Upstream showed 1:0.92 male:female ratio, midstream 1:0.83, and downstream 1:0.82. Male dominance was observed in the species over the entire study period except in the months of April, November, and December when females were more (Table 3).
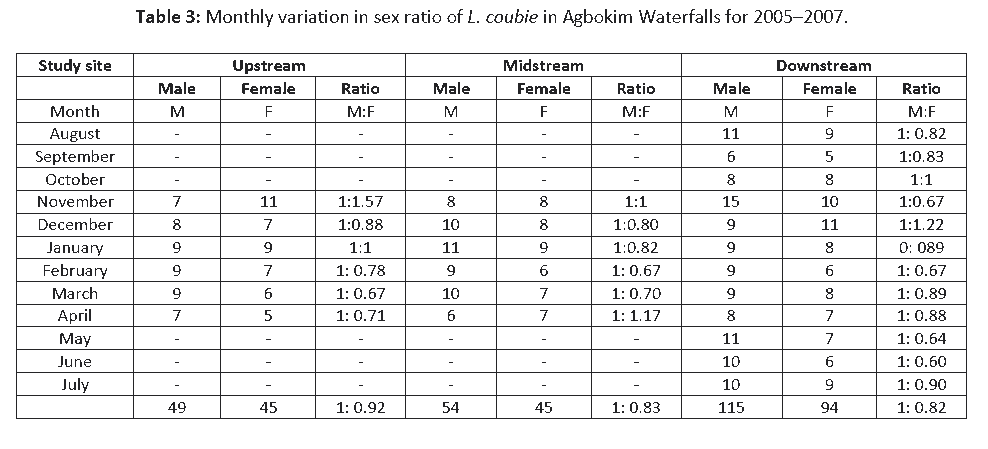
Table 3: Monthly variation in sex ratio of L. coubie in Agbokim Waterfalls for 2005–2007.
Reproductive biology
The study of fecundity of 184 specimens of L. coubie revealed that the relative fecundity ranged from 208 to 236 eggs cm-1 and 25–33 eggs gm-1. Relationship between fecundity and total weight, ovary weight, ovary length, and body breadths, etc. was found to be positively linear (Table 4). F= fecundity, TL = Total Length, SL = Standard Length, TW = Total Weight, OL = Ovary Length, OW = Ovary Weight, BB = Body Breadth. In the present study, it was found that the numbers of eggs increased linearly with the increase of body weight, body length, gonad weight, and gonad length. All the relationships were found to be linear (t-test of all the relationships showed that the values of the regression, coefficient ‘r’ was significant). The results of the fecundity values of L. coubie also showed significant difference (P < 0.05) between study sites with relative fecundity values of 198 eggs cm1 upstream, 229 eggs cm-1 midstream, and 254 eggs cm-1 with overall mean of 206 eggs cm-1.
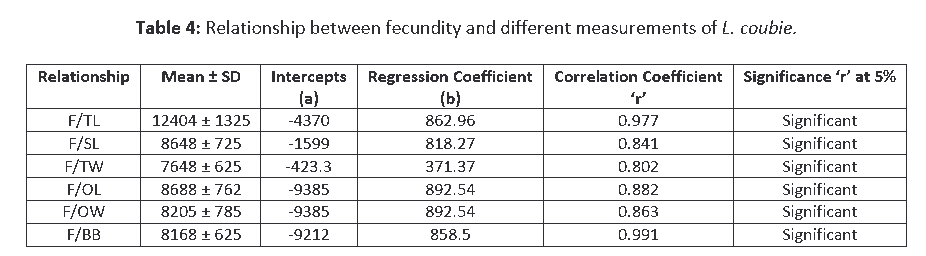
Table 4: Relationship between fecundity and different measurements of L. coubie.
Food and feeding habit
Of the 150 samples of L. coubie examined for food and feeding habit in the three reaches throughout the period of study, three were empty. The results revealed L. coubie as a detritivore but these results overlap with other feeding habits. The results also showed that L. coubie had the least food richness of all the dominant fish species in the study area with food richness of 7. Table 5 presents the food items in the stomachs of L. coubie, using numerical method and FO. In the numerical method detritus, phytoplanktons and plant materials formed the major food items in the diet of fish upstream with percentage occurrence of 36.8%, 32.1%, and 17.2%, respectively. The fish midstream had values of 32.7%, 32.7%, and 19.2%, respectively, while 31.9%, 36.9%, and 16.0%, respectively, were dominant items downstream.
