Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2012) Volume 3, Issue 1
Inland saline groundwater is abundant in various areas of the world but is not suitable for traditional agriculture irrigation and is also not suitable for marine aquaculture mainly because of potassium deficiencies. In the present work, we used marbled spinefoot rabbitfish Siganus rivulatus, as a model organism to investigate whether varying levels of aqueous K+ affect survival and growth, and whether ambient K+ has an effect on fish osmoregulation and haemolymph osmolality. A ten-week growth experiment was conducted where fish were reared in reconstituted inland saline water containing K+ concentrations equivalent to 15%, 40% and 100% (denoted as: Trt15, Trt40, and Trt100, respectively) of potassium in 15 ppt seawater. A 15 ppt seawater treatment (CSW) was used as a control. Fish weight, length, survival, Fulton-type condition index, blood and water osmolality, and serum Na+ and K+ concentrations were evaluated. Gill NKA activity was also estimated. Fish reared in Trt15 died during week 8. At the end of the experiment (week 10), no differences were observed in fish survival and total length among treatments Trt40 and Trt100 and control (P>0.05). Final weight was less in Trt40 and Trt100 than in CSW. Fulton-type condition index results showed that Trt100 was more suitable for fish than Trt40, but still not as suitable as 15 ppt sea water. Sodium (Na+) in serum was not affected by aqueous K+ levels, whereas serum K+ varied congruently with aqueous K+. Rabbitfish reared in Trt40 and Trt100 had greater blood osmolality than those reared in control seawater. Gill NKA activity also increased by 50% in Trt40 and Trt100 as compared to control. The present study shows that potassium-deficient 15 ppt inland well water is not suitable for long-term culture of Siganus rivulatus. Results suggest that inland saline well waters greatly increase activity of NKA and consequently energy expenditure. Furthermore, K+ does not appear to be the only limiting mineral in inland low salinity groundwater that affects fish osmoregulation.
Keywords: Potassium deficiency, low salinity well water, rabbitfish
Increasing demand for aquatic food products combined with limited production from capture fisheries is responsible for rapid coastal and inland aquaculture development. However, high cost of coastal land, strict regulations, and environmental concerns, have led aqua farmers to favor inland aquaculture. Furthermore, the fact that various areas of the world (e.g. Australia, USA, China, India, Thailand, Brazil, Ecuador) are underlain by saline ground waters not suitable for traditional agriculture [1-3] has greatly increased interest in inland aquaculture of saltwater species.
Inland saline well waters (ISWW) do not have the same ionic composition as marine or brackish waters [4] and are not suitable for survival and growth of marine species, unless chemically modulated. Saline ground waters are generally deficient in the key ion potassium (K+) [5,6]. Adding a potassium source such as muriate of potash (KCl) and K-Mag to pond waters in order to increase aqueous K+ concentrations generally improves performance/production of marine species [7-9].
As the most abundant intracellular ion in all animals, potassium plays basic physiological functions including maintenance of membrane potentials and cellular volume, acid/base balance, cardiac function and nerve impulse transmission. In fish, potassium plays additional critical roles in osmoregulation [10]. Some robust osmoregulators such as silver perch Bidyanus bidyanus [11,12] and rainbow trout Oncorhynchus mykiss [11] grow well in non-chemically modified inland saline groundwater. However, potassium deficiency was shown to negatively impact the performance of various other high market value aquaculture species such as Pacific white shrimp Litopenaeus vannamei [8,13-15] Australian snapper Pagrus auratus [7,16], barramundi Lates calcarifer [17-19], western king shrimp Penaeus latisulcatus [20], tiger shrimp Penaeus monodon [9,11,21], mulloway Argyrosomus japonicus [22,23] and giant prawn Macrobrachium rosenbergii [24]. Such results suggest that K+ availability in ISWW is necessary for survival and growth of aquatic organisms, yet the physiological mechanisms and effects of potassium deficiency on fish are not properly understood. Some research suggests the involvement of other aqueous ions such as Mg+2 and SO4-2 [4,25] on survival and growth but we could not find any that specifically discuss the effects of other ions.
