Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2016) Volume 6, Issue 1
The essential oil (EO) extracted from the flower of Coreopsis tinctoria Nutt. is a kind of plant-derived natural therapeutics against Cryptococcus neoformans. In our study, the anti-cryptococcal effects of the EO were observed from various aspects, such as minimal inhibitory concentration, minimum fungicidal concentration, germ tube formation, and biomass. The results indicated that the EO had potent antifungal activities on C. neoformans and that it was more effective than the commonly used antifungal agents fluconazole and amphotericin B. To further elucidate the mechanism of the antifungal action, the effect of the EO on the plasma membrane and the mitochondria of C. neoformans were investigated. The lesion in the plasma membrane was detected through flow cytometry and was further verified through the inhibition of ergosterol synthesis. The essential oil caused morphological changes in the cells of C. neoformans and a reduction in the ergosterol quantity. Moreover, mitochondrial membrane potential, mitochondrial ATPase, succinate dehydrogenase (SDH), and malate dehydrogenase (MDH) were detected. The reactive oxygen species (ROS) accumulation was examined through fluorometric assay. Decreased SDH, MDH, and ATPase in C. neoformans cells were also observed in a dose-dependent manner. The above dysfunctions of the mitochondria caused ROS accumulation in C. neoformans. A reduction in cell viability was prevented through the addition of L-cysteine, and this reduction indicates that ROS is an important mediator of the antifungal action of the EO. In summary, EO from C. tinctoria flower showed anti-cryptococcus activity. Its mechanism of action is to disrupt the permeability barrier of the plasma membrane and the mitochondrial dysfunction-induced ROS accumulation in C. neoformans. With better antifungal activity than the commonly used antifungal agents and less possibility to induce drug resistance, the EO can be used as a potential anti-cryptococcal agent.
<Keywords: Coreopsis tinctoria, Anti-cryptococcal, Mechanism, Essential oil
Cryptococcus neoformans is a kind of environmental yeast that causes opportunistic infection [1]. The most common and serious type of cryptococcal disease is pulmonary cryptococcosis, which is uncontrolled and leads to cryptococcal meningitis or meningoencephalitis [2]. Cryptococcosis is one of the main pathogens which infections in patients with human hematological malignancies, immunodeficiency virus (HIV) and therapeutic immunosuppression [3,4]. The mortality of disseminated cryptococcosis is 70%-80% in untreated patients compared with 17% in patients treated with systemic antifungal agents [5]. Many clinically used antifungals (e.g., amphotericin-B, fluconazole, 5-flurocytosine, miconazole, nystatin, etc.) suffer from various drawbacks in terms of toxicity, lack of fungicidal efficacy, high cost, drug-drug interactions, and emergence of resistant strains resulting from frequent usage [6,7]. As useful natural products from medicinal and aromatic plants, essential oils (EO) are known to possess antioxidant, antiviral, antibacterial and antifungal activities [8,9].
Coreopsis tinctoria Nutt., which belongs to the family Asteraceae, is a widely distributed medicinal plant in the Kunlun Mountains of the Xinjiang Uygur Autonomous Region (altitude: approximately 3000 m) in China [10]. C. tinctoria is widely used not only as an ornamental plant but also as functional food in China. It is rich in essential nutrients (e.g., vitamins, selenium, zinc, and amino acids [11,12], etc.) and many phytochemicals (e.g., flavonoids, polyphenols, polysaccharides, saponins, and EO) [13-19]. It has been demonstrated to have many nutraceutical properties, such as those for the management of diabetes, reduction of blood pressure and vasorelaxant effects, α-Glucosidase inhibitory effect and hypoglycemic activity, anticoagulating activity, anti-aging activity, and antioxidant and antiinflammatory activities [20-24]. In recent years, the C. tinctoria flowering top from the Kunlun Mountains in Xinjiang has become a popular health product used as herbal tea or beverage in China. However, to our knowledge and according to a literature survey, no reports have been made yet on the anti-cryptococcus activity and mechanism of EO from Coreopsis tinctoria flowering top. Accordingly, this study aimed to determine the mechanism of action of the EO on C. neoformans.
Plant material
The flower parts of C. tinctoria plants were harvested from the Xinjiang Uyghur Autonomous Region of China in September 2012 and were identified by Prof. Jianwei Hu (College of life science, Tarim University). The No. 1-81 was deposited at the herbarium of the Institute of Traditional Chinese Medicine & Natural Products, Tarim University School of Life Sciences.
Gas chromatography-mass spectroscopy (GC-MS) analysis
Air-dried C. tinctoria flowers (200 g) were subject to hydrodistillation for 4 h using a Clevenger-type apparatus. The EO was dried over anhydrous Na2SO4 and preserved in a sealed vial at 4°C until further use. The EO yield was 0.15% (v/w).
