Journal of Thermodynamics & Catalysis
Open Access
ISSN: 2157-7544
ISSN: 2157-7544
Review Article - (2015) Volume 6, Issue 2
In this overview an attempt was made in order to delineate the biogeochemical cycling of carbon, nitrogen and phosphorous in the Earth’s system and also to relate their occurrence with the Earth’s system Thermodynamics. These global cycles result in the maintenance of a unique thermodynamic state of the Earth’s system which is far from thermodynamic equilibrium. The transfer of matter and energy between the Earth’s systems reservoirs is studied in relation to processes using solar radiation to conserve energy e.g. photosynthesis. The generated chemical energy is used to sustain life, but also to provide additional free energy to geochemical reactions, thereby altering their rates.
Keywords: Nutrients; Biogeochemistry; Carbon; Nitrogen; Phosphorus; Thermodynamics
The term “biogeochemical cycles” is used in order to describe the transfer and transformation of matter and energy between the biosphere and the other active reservoirs of Earth namely the atmosphere, hydrosphere, and lithosphere [1]. During the functioning of a biogeochemical cycle, a series of molecular species which are essential for supporting life on the planet, also referred to as nutrients, are being constantly transported and chemically transformed. The main chemical elements making up the basic nutrients circulating within a specific biogeochemical cycle and which are involved in the structuring of complex bio-polymers (e.g. proteins, DNA, RNA, etc.) are C, N, S, P, and O. A biogeochemical cycle is activated via coupling of the biosphere to nutrient fluxes supplied by specific sites or active centers on Earth (e.g. volcanoes and hydrothermal fields). Such sites/centers act as a constant source of matter (e.g. volatile chemical species) and/ or energy on the Earth surface, either on a regional or a global scale, which can be used by different living forms [1]. The biosphere, being the top level ecosystem, requires numerous nutrients for its function and thus it has established a series of complex interactions with the different Earth reservoirs. These interactions are regulated by both negative and positive feedback control mechanisms and constitute the different biogeochemical cycles [2]. It is thus deduced that the efficient function of a biogeochemical cycle is strongly dependent and constrained by the inflow and outflow of specific nutrients through critical sites or active centers of the Earth. Such sites/active centers are justifiably considered to serve as vital “planetary organs” whose disintegration would lead to collapse of the biogeochemical cycle [3]. As a self-protective action, the biosphere possesses some of these vital organs in the form of specialized trophic groups which are guided by autotrophic organisms that occupy specific habitats of the planet and lead to energy storage via the production of complex biomolecules. Since the maintenance of a biogeochemical cycle depends on processes involving energy flow and storage, it is deduced that it can be described by using the principles of thermodynamics [1,4]. Earth systems are not isolated (closed systems) since there exist strong interactions between the atmosphere, hydrosphere, lithosphere and biosphere. The numerous thermodynamic processes which take place can be viewed and separated into those that continuously perform work and create free energy and those that dissipate free energy and continuously produce entropy. Entropy production is a direct measurement of the degree of irreversibility of a thermodynamic process and it is the driving force for directing a thermodynamic system to adopt and be maintained in a state far from thermodynamic equilibrium (TE). The possibility of the Earth, as a thermodynamic system, to sustain a state away from TE can be resolved when considering the fact that Earth may exchange entropy with space [4]. Thus, it seems that non-equilibrium thermodynamics could provide a holistic basis for describing and predicting the emergent, far from TE, thermodynamic state of the Earth as one system as well as of different subsystems, e.g. environmental or ecological systems, [5]. Using this perspective as a starting point, one may consider examining the possible role of life, existent in the biosphere, in directing and maintaining a biogeochemical process in a steady state far from TE [5-8].
Carbon is present in two forms: organic and inorganic. The biochemical and geochemical cycling of these forms through the Earth processes is referred to as the global carbon cycle. The global carbon cycle is closely linked to the global cycles of other elements such as oxygen, nitrogen, and sulfur. Organic carbon (OC) is present in both living and nonliving systems providing the basic fuel for all of the Earth’s biogeochemical processes [9]. Therefore, understanding the formation, cycling and preservation of OC is essential for understanding the cycling of all elements which are biologically relevant, the formation of economically-exploitable oil (petroleum) and coal deposits, and for predicting the influence of human activity on the natural balance of these systems.