A model for the NaCl extrusion by the marine chloride cells (CCs) was described in detail by Hirose et al. [26]. Gill Na+/K+ ATPase (NKA) abundant in the baso-lateral plasma membrane complex of the CCs is a key enzyme in osmoregulation [26,27]. It is thought that modification of aqueous Na / K ratios affect the activity of the NKA [4]. Sampaio and Bianchini [28] suggested that reduction in growth of fish reared in non-optimal salinities could be partially due to an increase in energy expenditure associated with a higher NKA activity. This negative correlation between fish growth and NKA activity might be one of the reasons for the reduction in survival and growth of fish reared in low K+ saline ground water.
Siganus rivulatus [29] is an herbivorous marine teleost that is suitable for aquaculture [30,31]. S. rivulatus grow and survive well in salinities between 10 and 50 ppt with an optimum salinity of 35 ppt [32], and are quite eurythermal (17 °C to 32 °C) with an optimum temperature for growout of 27 °C [33]. Like most euryhaline teleost species, Siganus rivulatus tolerates a wide range of salinities by modifying the activity and/or expression of gill NKA [32,34-39]. In the present study we assessed survival and growth of the marine euryhaline S. rivulatus in low salinity inland-like potassium deficient well waters and studied effects of varying levels of aqueous K+ on NKA activity and fish osmoregulation.
Fish Acquisition and Holding
The present work was performed at the American University of Beirut (AUB) aquaculture research laboratory. Rabbitfish juveniles were caught in traps off the coast of Beirut and transported to AUB. Fish were acclimated from 36 ppt to 15 ppt seawater in a 1m3 quarantine tank, and transferred to the experimental culture system. The acclimation process consisted of gradual decrease of salinity over ten days at a rate of 2 ppt per day, followed by a two-week acclimation period at endpoint salinity 15 ppt. For the growth experiment, we used twelve 180 L square fiberglass tanks (60 cm L x 60 cm W x 50 cm H). Water was recirculated into trickling biological filters and settling sump tanks before returning to the culture tanks. Aeration was provided via a regenerative blower and submerged air diffusers. Water temperature was maintained at an average 27.2 ± 0.06 °C (mean ± SE) using 300 W submersible heaters placed in the filter sump tanks. Average pH 8.08 ± 0.03 (mean ± SE) was controlled using sodium bicarbonate. In addition to natural daylight, fluorescent lighting provided a 14 L: 10 D light regime. Fish were offered a commercial trout feed (Zeigler Bros Inc.) to apparent satiation three times daily. Salinity, water temperature, dissolved oxygen concentration, and pH were measured daily. Total ammonia nitrogen (TAN) and nitrite nitrogen (NO2-N) concentrations were measured twice weekly using the Solorzano [40] and Parsons et al. [41] methods, respectively. Ammonia-N and nitrite-N concentrations remained below 0.04 mg/L and 0.2 mg/L, respectively.
Experimental Design
The growth experiment was conducted over a period of ten weeks. Fish were size sorted by hand (7.7 ± 0.55 g, 8.5 ± 0.20 cm; (mean ± SE)) and randomly stocked as groups of 12 fish per tank, with three replicate tanks per treatment. Water in the tanks was formulated to resemble 15 ppt inland saline well water as reported by Saoud et al. [5]. Briefly, 4 ppt seawater was mixed with dechlorinated tap water and salinity raised to 15 ppt by addition of 1.64 g/L MgCl2.6H2O, 1.92 g/L MgSO4.7H2O, 0.02 g/L NaBr, 0.28 g/L CaCO3, and 9 g/L NaCl. The reconstituted water was mixed by vigorous aeration for three days and then allowed to settle for 3 days. A sample was then analyzed by ICP and chemical makeup found to be similar to typical Alabama inland low salinity waters deficient in potassium [5]. KCl was added to two treatment waters at concentrations of 100 and 160 mg/L K+, theoretically equivalent to 62.5% and 100% of 15 ppt seawater, respectively. However, potassium concentrations in the waters as measured by ICP were 93.89 mg/L and 224.6 mg/L, equivalent to 40.3% (Trt40) and 96.4% (Trt100) of potassium in 15ppt seawater, respectively. No potassium was supplemented to the third treatment which contained 35.6 mg/L potassium, equivalent to 15.3% of K+ concentration (Trt15) in 15 ppt seawater. The fourth treatment consisted of 15 ppt seawater and was used as a control (CSW) (Table 1). At the end of the experiment, fish were harvested, individually weighed and individual total length recorded. Five fish from every tank were randomly taken to measure plasma osmolality and gill tissue NKA activity.