The chemical composition of EO was analysed using GC-MS. The EO (20 μL) was dissolved in acetone (200 μL), and 1 μL of the solution was injected into the GC-MS (Agilent 7890A-5975C, USA). The capillary column was HP-5MS (length = 30 m, thickness = 0.25 μm, and i.d = 0.25 mm). Helium was used as the carrier gas at a flow rate of 1 ml·min-1. The GC column oven temperature was increased from 50°C to 300°C at a rate of 10°C·min-1, with a final hold time of 10 min. Injector and detector temperatures were maintained at 280°C. The EI mode was at 70 eV, mass spectra were recorded in the 30 amu-600 amu range, and the ion source temperature was 230°C. The EO components were quantified by the relative percent peak area of total ion current curve (TIC) from the MS signal and identified by comparing their mass fragmentation pattern with those stored in the spectrometer database using NIST05.LIB and NIST05s.LIB (National Institute of Standards and Technology).
Microorganisms
The C. neoformans strains were purchased from the China General Microbiological Culture Collection Center. The fungal strain cultures were maintained on a Sabouraud dextrose agar (SDA) slant at 4°C. The old cultures were transferred to a fresh slant every two months to avoid a decline in strain viability.
Antifungal activities of EO against C. neoformans
The C. neoformans strains were grown on SDA plates in the dark at 17 ± 2°C for four days. The concentrations of spores were adjusted to 1.0 × 108 CFU/mL using a hemocytometer. The EO (80 μL) was dissolved in 1 mL 0.01% Tween 20 to obtain a working concentration.
The broth microdilution protocol based on the CLSI reference document M27-A3 with modification was used to determine minimal inhibitory concentration (MIC) of yeasts [25].
Briefly, tests were performed in sterile U-bottom 96-well plates. The drug was serially diluted twofold with RPMI 1640 (without sodium bicarbonate and L-glutamine, pH 7.0), and 100 μL of the drug solution at each concentration was used. The concentrations of EO, fluconazole (FCZ), and amphotericin B (AmB) ranged from 40 μL/mL - 0.038 μL/mL and 40 μL/Ml - 0.38 μg/mL, respectively. The inoculum C. neoformans suspension was diluted 10 times with RPMI 1640 broth to obtain a working suspension (approx. 1 × 106 CFU/mL), and 100 μL of the diluted suspension was added to each well. Wells without inocula were used as negative controls, and wells with 0.01% (v/v) Tween 20 but without the EO were used as positive growth controls. The microplates were incubated at 17 ± 2°C for two days. MICs (the lowest concentrations of the test substances that prevented the visible growth of microorganisms) were then determined. To evaluate the minimum fungicidal concentrations (MFCs; the lowest concentrations yielding negative subcultures or only one colony) of the tested drugs, 100 μL solutions from each negative well and from the positive growth control were transferred onto the SDA. The colonies were counted after four days’ incubation at 17 ± 2°C. All experiments were performed in triplicate.
Germ tube formation assay
Germ tube formation was tested following the procedure described previously but with a slight modification [26]. The EO was dissolved in 0.1% Tween 20 to final EO concentrations of 0.391, 0.781, and 1.563 μL/mL, with a final Tween 20 concentration of 0.01%. The controls contained 0.01% Tween 20 without the EO. The final concentrations of AmB and FCZ (positive control) were 3.91 and 50 μg/mL, respectively. The drug solutions were inoculated with spore suspension of each C. neoformans containing 1.0 × 106 CFU/mL and then homogenized by inverting the test tubes three or four times. A total of 100 μL of C. neoformans mixture was placed on separate glass slides and incubated in a moisture chamber at 17 ± 2°C for 8 h. The slides were then observed under the microscope for spore germination. About 200 spores were counted, and the percentage spore germination was calculated.
Determination of biomass
The EO was dissolved in 1 mL 0.01% (v/v) Tween 20 and added to 48 mL of the SDB culture medium to make final concentrations of 0.391, 0.781, and 1.563 μL/mL, and then inoculated with 1 mLC. neoformanscontaining 1.0 × 106 CFU/mL. Control plates (with 0.1% Tween 20 but without the EO) were inoculated with the same procedure. The final concentrations of AmB and FCZ (positive control) were 1.563 and 50 μg/mL, respectively. The flasks were then incubated for 72 h shaking (180 rpm) at 17°C. After incubation, the cells were filtered with sterile Whatman paper and washed twice with sterile distilled water. The cells were dried at 60°C for 6 h and then at 40°C overnight, and weighed. Percent growth inhibition on the basis of dry weight was calculated as follows: (Control weight-Sample weight)/ Control weight × 100 [27]. All experiments were conducted in triplicate.