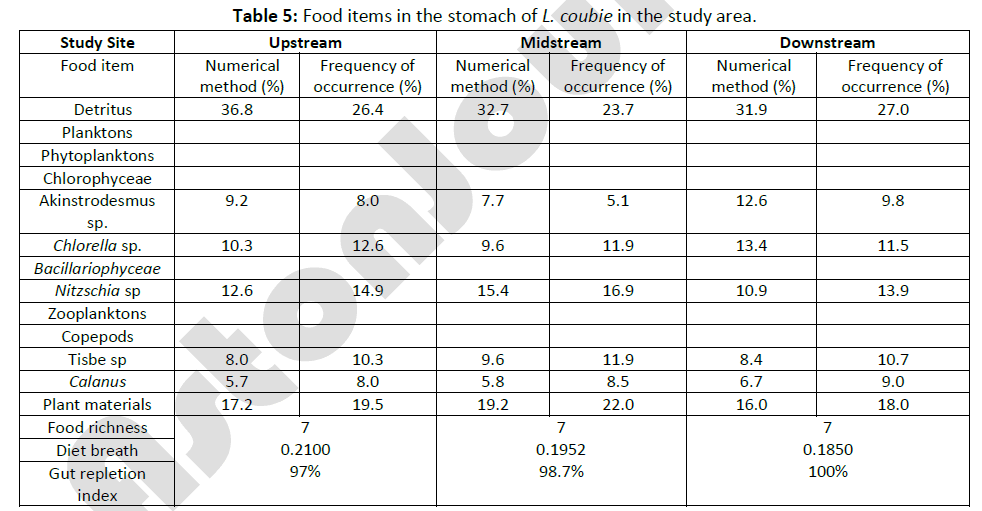
Table 5: Food items in the stomach of L. coubie in the study area.
Length–weight relationship (LWR)
Sizes of both male and female L. coubie ranged from 9.0–22.2 cm standard length (SL) and 38.2–619.6 g total weight (TW) with no significant difference between sexes, and overall patterns of length–weight relationships as:
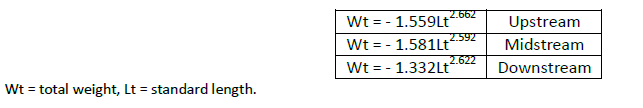
The annual mean length exponents (allometric coefficient) for fish from the three study sites were 2.662 ± 0.291 (upstream), 2.592 ± 0.368 (midstream) and 2.622 ± 0.315 (downstream) (Figure 2) with an overall mean of 2.625 ± 0.335 with coefficient of variability (CV)= 46.1–48.2% for the three study sites. There was significant correlation between length and weight in the study sites (upstream: 0.928 ± 0.039, midstream: 0.936 ± 0.036 and downstream: 0.922 ± 0.048).
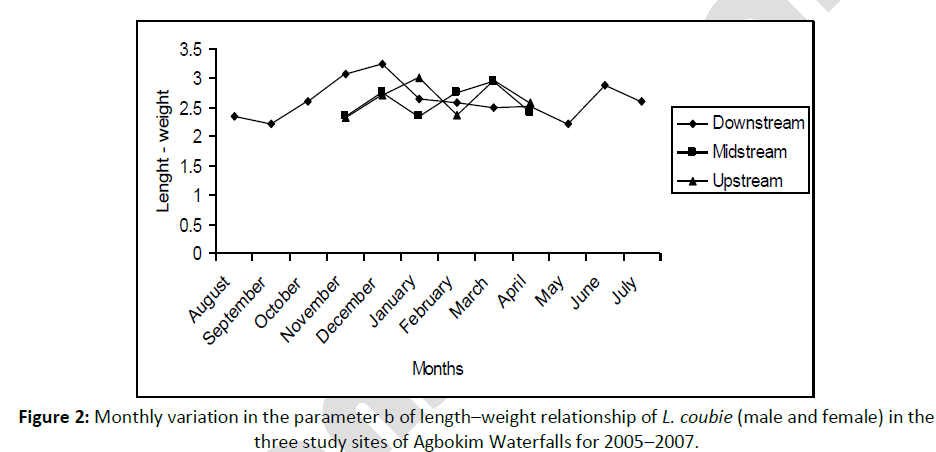
Figure 2: Monthly variation in the parameter b of length–weight relationship of L. coubie (male and female) in the three study sites of Agbokim Waterfalls for 2005–2007.