Figure 1: Representative photomicrograph of histopathological features in pulmonary necropsies.
Plasma and Water Osmolality
Fish were anaesthetized using MS-222 (Tricaine Methanesulfonate, 15 mg/L) and blood was extracted from the caudal arch. Blood samples were centrifuged at 3,000 rpm for 3 min and plasma used for osmolality measurement. Water samples from the various treatments were also collected for osmolality estimation. Osmolality was estimated using a Wescor® vapor pressure osmometer (Vapro 5520).
Blood Serum Concentrations of Na+ and K+
Serum was obtained by centrifugation of coagulated blood samples in 2 mL Eppendorf® micro-centrifuge tubes at 4,000 rpm for 10 minutes at 4 °C. Serum was acid digested (65% HNO3, 30% H2O2) using Milestone ETHOS Plus Medium Pressure digestion microwave (12 digestion Teflon vessels, TFM). Blank and samples were spiked with K+ and Na+ solutions (20 mg/L). Concentrations of Na+ and K+ were estimated using flame photometry (Cole Parmer Digital Flame Photometer, Vernon Hills, IL, USA). Calibration of the machine was performed using standard solutions of K+ and Na+ prepared by serial dilution of respective stock solutions (1000 mg/L).
Gill Tissue Na+-K+-ATPase Activity
Fish were killed by giving them a strong blow to the head and then inserting a needle in the brain. Gills were removed and the third and fourth caudal holobranchs were separated from their corresponding arches, and homogenized on ice in 1 ml Histidine buffer (150 mM, pH 7.4) for 4 minutes at medium speed (26,000 rpm), using a 7 mm POLYTRON® homogenizer. Aliquots were stored at -70 °C.
NKA activity was determined by measuring the level of Inorganic Phosphate (Pi) released by the gill chloride cells, using a modified colorimetric assay described by White [42]. Briefly, gill tissue samples were diluted with histidine buffer (150 mM, pH 7.4) to a concentration of 2 μg/μL protein. One percent saponin was added at a ratio of 1:3 v:v. Serine/threonine phosphatase inhibitor (30 μL) was added and the whole mixture was incubated at room temperature for 30 minutes. Gill samples (30 μL) were incubated for 10 minutes at 37 °C, in three Eppendorf® micro-centrifuge tubes containing the substrates of the NKA enzyme (25 μL KCl (200 mM); 25μL NaCl (1240 mM); 25μL MgCl2 (40 mM)) and 3 tubes containing choline chloride (75 μL). ATP (30 mM; 25 μL) was added to all tubes and incubated for 30 minutes at 37 °C. Reaction was stopped by adding an equal volume of Trichloroacetic acid (TCA, 50%) and tubes were centrifuged at 3,000 rpm for 5 minutes at 4 °C. Supernatant was added to color reagent Molybdate stain (5% ferrous sulfate FeSO4, 10% ammonium molybdate [(NH4)6Mo7O24·4H2O], and 0.5M sulfuric acid H2SO4). Reaction rate was assessed spectrophotometrically by measuring absorbance at 800nm. NKA activity was assayed by calculating the difference in Pi released in presence and absence of the Na+/K+-ATPase substrates, according to the following formula:
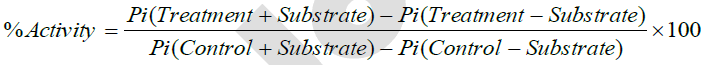
Protein concentration in gill homogenates was determined using the BIO-RAD protein assay (Bio-Rad Laboratories, USA) with bovine serum albumin (0.8 mg/mL BSA) as a standard. Absorbance was determined at 595 nm using a spectrophotometer (Thermo Electron Genesys 20); regression analysis was performed using GraphPad software and protein concentrations were obtained from the generated standard curve.
Data Analysis and Statistics
Statistical analyses were performed using SPSS 16.0. The Fulton-type condition index (K) of the fish was calculated as follows: CI= 100,000 W/L3, where W= fish weight (g) and L= total length (mm). Effects of treatments on survival, weight, length, NKA activity, and osmolality were analyzed using one-way ANOVA (P<0.05). Student-Newman- Keuls (SNK) and Dunnett t test (2-sided) were used for mean separation.