Determination of ergosterol content in the plasma membrane of C. neoformans
Membrane damage was examined following the procedure described previously but with a slight modification [27]. Briefly, a single C. neoformans colony from an overnight SDA culture plate was inoculated into 60 mL of SDB containing different concentrations of the EO. The EO was dissolved in 0.1% Tween 20, and the final concentrations were 0.391, 0.781 and 1.563 μL/mL. FCZ (50 μg/mL) was used as the positive control. The negative control contained only 0.01% (v/v) Tween 20. The cultures were incubated for 24 h at 17°C in a shaker at 200 rpm. Stationary-phase cells were harvested by centrifugation at 5000 g for 5 min and washed once with sterile distilled water. The wet weights were then determined. About 10 mL of 25% alcoholic potassium hydroxide solution was added to each pellet and vortex mixed for 10 min. Cell suspensions were transferred into clean glass tubes and incubated in an 85°C water bath for 4 h. Sterols were extracted from each sample by adding a mixture of 1 mL sterile distilled water and 3 mL n-heptane, followed by vigorous vortex mixing for 10 min. The n-heptane layer was transferred into a clean tube and stored at -20°C for 24 h. Prior to analysis, 1 mL aliquot of sterol extract was diluted twofold with 100% ethanol, and the spectra between 230 nm and 300 nm were scanned with a spectrophotometer. The presence of ergosterol (282 nm) and the late sterol intermediate 24(28) dehydroergosterol (DHE, 230 and 282 nm) in the n-heptane layer led to a characteristic curve. The ergosterol content was calculated as a percentage of the wet weight of the cells, which was based on the absorbance and wet weight of the initial pellet. The calculation formula of the ergosterol content is as follows:
% Ergosterol + % 24(28)DHE = [(A282/290) × F]/pellet weight,
% 24(28)DHE = [A230/518 × F]/pellet weight,
% Ergosterol = [% ergosterol + % 24(28)DHE]- % 24(28)DHE,
where F is the factor for dilution in ethanol, and 290 and 518 are the E values (in percentage per centimetre) determined for crystalline ergosterol and 24(28) DHE, respectively.
Determination of plasma membrane damage
Membrane damage was examined following the previously described procedure with a slight modification [28]. C. neoformans suspensions were grown in the SDB for 24 h at 17°C in a shaker at 200 rpm. The cell suspension (1 × 106 CFU/mL) of C. neoformans was then added into each glass tube. A certain amount of the EO was added into the tubes to make final concentrations of 0.391, 0.781, and 1.563 μL/mL. Samples with only 0.01% Tween 20 were used as the control, and those with 50 μg/mL FCZ were used as the positive control. The mixtures were then incubated at 17 ± 2°C for 12 h in an incubator shaker. The cells were washed and re-suspended in 500 μL PBS and stained with 1 μg/mL Propidium iodide (PI, Sigma Co.) in PBS for 30 min at 17 ± 2°C. Unstained cell suspensions were always included as auto-fluorescence controls. For each sample, a scattergram analysis was performed to evaluate morphological changes, and the percentage of PI-positive cells was determined using FAScalibur (BD, USA) at FL2 channel (620 nm). All tests were performed in triplicate.
Measurement of mitochondrial membrane potential (mtΔψ).
The effect of the EO on the mt Δψ of C. neoformans was measured by flow cytometry using rhodamine 123 (Rh123) according to a previously described procedure with some modifications [26]. Cells exposed to various doses of the EO (0.391, 0.781, and 1.563 μL/mL) at 17°C for 12 h were harvested by centrifugation at 5000 g for 5 min, washed twice, and then re-suspended at 6 × 106 CFU/mL in PBS. Rh123 was added to a final concentration of 0.1 mg/mL and incubated for 30 min in the dark at 17°C. For all cytofluorimetric experiments, forward and side scatters were gated on the major population of cells, and 6 × 106 CFU/mL cells per sample were measured. mt Δψ was expressed as the fluorescence intensity of Rh123, which was read using a FAScalibur (BD, USA) at FL1 channel (525 nm). The data were analyzed using Cell Quest Pro software. All tests were performed in triplicate.