Seasonal variations in allometric coefficient (b) of the LWR of L. coubie for the two seasons and in the three study sites were significant (P < 0.05) (Figure 3) with overall pattern of the LWR as:
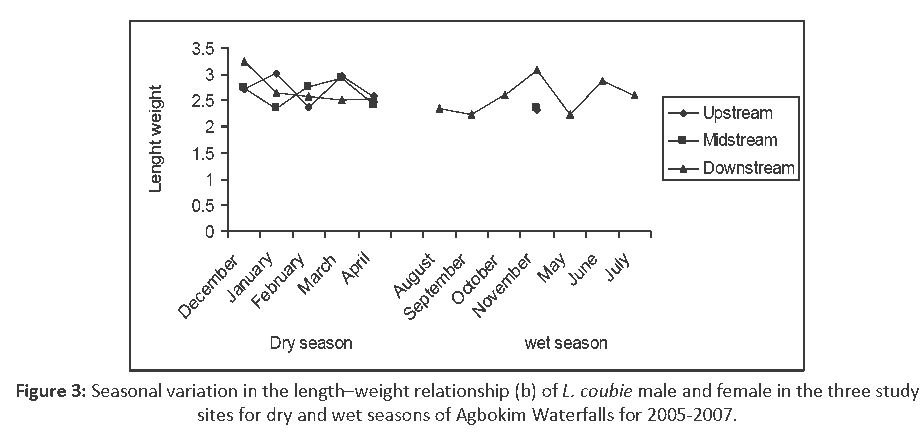
Figure 3: Seasonal variation in the length–weight relationship (b) of L. coubie male and female in the three study sites for dry and wet seasons of Agbokim Waterfalls for 2005-2007.
Condition factor ‘K’
Monthly variation in the condition factor ‘K’ for L. coubie (combined male and female) ranged from 0.509 to 0.915 with annual mean 0.642 ± 0.127 upstream, 0.634 ± 0.106 midstream and 0.648 ± 0.146 downstream with overall mean 0.641 ± 0.152 (0.6 ± 0.2) (Figure 4). The seasonal variation of the condition factor ‘K’ for dry and rainy season ranged from 0.509 to 0.772 with seasonal mean 0.613 ± 0.118 (upstream), 0.635 ± 0.118 (midstream) and 0.663 ± 0.161 (downstream) for dry season, while rainy season values are 0.786 ± 0.166 (upstream), 0.625 ± 0.147 (midstream) and 0.903 ± 0.002 (downstream) (Figure 5).
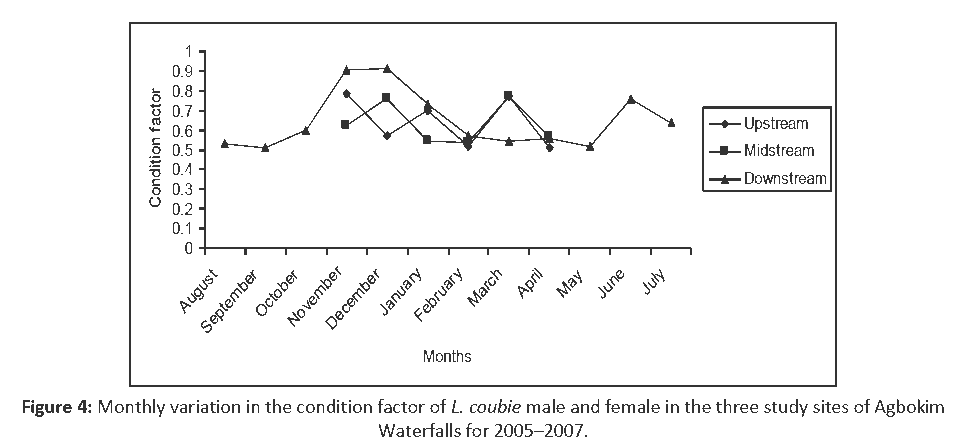
Figure 4: Monthly variation in the condition factor of L. coubie male and female in the three study sites of Agbokim Waterfalls for 2005–2007.

Figure 5: Seasonal variation in the condition factor of L. coubie male and female in the three study sites for Dry and Wet seasons of Agbokim Waterfalls for 2005–2007.
4. Discussion Analysis of the sex ratio of L. coubie showed an overall 1:0.84 male:female ratio with no significant difference (p > 0.05) between sampling sites. This result is not in agreement with some other species studied in the country for Synodontis schall in Zaria, Nigeria [37] for Cynoglossus species of Qua Iboe [38] and Cross River Systems of South East Coast, Nigeria for Macrognathus pancalus [39], who had more females than males but with a few exception of some monthly male dominance. Sex ratio divergence might also be explained by partial segregation of ripe forms either through preference school formation, hence rendering one sex most vulnerable to capture [40]. It is important to note that the more males than females recorded in this study could be favorable to the fishery because it can serve as a regulatory mechanism for the sex ratio [41]. This may be due to the fact that the gears were not set close to the breeding grounds. It has been pointed out that in African water bodies it is common that in populations of fish, the males dominate because they generally present more growth than females without them representing a risk situation for the fishery. Fecundity results showed highest values occurred from fish downstream and lowest from fish midstream. The results from the study area are however similar with that recorded in other studies [41,42]. Results have shown that some environmental properties especially soft water were responsible for the bursting of bighead carp Hypophthalmich thysnobilis and thus suggested it as a factor that could limit the spread of this species [43]. It was also noted that hatching rate and egg size, were significantly affected by the different water qualities [42]. It is possible however that some environmental properties especially the high water turbulence midstream, with its ecological influence downstream can have negative impact on fecundity which could have been responsible for overall low fecundity values in the study area. Maximum mean fecundity value of 23648 ± 721.358 in the waterfalls was very low compared with 110,005 ± 12,007 eggs from L. coubie in Cross River, but higher than 3576–3897 eggs recorded for same species in a reservoir at Ilorin [43] and 679–12906 eggs for species from Cross River Estuary [44].