Fish Survival and Growth
All fish reared in Trt15 died at the beginning of week 8. However, no mortality was observed in Trt15 during the first 7 weeks. Fish reared in 15 ppt control seawater, Trt40 and Trt100 survived until the end of the experiment (week 10) with no significant differences (P>0.05) in survival among treatments (Table 2). There was also no significant effect (P>0.05) of aqueous K+ concentration on total length among treatments. However, weight of fish reared in Trt40 and Trt100 was significantly less (P<0.05) than that of fish reared in control seawater. Final condition index of fish reared in Trt100 was significantly greater than K of fish reared in Trt40, but both had a K significantly less than control condition index.

Table 2: Initial weight Wt(i), final weight W(f), initial length L(i), final length L(f), initial Fulton-type condition index CI(i), final condition index CI(f), and survival (S) of rabbitfish Siganus rivulatus reared in 15 ppt control sea water (CSW), Trt15, Trt40 and Trt100. Data represent mean values.
Plasma and Water Osmolality
Plasma osmolalities of fish reared in Trt40 and Trt100 for 10 weeks (417.8 and 426.5 mmol/kg, respectively) were significantly greater (P<0.05) than osmolality of fish reared in 15 ppt control seawater (383.4 mmol/kg). Water osmolalities of CSW, Trt40 and Trt100 were 432, 440 and 439 mmol/kg, respectively, and were not significantly different from each other (Table 3).

Table 3: Tank water osmolality (mmol/kg), mean plasma osmolality (mmol/kg) and standard error of mean (SE) of blood osmolality of rabbitfish Siganus rivulatus (n = 15) at the end of a 10 week growing period in 15 ppt control sea water (CSW), Trt40 and Trt100.
Blood Serum Concentrations of Na+ and K+
Flame photometry analyses of digested blood serum showed no significant differences (P>0.05) in serum Na+ concentrations among all reared fish. However, potassium concentration in Trt40 was lower (P<0.05) than serum potassium concentration of fish in Trt100 and in 15 ppt seawater control fish (Table 4). Serum potassium was similar in Trt100 and control fish.
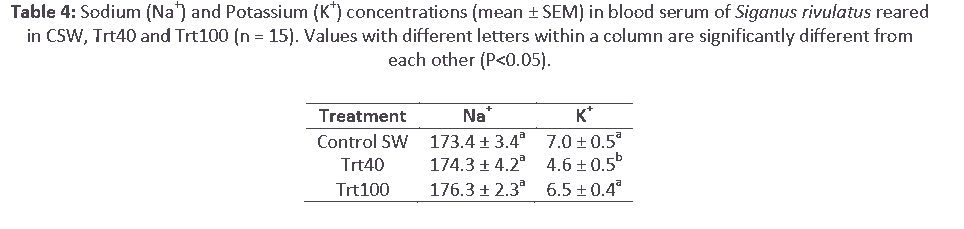
Table 4: Sodium (Na+) and Potassium (K+) concentrations (mean ± SEM) in blood serum of Siganus rivulatus reared in CSW, Trt40 and Trt100 (n = 15). Values with different letters within a column are significantly different from each other (P<0.05).
NKA Activity
Gill Na+-K+-ATPase activity of fish in Trt40 and Trt100 was greater than that of gill NKA activity in control fish (P<0.05). NKA activity was not significantly different among treatments Trt40 and Trt100. Percent increase (compared to control) in gill pump activity was 45.8% for fish reared in Trt40 and 49.5% for those reared in Trt100 (Table 5).
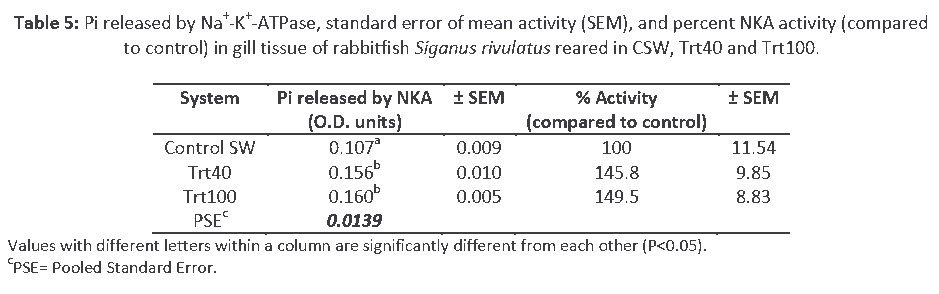
Table 5: Pi released by Na+-K+-ATPase, standard error of mean activity (SEM), and percent NKA activity (compared to control) in gill tissue of rabbitfish Siganus rivulatus reared in CSW, Trt40 and Trt100.