Much of the OC created during photosynthesis by microorganisms (e.g. cyanobacteria) and higher plants or by chemosynthetic autotrophs is decomposed by heterotrophic organisms. Heterotrophic processes occur in the water column of marine and aquatic environments, as well as in sediments, soils, and rocks. Much of the decomposition of organic matter (OM) occurs under low temperature (<50°C) and pressure during diagenesis.
The production of organic matter during photosynthesis is described by the following equation:
6CO2+6H2O+chlorophyll+sunlight→C6H12O6+6O2 (photosynthesis)
ΔG0=2870 KJ mol-1
The reaction known as respiration, which describes the oxidation of organic carbon leading to the production of carbon dioxide and water and the release of energy contained in the chemical bonds making up the OC, is given by the equation:
C6H12O6+6O2→6CO2+6H2O+Energy (respiration)
The cycling of carbon in a global scale due to anthropogenic activities (Figure 1) also involves OC fluxes to and from the major OC reservoirs (Figure 2). Sedimentary rocks constitute the largest OC reserve, making up approximately 99.5% of the total global OC. However, OC only represents one-fifth of the total sedimentary carbon, since inorganic carbon is the dominant fraction [10]. Sedimentary OC occurs primarily as kerogen, coal beds, and petroleum reservoirs, which are ultimately derived from biogenic processes or sources. Kerogen is an amorphous, highly insoluble particulate material that is formed by two pathways - namely selective preservation and geo-polymerization both of which occur under low temperature and pressure conditions [11]. Selective preservation involves the formation of kerogen from bio-macromolecules that are resistant to microbial degradation. Geopolymerization occurs through a random series of condensation and polymerization reactions which involve humic substances. Petroleum is defined as a naturally occurring fluid (mixture of liquid and dissolved gases) and which is formed primarily from the remains of microbes (algae and bacteria), although some types of petroleum are formed from the remains of higher plants. In contrast, coal is a solid and is formed from the remains of higher plants.
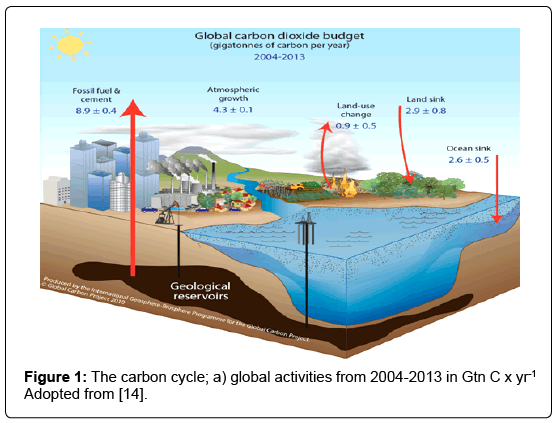
Figure 1: The carbon cycle; a) global activities from 2004-2013 in Gtn C x yr-1 Adopted from [14].
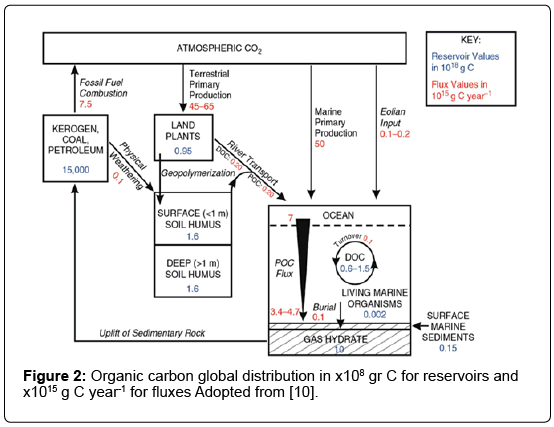
Figure 2: Organic carbon global distribution in x108 gr C for reservoirs and x1015 g C year-1 for fluxes Adopted from [10].