Dietary habits based on stomach content analysis, is widely used in fish ecology as an important means of investigating trophic relationship in the aquatic communities [45,46] and in the creation of trophic models as a tool to understanding complex ecosystems [47]. Similar investigations in the tropics had shown that the natural food tend to vary quantitatively and qualitatively with seasons of the year [48,49]. L. coubie had been described as detritivore because of the fish’s preference for detritus, although with some overlap [25,50]. In this study, detritus was dominant food item, followed by phytoplankton. The high values of phytoplankton in the stomach content, presents L. coubie also as planktivore. This assertion had been justified by the report that when a fish is feeding, it is the most available food item that it feeds on. This could account for the large number of phytoplankton in the stomachs [51]. Gut Repletion Index of 97–100% indicates that the fish species is a regular feeder. Low values of diet breadth showed that the species does not exhibits trophic flexibility, which is an ecological advantage that enabled the fish to switch from one food category to another in response to fluctuation in their abundance [45]. This probably explains the low Food Richness Index. Results of the length–weight relationship indicated that specimens of L. coubie exhibited negative allometry in the three study sites. The population of L. coubie in the three study sites can therefore be considered as having homogenous groups with body weights varying indifferently from the cube of total length. Seasonal variations in the length–weight relationship (LWR) for L. coubie also indicated negative allometry (b < 3.0). The negative allometry exhibited by the species may be as a result of the hydrological, ecological and human factors. The slightly higher b values downstream may be as a result of the stable ecological nature of downstream. Many authors have reported both isometric and allometric growth for different fish species from various water bodies. Allometric growth patterns for Tilapia species from Umuoseriche Lake had been reported [51]. Also isometric growth for Pseudotolithus elongatus from QuaIboe estuary was reported [52]. Isometric growth pattern for E. fimbriata had also reported from Cross River estuary in Nigeria [53]. The b value for L. coubie (2.6) in this study was almost the same with L. mrigala (2.657) in Kaptai Lake, Bangladesh, but different from isometric value (3.08) for L. coubie from tributaries of the Volta River, Ghana [52]. Others include b values of 3.22 for L. parvus and 3.07 for L. senegalensis representing isometric growth in the Volta River, Ghana. Seasonal variation was not significant (p > 0.05) [52].
Variation in the condition of L. coubie from Agbokim waterfalls was not significantly different between sampling site. The values were however lower than 2.66 obtained for L. boga and 2.77 for L. coubie (43) from a reservoir in Nigeria. The fact that 70% of the samples examined had condition factor above the mean condition factor for the entire population indicate that they are efficient in finding food [54]. Condition factor K reflects through its variations, information of the physiological state of the fish in relation to its environment. From a nutritional point of view, there is the accumulation of fat and gonadal development [55]. From a reproductive point of view, the highest K-values are reached in some species [56]. Condition factor gives information when comparing two populations living under certain feeding, density, climate and other conditions; when determining the period of gonadal maturation; and when following up the degree of feeding activity of a species to ascertain the feeding efficiency [57]. It had been reported that low K-values during the more developed gonadal stages might mean resource transfer to the gonads during the reproductive period [58]. Also, it has been shown that values of the condition factor vary according to seasons and are influenced by environmental conditions [59].
Low diet breadth in this study shows that L. coubie in the waterfalls will not be able to switch from one food to another in response to availability. Downstream provides better spawning condition for L. coubie in the tropical waterfalls ecosystems.
The authors declare that they have no competing interest.
GU carried out the experimental design and drafted the manuscript; AJ conducted fecundity experiments and drafted the manuscript while BO identified some species and performed statistical analysis.
The research was supported by a grant from the Cross River State Government of Nigeria. We are grateful to staff of the Department of Fisheries and Wildlife of the University of Ibadan for their technical contribution.