Results of the present study suggest that potassium-deficient 15 ppt low salinity well water is not suitable for long-term culture of Siganus rivulatus. When potassium is supplemented to ISWW, survival and growth of S. rivulatus improves. Various workers have reported poor survival and/or growth of marine organisms when reared in ISWW, and improved survival and/or growth when K+ was supplemented to the water [7,9,19, 20,22,24]. A few teleost species such as silver perch and rainbow trout have demonstrated good survival and growth in unmitigated ISWW [11,12] , but these are freshwater species and thus do not get much potassium from surrounding waters anyway. However, for marine species accustomed to environmental potassium, deficiency of K+ in the water affects osmoregulation and survival. Partridge and Creeper [17] hypothesized that fish tolerance to K+ deficiency is inversely proportional to salinity, a hypothesis supported by Jain et al. [18] and Partridge and Lymbery [19] who reported similar findings when studying effects of aqueous K+ on barramundi. Similarly, Zhu et al. [43] found that Pacific white shrimp Litopenaeus vannamei were more tolerant to abnormal Na/K ratios at low salinities (15 ppt) than at higher salinities (30 ppt). According to Partridge and Lymbery [19], the lower requirement for K+ at 15 ppt is either because of an improved uptake and/or because of reduced loss of the ion at this salinity. Also, fish cultured in near iso-osmotic salinities theoretically lose less water compared to those living in hyperosmotic salinities, and absorb fewer monovalent ions through diffusion. Consequently, there is a decrease in NaCl excretion across the gills which results in reduced loss of K+ ion that usually occurs concomitant to NaCl excretion [44]. In the present work, fish were maintained at their salinity of isosmolality [32] and thus effects of potassium deficiency in the water would have been reduced. Nonetheless, in Trt15 potassium concentration must have been less than the tolerance limit of the fish even at the salinity of isosmolality.
The fact that some species of fish tolerate low aqueous potassium when reared at isosmotic salinities whilst other species do not suggests that requirement for K+ is species specific. For example, Partridge and Lymbery [19] stated that barramundi Lates calcarifer has greater tolerance for inland saline groundwater than other teleost species investigated. Nevertheless, similar trends of tolerance to K+-deficiency in ISWW are observed among various species. For example, poor performance of fish in 15%-K at near iso-osmotic salinity (15 ppt) observed in our study and improved performance at 40.3%-K are corroborated by results of Doroudi et al. [22] working with mulloway Argyrosomus japonicus. They found that survival was least in waters containing 25% of the K+ concentration of seawater, and optimal conditions for growth and survival, over 44 days, were in waters containing K+ in excess of 40% of seawater equivalence. Similarly, 25%-K was insufficient for the survival of the Australian snapper Pagrus auratus at 20 ppt [7] but K+ concentrations above 40% seawater did not affect snapper survival. That rabbitfishes in Trt15 survived for as long as eight weeks is possibly because at small sizes fish were capable of maintaining homeostasis but as body size increased, homeostasis became more difficult to maintain. However, the idea that small fish are more capable of maintaining osmotic homeostasis than large fish remains to be proven. Most reports suggest that 40% K-equivalence seems to be the lower tolerance level of marine teleosts and 40.3% K-equivalence was found suitable in the present work. Further investigations using intermediate K equivalence levels (ranging between 15% and 40%) are required for determining the minimum tolerable K+ concentration of S. rivulatus at 15 ppt ISWW salinities.