The most common type of coal, known as humic coals, is formed via the processes of peatification and coalification, which involve both biological and geochemical processes [11]. The OC presence in ancient sedimentary rocks provides these economically important fossil fuels as well as a molecular record of life, which has allowed to study the response of natural systems to tectonic, environmental, and biological change [12]. Sedimentary OC affects biologically mediated cycles on geologic time scales, although there is evidence that these cycles seem to be accelerated by humans through fossil fuel combustion [13].
The major fluxes between organic pools are driven by marine and terrestrial primary production. Photosynthesis, the harvesting of light energy and subsequent utilization to convert inorganic carbon to reduced (organic) forms in the tissue of plants, is the primary mode of primary production. However, chemosynthesis by microbial organisms, which uses chemical energy rather than sunlight, is also important. Photosynthesis is carried out by prokaryotes such as cyanobacteria as well as eukaryotic algae and higher plants. The fixation of carbon by plants accounts for the presence of molecular oxygen in such high concentration in the atmosphere. Thus, the cycles of carbon and oxygen are closely linked over geologic time [14-16].
In order to maintain current atmospheric oxygen levels, rapid cycling of carbon in the surface pools is required. These pools exchange carbon on relatively short timescales (102-104 years). In contrast, the OC that remains in the sedimentary system is cycled slowly (about 108 years [1]. Sedimentary reservoirs become exchangeable only through geologic uplift, followed by oxidative weathering of OC by exposure to the atmosphere. Net global Primary Production (NPP) on land (45–65 × 1015 g C year-1) is dominated by more labile non woody plant tissues such as leaves, grasses, and herbaceous annuals [1]. The highest gross terrestrial OC production (82 × 1015 g C year-1) and storage as biomass (56%) occur in the tropics, while polar regions contribute the least (8 × 1015 g C year-1, 0.5%; [17]. Within these regions, forests on one hand and deserts and tundra on the other, are the most and least productive respectively [17], thus reflecting global patterns of temperature and precipitation. Marine primary productivity (about 50 × 1015 g C year-1) is dominated by open water (pelagic) phytoplankton productivity because open water constitutes approximately 75% of the total ocean area [18,19].
However, humans may have reduced total NPP by up to 40% due to wildfires and pollution, which may be the largest sink for primary production to support a single species in the history of life on the Earth [20]. Fossil fuel combustion and destruction of forests (especially the tropical ones) have decreased the global pool of OC and increased atmospheric carbon dioxide, which acts as a greenhouse gas. Global atmospheric concentrations of CO2, methane (CH4), and nitrous oxide (N2O) have increased markedly as a result of human activities since preindustrial times (i.e., 1750). In 2005, the concentrations of atmospheric CO2 (379 ppm) and CH4 (1,774 ppb) exceeded the natural range over the last 650,000 years [13]. Understanding the changes in the size and composition of the OC reservoir through space and time will help us to better predict the changes in its global cycle as well as the associated cycles of other biologically important elements.
Over the last century, the impact of human activity on the nitrogen cycle has become global [21]. At the beginning of the twentieth century most reactive nitrogen in soil was produced bacterially amounting to about 100 million t yr-1. At the beginning of the twenty-first century, anthropogenic nitrogen production amounts to ca. 160 million t yr-1 [22], mostly as fertilizer (more than 100 million t), with much of the rest being added to the biosphere inadvertently as a by-product of the burning of fossil fuels for transport, industrial and domestic purposes. With the current rate of nitrogen production [21] reports that global production of reactive nitrogen could reach between 250 million and 900 million t yr-1 by 2100.
A major problem arises from the fact that the efficiency with which organisms pick up nitrogen in the soil is low (generally in the range 25 to 50%), with the excess leached and washed from the soil predominantly in the form of nitrate [24]. The resulting massive introduction of nitrogen into the natural system creates problems in a wide variety of terrestrial and marine ecosystems [25-27]. Eutrophication, recognized as an endemic problem in the surface waters of agricultural areas of the developed world in the 1960s and 1970s, has now extended to the marine environment, causing hypoxia (so called “dead zones”) in near shore waters.