Fish growth can be expressed as a change in length or weight. In our study, rabbitfish grew in weight and length in all treatments (even in Trt15, before death). Final length of fish in the various treatments was similar to fish length in the control thus suggesting that growth in length was not compromised by potassium treatments at or above 40.3 %. No difference in weight was observed among fish in treatments Trt40 and Trt100; however, fish reared in Trt100 did grow less than fish in control seawater (having the same K+ concentration). This difference in weight between fish in 100% K+ in ISWW and those in brackish seawater was not observed for Australian snapper by Fielder et al. [7] or in barramundi by Partridge and Lymbery [19]. The fact that fish in the present study grew better in the control 15 ppt seawater than in ISWW with equivalent potassium content suggests that minerals other than K+ might be limiting growth. Similar findings were reported by Prangnell and Fotedar [20] while studying the effects of aqueous potassium on shrimp growth. They found that Penaeus latisulcatus reared in 30 ppt ISWW with 100% and 80% K+ concentration as compared to seawater resulted in slower growth and reduced molt increment. Although Fielder et al. [7] observed no differences in Pagrus auratus growth when reared in seawater or ISWW with equivalent K+ content, their ISWW had a total ionic profile similar to seawater (i.e. content of Sodium, Chloride, Calcium, Sulfate, Magnesium, Fluoride, Phosphates, Aluminum, Boron, Copper, Iodide, Iron, Lead, Lithium, Manganese, Mercury, Molybdenum, Strontium, and Zinc were similar to those found in equivalent salinity seawater). In the present study, the major ions in the reconstituted waters were similar to those in 15 ppt seawater except for Calcium, Boron, and Strontium. Various researchers report an effect of Ca++ deficiency on osmoregulation and on Na+ concentration in fish blood [46]. The Ca++ concentration in trt100 water was less than in CSW but was not so low as to affect the fish dramatically. Marine waters generally do not contain a lot of Ca and the reason our CSW contained a high concentration of Ca was that the seawater was diluted with high hardness dechlorinated tap water. Consequently, although potassium supplementation did improve survival and growth, it was not enough to mimic performance in seawater and suggests the existence of other limiting factors, possibly minor trace elements such as Sr and B. Our results suggest that a better understanding of fish performance in ISWW requires an evaluation of effects of various elements in addition to K+. Saoud et al. [4] found that shrimp reared in ISWW in Alabama were affected by sulfate and magnesium as well as potassium and Wudtisin and Boyd [25] observed an effect of magnesium deficiency on shrimp survival and growth.
Weight to length relationships, such as Fulton-type condition index, are usually used to evaluate fish physiological health or ‘well-being’ [47]. Improvement of fish condition in the present experiment is probably because of the high-protein feed offered compared to the less nutritional algae usually consumed in nature [48]. At the end of the experiment, condition index was greater in Trt100 than in Trt40 reflecting better feed conversion or smaller loss of energy by the fish in ISWW with greater potassium concentration. However, the better CI observed in fish reared in 15 ppt CSW reinforces the aforementioned hypothesis that the K+ added to the water was a necessary but not sufficient supplement for ISWW to become conducive to good siganid growth.
Aqueous K+ concentration in Trt100 was equivalent to CSW and greater than in Trt40. Blood serum potassium concentration in Trt40 was less than in Trt100 and CSW, suggesting that plasma K+ concentration is dependent on water K+ concentration. Partridge and Lymbery [19] found that when barramundi were reared in 15 ppt ISWW, blood plasma concentration of Na and K did not differ among treatments. But again, because barramundi is a fresh water species, we are not able to draw any conclusive comparison. In marine teleosts, K+ is ingested with food and water rather than passing across the gills, and K+ efflux is not affected by K+ concentration in the water [49]. Accordingly, fish reared in Trt40 did not have an increased K+ efflux, but rather a reduced K+ influx by drinking low-K+ water. Feed used in the experiment was probably not enough by itself to reestablish homeostatic K+ levels.
Blood sodium was not affected in Trt40 and Trt100. However, because blood K+ was affected in Trt40, the Na/K ratio was disrupted. Gill NKA activity increased by 50% in fish reared in experimental water compared to control seawater but no difference in gill NKA activity was observed between fish in Trt40 and Trt100. These results suggest that Na/K ratio was not the only factor disrupting NKA activity. As hypothesized earlier in the discussion, variations in Ca++ concentrations might also be affecting osmoregulatory physiology of the fish. A driving force for most osmoregulatory mechanisms is NKA and an imbalance in the concentration of many ions is likely to affect the activity of NKA. That NKA activity levels in both treatments were not significantly different from each other can be attributed to one of two reasons; 1- either NKA reached a maximum activity (saturation kinetics) in Trt40, or 2- K+ deficiency in this treatment was not actually affecting osmoregulation. However, considering the low blood K and condition index observed in Trt40, we believe that the first hypothesis is more probable.
Partridge and Lymbery [19] observed no differences in gill NKA activity among fish reared for 4 weeks in 15 ppt ISWW with 25%, 50% and 75% K+ equivalence. Fish gill NKA in the present experiment were compared at 10 weeks and by then, those at 25% equivalence had died. Partridge and Lymbery [19] proposed that the direct physiological mechanism responsible for death in juvenile barramundi reared in low K+ (25%) water for protracted periods is skeletal muscle necrosis/myopathy induced by unsustainable buffering of blood potassium levels from the muscle. Such an effect was not tested in the present work. Also, Partridge and Lymbery [19] did not compare their fish to a control seawater treatment at similar salinity. Accordingly, we cannot know if barramundi would have shown the same response as S. rivulatus in seawater but changes in gill NKA activity showed similar trends in fish of both species reared in ISWW at various K+ equivalencies.
Blood osmolality in teleost fish usually ranges between 280 and 360 mmol/kg, and its regulation in stressful osmotic conditions is mainly species-dependent [45]. Saoud et al. [32] reported that S. rivulatus is a strong osmoregulator that shows no significant variation in blood osmolality when reared in seawaters ranging from 10 ppt to 45 ppt. In the present work, plasma osmolality increased in fish maintained in the reconstituted ISWW as compared to fish in 15 ppt seawater but was similar to culture water osmolality. This suggests that fish might have been maintaining near-isosmolality with culture water in order to reduce energetic loss associated with the increase in NKA activity observed in the reconstituted ISWW. Furthermore, plasma osmolality varied between fish in 15 ppt CSW and fish in Trt100, again corroborating our hypothesis that something other than potassium is affecting osmoregulation in ISWW.
Sodium and Chloride regulation in fish living in hyper or near iso-osmotic salinities is mediated by gill MRC transporters/pumps. The NKA pump maintains an electrochemical gradient across the membrane of MRCs by actively exchanging three sodium ions from intracellular space to the outside for two potassium ions from the extracellular medium inwards [50]. This electrochemical gradient constitutes the driving force for the basolateral Na-K-2Cl co-transporter [51,52] that allows the extrusion of Na and Cl. Na+ ions are driven to the external medium through leaky junctions down the trans-epithelial electrochemical gradient. Removal of extracellular K+ has an effect similar to ouabain regarding the inhibition of the NKA, leading to subsequent inhibition of NaCl extrusion [53-55]. Thus, we suggest that, in Trt15 and Trt40, where levels of water K+ were reduced compared to equivalent salinity seawater, Na and Cl concentrations would have started increasing in the blood, causing blood osmolality to increase, and stimulating osmoregulatory mechanisms in the fish (mainly under hormonal control). However, Na+ concentration in the blood does not seem to have increased but NKA activity did increase. This suggests that the fish were expending energy in order to maintain normal cellular K+ levels and re-establish Na+ concentration gradient across the cells. Increased NKA activity compromises survival and growth. In K+-unsupplemented Trt15, very low K+ levels probably disrupted the Na/K ratio, and we hypothesize that death occurred either because NKA had broken down (i.e. reached saturation kinetics) or when energy reserves in the fish were all drained by week eight. Both hypotheses remain to be proven.
The present study suggests that although K+ is the most limiting osmolyte in ISWW, other minerals are also important and their effects should be evaluated. Presently, we would advise farmers to supplement their waters with low-value potassium sources but also to try to rear freshwater species rather than marine species in ISWW until all deficient minerals are determined and mitigation protocols developed.
NM performed most of the work and wrote the manuscript. JG assisted with water preparation, fish maintenance, stocking, weighing, dissection and analytical tests. SK offered her lab and helped with the biochemical assays and experimental design in general. IPS supervised the work after writing the grant proposal, assisted with statistical analysis, supervised the writing and followed up on the manuscript. The majority of the work was performed in his lab.
The present work was supported by funds from the Lebanese National Council for Scientific Research and from the American University of Beirut Research Board. The authors thank Zeina Dakroub for her help in performing biochemical tests.