However, the path of nitrogen in the biosphere can be viewed as a cycle of reactions through which elemental nitrogen enters into the soil from the atmosphere, undergoes a series of transformations, and eventually comes out of the soil and re-enters the atmosphere. The complete process is collectively known as the nitrogen cycle (Figure 3). The important steps in the nitrogen cycle are nitrogen fixation, mineralization, immobilization and volatilization each of which is mediated by different microorganisms.
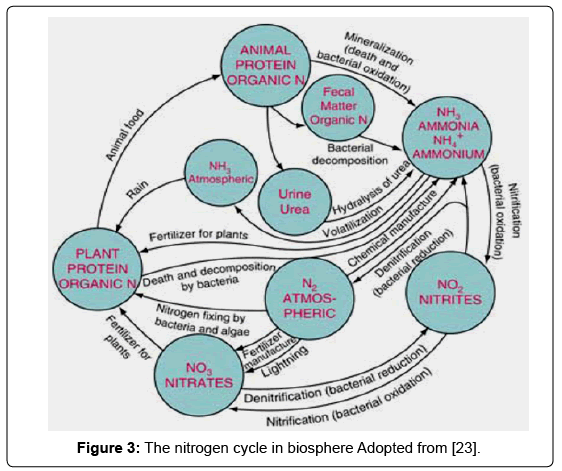
Figure 3: The nitrogen cycle in biosphere Adopted from [23].
The ultimate source of soil nitrogen is atmospheric nitrogen. About 1 and 10 kg ha-1 of atmospheric N2 is transformed to the ammonium and nitrate forms by electrical discharges during thunderstorms and washed into the soil through precipitation (Figure 4). In addition N2 is converted into organic nitrogenous compounds by certain microorganisms, with or without the association of a host. Mineralization is brought about by two distinct processes both of which involve the metabolic reactions of microorganisms. The first process known as ammonification, breaks down organic nitrogen compounds to NH3 or NH4+ with the initial step being the enzymatic breakdown to amines (called proteolysis or aminization). In the second process, known as nitrification, NH3 or NH4+ is oxidized to NO2- or NO3- (the latter being the predominant available form of nitrogen in well-aerated, cultivated soils). Nitrification is used by several organisms as an energy source [28]. The following reactions describe the overall energy yielding steps:
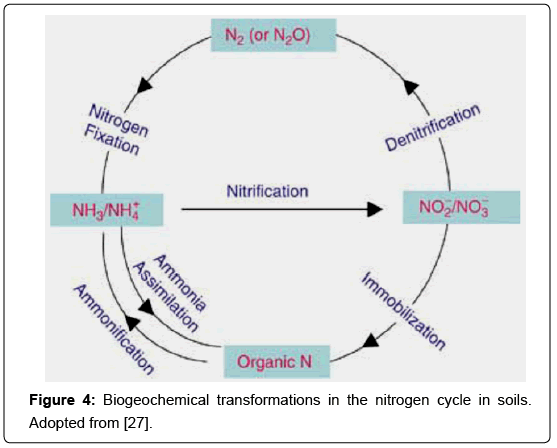
Figure 4: Biogeochemical transformations in the nitrogen cycle in soils. Adopted from [27].
NH4++1.5 O2→NO2-+H2O+2H+, (mediated by Nitrosomonas)
ΔG0=-290 KJ mol-1
NO2-+0.5O2→NO3-, (mediated by Nitrobacter)
ΔG0=-82 KJ mol-1
Although nitrogen gas (N2) is about three quarters of the atmosphere (approximately 386 × 1013 t), it constitutes a limiting factor for the growth of plants and animals because nitrogen cannot be assimilated by most of the organisms in its elemental form. Plants utilize mostly soil nitrogen (present mostly as inorganic ammonium and nitrate ions) for protein synthesis while animals use plants as their main nitrogen source.
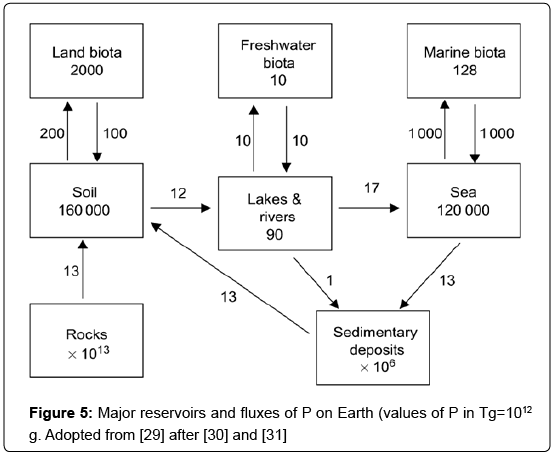
Phosphorus comes in 10th place in the order of elemental natural abundance in the Earth's crust, with an average concentration of about 1 g kg-1. The various compartments and estimated annual fluxes of global P are shown in Figure 5. Sediments and rocks constitute the largest P pool. The global P cycle can be delineated in the primary inorganic cycle, on one hand mineral weathering, leaching and erosion transfer P from land to sediments, and on the other volcanism and uplifting of sediments replenish terrestrial P, thereby completing the cycle [29].
This cycle has a frequency in the order of measured in millions of years. The two biological cycles involve movements of P with short frequencies (in the order of years and even weeks) and take place in both the land-based and water-based biospheres. There is an annual flux of 4.3 Tg P yr-1 between land and atmosphere, and a flux of 1.4 Tg P yr-1 from atmosphere to ocean [30]. Those parts of the natural cycle are small and do not significantly alter the data presented in figure 5. Richey's [30] estimate of the other global P fluxes is quite similar with that of Pierrou [31] which has been employed in figure 5. The highest flux occurs between the sea and marine biota, followed by the flux between the soil and its vegetation. However, the human impact on the global P cycle has been substantial over the last 150 years. Since this anthropogenic modification was in effect well before the onset of scientific efforts to quantify the cycle of P, one can only guess at the “pre-anthropogenic” mass balance of P. Several aspects of the P cycle are well-constrained in figure 6. Phosphorus is initially solubilized, mainly from apatite minerals, by chemical weathering during soil development. Physical weathering also plays a role by producing fine materials with extremely high surface area/mass ratios, which enhances chemical weathering in continental environments (i.e., floodplains, delta systems) [32].
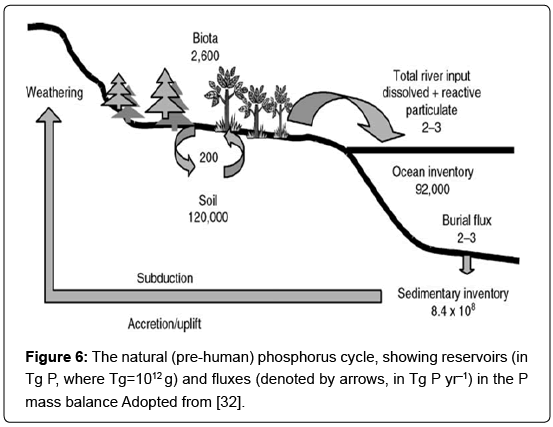
Figure 6: The natural (pre-human) phosphorus cycle, showing reservoirs (in Tg P, where Tg=1012 g) and fluxes (denoted by arrows, in Tg P yr–1) in the P mass balance Adopted from [32].
The cycling of P in soils has received much attention, in terms of both fertilization and the natural development of ecosystems. Of the approximately 122,600 Tg P within the soil/biota system on the continents, nearly 98% is held in soils in a variety of forms (Figure 6). The exchange of P between biota and soils is relatively rapid, with an average residence time of 13 years, whereas the average residence time of P in soils is 600 years. The P contained in the apatite minerals of the soil is the type which is most easily weathered via its reaction with dissolved carbon dioxide as in the following equation:
Ca5(PO4)3OH+4CO2+3H2O→5Ca+2+3HPO4 2-+4HCO3-
In soils, P is released from mineral grains via several processes. Firstly, the reduced pH produced from respiration-related CO2 in the vicinity of both degrading organic matter and root hairs dissolves P-bearing minerals (mainly apatites) and releases P to root pore spaces. Secondly, organic acids released by plant roots can also dissolve apatite minerals and release P to soil pore spaces [33]. Phosphorus is very immobile in most soils, and its slow rate of diffusion from dissolved form in pore spaces strongly limits its supply to rootlet surfaces. Furthermore, much of the available P in soils is in organic matter, which is not directly accessible for plant nutrition. In order to increase the supply of P to their roots, plants employ the enzyme phosphatase and the fungi Mycorrhizae Phosphatase is an enzyme that is often excreted by plants and soil microbes and which can release bioavailable inorganic P from organic matter. The symbiotic fungi Mycorrhizae can coat plant rootlets, excreting phosphatase and organic acids to release P and providing an active uptake site for rapid diffusion of P from soil pore spaces to root surfaces [1]. The phosphorus content in plants, normally ranging between 0.1% and 0.4% of the dry weight (depending on the stage of plant development and plant part) is considerably less than that of nitrogen and potassium, the other two major nutrients. The average total P content of the soil is about 430 mg kg-1, the lowest among the major essential plant nutrients [34]. In contrast, soils contain on average 2000 and 15000 mg kg-1 of N and K, respectively. Therefore most agricultural soils require the addition of P fertilizers to achieve the expected crop productivity. It is obvious that the P cycle of the biosphere is meaningful only when viewed across terrestrial and aquatic ecosystems. Detailed knowledge of the P budget of terrestrial ecosystems, especially agro-ecosystems, grasslands and forestry-based systems, is needed to assure sustainable management of those landbased resources and the environment.
The circulation of nutrients like carbon, nitrogen, phosphorous between living and nonliving compartments on the Earth is defined as cycling. During cycling there are many processes like the metabolic pathways of the organisms as well as chemical reactions and geological processes that delineate the transformations of the nutrients. These transformations promote the energy transfer between the different reservoirs of the Earth’s system [3]. The framework of reservoirs is strongly constrained by the inflow and outflow of the nutrient through critical locations where the energy is collected, transformed, or derived to another reservoir [3]. In the biosphere, the energy collectors supply nutrients to the rest of their biological components. However, what are the general principles that govern the way in which biosphere, atmosphere, hydrosphere, geosphere interactions regulating the Earth’s system functioning? It is a fact that the interactions between the different Earth’s systems happen spontaneously. This aspect can be understood and quantified in general terms of thermodynamics. In specific the second law of thermodynamics is about to explain this aspect of the system’s function.
In this term, entropy can be used as a measure for the lack of gradients and free energy. By depleting gradients and sources of free energy, these processes are directed towards the state of Thermodynamic Equilibrium (TE) at which the entropy of the system and its surroundings is maximized. In this state, matter is well mixed and no free energy is available to perform physical work or run chemical reactions [4]. That the Earth system is not in a “steady state”, seems like a violation to this general trend and thus to the second law of thermodynamics. But why the Earth’s system function maintained far away from thermodynamic equilibrium To prevent such a “steady state” of Earth, some processes need to create gradients and sources of free energy in a setting that does not violate the second law.
During these processes (e.g. the creation of nutrients) physical and chemical work is needed in order to keep global cycling of matter, (e.g. organic matter production – consumption). In such a “living” state of the Earth system, these processes can be viewed and separated into those that continuously perform work and create free energy, and others (the irreversible ones) that dissipate it and continuously produce entropy [4]. One critical aspect that distinguishes Earth’s system from the ideal – isolated systems in thermodynamic equilibrium is that Earth systems strongly interact. To keep these cycles running, energy exchanges with space are needed, while in a planetary scale we can safely neglect mass exchange with space although there may be some climatic effects of, e.g. cosmic rays on clouds. But what are the engines which continuously operate to keep this system running? In a global scale the production of organic matter, the water cycle as a well the atmospheric circulation and nutrients cycling are the main engines that keep the Earth’s system functioning. These engines ultimately run on gradients of radiative fluxes, which result from the vastly different radiative temperatures of the Sun (Tsun ≈ 5760 K) and Earth (Tearth ≈ 255 K). Gradients in radiative fluxes can result in gradients of heating, which drive heat engines such as the atmospheric circulation or photochemical engines that make use of low entropy solar radiation directly (Figure 7).
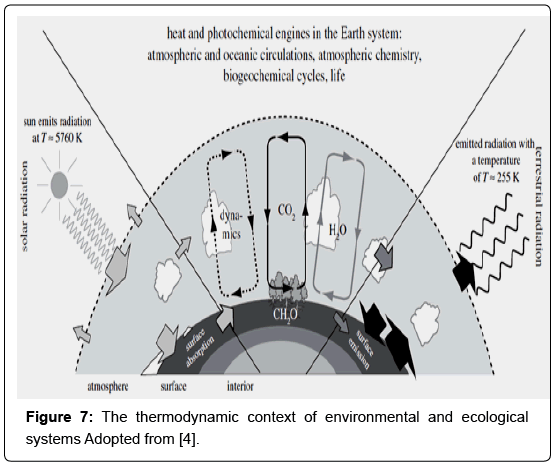
Figure 7: The thermodynamic context of environmental and ecological systems Adopted from [4].
In this sense, spatial and temporal differences in the absorption of solar radiation provide the radiative forcing gradients that drive the atmospheric heat engine, which provides the means to transport chemical elements within the Earth system. That is, the atmospheric circulation continuously performs work, and the dissipated heat is rejected to space in the form of long-wave radiation. Hence, in total, the absorption of solar radiation and the subsequent emission of terrestrial radiation to space provide the ultimate driving force to operate Earth systems far away from TE [4]. Solar radiation also provides the photochemical energy to drive photosynthesis, which in turn provides the major source of free energy to drive geochemical cycles within the Earth system. The heat generated by the consumption of free energy is again ultimately rejected to space in the form of long-wave radiation. This state far from TE is then reflected into an atmospheric composition with 21% reactive oxygen and an atmosphere that is mostly nonsaturated with vapour [4]. This notion of an atmospheric composition far from TE in turn has been interpreted as a sign of widespread life on Earth and resulted in the formulation of the controversial Gaia hypothesis [35-37].
However, non-equilibrium thermodynamics would provide a basis to understand the emergent state of environmental and ecological systems far from TE [5-8]. At the Earth’s system (far from TE) the exchange of energy or mass, within the surroundings (biosphere, atmosphere, hydrosphere, geosphere) of the system conform to the second law of thermodynamics. The second law states that the total entropy increase, that is, the entropy increase of our system dS/dt plus the increase in entropy in the surroundings dSsurr/dt, can only increase. Thus the term ‘entropy production’ (σ) refers to the increase of entropy of the system because of a certain, irreversible process (e.g. mixing by heat exchange), so we can write the entropy balance using this terminology in the form of [4]:
dS/dt=σab
(dS/dt)+(dSsurr/dt) ≥ 0
In other words, while the local system can evolve away from TE, the system plus its surroundings evolve closer (and faster) to TE. The entropy increase of the surroundings is equivalent to the Net Entropy Exchange (NEE) between our system and the surroundings, so that we extend our entropy balance to:
dS/dt=σab-NEE
with the entropy production σab given by:
σab=Fab ((1/Tb)-(1/Ta))
Where Fab is the heat flux between the surrounding and the surrounding b with temperatures Ta and Tb respectively.
In steady state where (dS/dt=0), entropy production σab is equal to the NEE, so that entropy fluxes across the boundary can be used to compute entropy production in steady state. The net entropy exchanges NEEa and NEEb of surroundings a and b are given by the exchange fluxes of heat and the respective temperatures at which heat is exchanged [4]:
ΝΕΕa=((Fc+Fab)/Ta)+(Fout, a/Ta)-(Fin/Tin)
ΝΕΕb=(Fout, b/Tb)-((Fc+Fab)/Ta)
For the whole system, the NEE is:
NEE=(Fout, a/Ta)+(Fout, b/Tb)-(Fin/Tin)
Where, Fin is the influx into surrounding a, Fc is a conductive flux from surrounding a to surrounding b, Fab is the flux by an alternative process, and Fout,a and Fout,b are the heat losses from the surrounding a and b to the environment.
Hence, it is the entropy exchange across the system boundary that maintains steady states away from TE. Essentially, all processes within the Earth system are associated with irreversibility and entropy production and their entropy production can be represented as the product of a thermodynamic force and flux. During entropy production (σab) given at the above equation, the thermodynamic flux is the heat flux, and the thermodynamic force is the gradient of 1/T [4].
In the case of mass fluxes, entropy production is expressed as the product of a mass flux and the gradient in chemical potential, divided by temperature. In addition there are other pairs of conjugated variables that can perform work like the wind stress and velocity, pressure and volume, and so on. Since in steady state the rate at which work is performed equals dissipation, changes in these conjugated variables result in entropy production [5]. A quantification of entropy production that allows us to compare the relative contribution of irreversible processes to the overall entropy production by the Earth system as a whole is given in figure 8. The white arrows denote fluxes of solar radiation, black arrows terrestrial radiation, grey arrows fluxes of heat, and thin lines material cycling (Figure 8). The boxes describe irreversible processes. The symbols are: F, flux of energy or heat (in Wm-2); Ω, solid angle (in radians); T, temperature (in K); and σ, entropy production (in W m-2 s-1) [38].
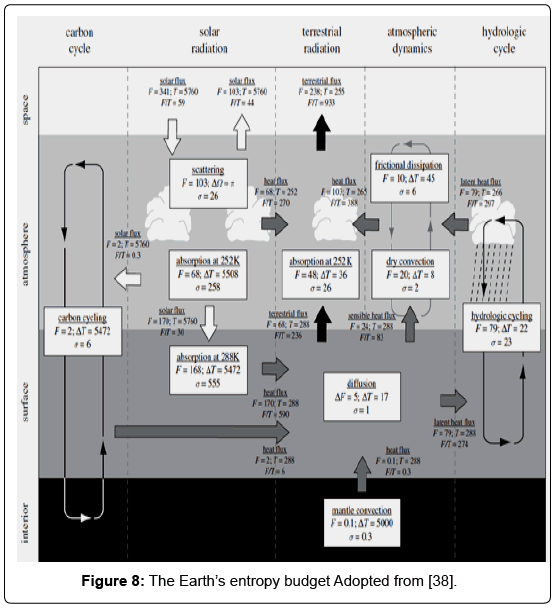
Figure 8: The Earth’s entropy budget Adopted from [38].
It must be noted that biotic activity strongly shapes atmospheric CO2 concentrations and contributes the most entropy production in the budget shown in figure 8. Photosynthesis uses the low entropy of sunlight to drive chemical reactions that remove CO2 from the atmosphere and build up free energy in the form of carbohydrates. When these carbohydrates are respired, e.g. by building or maintaining biomass, or by the activity of heterotrophs, the chemical energy is converted into heat. The irreversibility is associated with the conversion of solar radiation into heat, which can no longer be used to drive photosynthesis [5,38-42].
In the Earth’s system the exchange of energy and/or mass with the surroundings is a critical component that allows systems to evolve away from Thermodynamic Equilibrium without violating the second law of thermodynamics. In this concept cycling of nutrients is related with the necessary energy transformations through biogeochemical processes that affect their occurrence and distribution in surrounding systems like the biosphere, atmosphere, hydrosphere, geosphere. Nutrients are essential to life because they are major constituents of the leaving organisms and also involved to the storage and/or the release of energy. The main biogeochemical cycles involve different molecular species whose composition provides the essential elements for both building polymeric structures and the biochemical transduction of energy in cells, like C, N, and P.
The biogeochemical cycles are activated when the biosphere couples to the nutrient fluxes that are supplied from specific sites or active centers on Earth (e.g. volcanic centers, hydrothermal systems) that are essentially energy sources which maintain the supply of matter and energy to the Earth surface. The biosphere, as the top-level ecosystem, has its own energy transducers, a role played by the primary producers that occupy the different habitats of the planet. They promote the transport of energy in the form of molecular compounds from the lithosphere to the different reservoirs of the Earth. Most of these active systems are powered by the release of the inner heat of Earth, which, in the form of aerial or subaqueous systems, introduce at the regional or global scale different volatiles and other chemical species usable by living forms. Matter and energy cycling is accomplished when the Earth fluxes drive the nutrients and other components to specific sinks where they are generally recycled by biochemical or geochemical mediated reactions to compounds having a different chemical state.
The authors would like kindly acknowledge the anonymous reviewers for their comments and suggestions helped to improve the quality of the manuscript.