Measurement of the reactive oxygen species (ROS) formation
The endogenous amounts of ROS in C. neoformans were measured by flow cytometry using 2′,7′-dichlorofluorescein diacetate (DCFHDA) as a ROS indicator as described previously with minor modifications [27]. Exponentially growing C. neoformans cells were suspended in sterile PBS to 6 × 106 CFU/mL. Serial twofold dilutions of the EO (0.391, 0.781, and 1.563 μL/mL) were added to the cell suspensions for 12 h at 17°C. Samples with only 0.01% Tween 20 were used as control, and those with 50 μg/mL FCZ were used as positive control. The cells were harvested by centrifugation at 5000 g for 5 min, washed twice, and re-suspended to 6 × 106 CFU/mL in PBS. DCFHDA was added from a stock solution of 1 mM to a final concentration of 10 mM using a ROS kit (C1300; Apply gen Technology Co.). After 30 min of incubation at 17°C, the cells were collected by centrifugation and re-suspended in PBS to 6 × 106 CFU/mL. ROS was expressed as the fluorescence intensity of DCFH-DA, which was read using a FAScalibur (BD, USA) at FL1 channel (520 nm). The data were analyzed using the Cell Quest Pro software. Additionally, the experiments were performed with the presence of the antioxidant Cys and the EO at concentrations of 60 mM and 0.781 μL/mL, respectively. Control group without Cys and EO. The results were corrected by subtracting the fluorescence value of dill oil ± antioxidant in the corresponding concentration without cells but with DCFH-DA. All tests were performed in triplicate.
Measurement of mitochondrial dehydrogenases
The effect of EO on the activities of mitochondrial dehydrogenases was determined using the XTT method as described previously with some modifications [29]. XTT (Sigma) was prepared as a saturated solution at 1 mg /mL in PBS. The solution was filter sterilized through a 0.22 mm pore size filter, aliquoted, and then stored at -80°C. Prior to each assay, an aliquot of stock XTT was thawed, and 10 mM menadione (Sigma), which was prepared in acetone, was added to a final concentration of 1 mM. C. neoformanswas cultured and diluted to 6 × 106 CFU/mL. The cells were inoculated in 96-well flat-bottomed microplates. Different concentrations of the EO (0.391, 0.781, and 1.563 μL/mL) or the medium alone were added, and the cells were incubated at 17°C for 24 h. A 100 mL aliquot of the XTT/menadione solution was added to each well and to the control wells to measure the background XTT reduction levels. The plates were incubated at 17°C for 4 h in the dark. Fungal mitochondrial dehydrogenase reduced the XTT tetrazolium salt to XTT formazan, thus resulting in a colorimetric change correlated with cell viability. The colorimetric change was measured using a microtiter plate reader (KHB ST-360; Experimental System Co.) at 492 nm.
Preparation for the mitochondria
Briefly, C. neoformans cells (6 × 106 CFU/mL) were used to inoculate 60 mL of the SDB containing various concentrations (0.391, 0.781, and 1.563 μL/mL) of the EO. The cultures were incubated for 12 h shaking (200 rpm) at 17 ± 2°C. The cells were harvested by centrifugation at 5000 g for 5 min, washed twice with sterile distilled water, suspended, and homogenized in a medium (50 mM Tris pH 7.5, 2 mM EDTA, and 1 mM phenyl methyl sulfonyl fluoride). Then, 2% glucose was added to the mixture and centrifuged at 2000 × g for 5 min to remove cell debris and broken cells. The supernatants were collected and centrifuged at 10,000 × g for 30 min, and the pellets were collected as mitochondria. The pellets was resuspended in 50 mM Tris (pH 7.5) - 2 mM EDTA-20 % (v/v) glycerol and stored at -80°C.
Measurement of the succinate dehydrogenase, malate dehydrogenase, and ATPase activities in the mitochondria of C. neoformans
The effects of the EO on the activities of mitochondrial succinate dehydrogenase (SDH) and malate dehydrogenase (MDH) of C. neoformanswere detected using SDH, MDH, and Micro-ATPase Assay Kit obtained from the Institute of Biological Engineering of Nanjing Jianchen (Nanjing, China) following the manufacturer’s protocol. Protein content was determined as described by Bradford, and bovine serum albumin was used as a standard [30].
Statistical analysis
The statistical analysis was performed using statistical software (SPSS, 13.0; Chicago, USA). All data are reported as means ± standard deviations. The significant differences between the mean values were determined by Duncan’s Multiple Range test, following one-way analysis of variance. P value < 0.05 was considered statistically significant.
Chemical composition of the EO
Twenty-two components of the essential oil, accounting for 99.4% of the total oil composition, were identified by GC-MS (Table 1).
| Peak No | Compound | Composition (%) |
|---|---|---|
| 1 | 1R-alpha-Pinene | 11.70 |
| 2 | Beta-Phellandrene | 0.51 |
| 3 | Beta-Myrcene | 1.86 |
| 4 | Alpha-Phellandrene | 1.92 |
| 5 | 1-methyl-2-(1-methylethyl)benzene | 1.09 |
| 6 | Limonene | 52.56 |
| 7 | 3-carene | 1.44 |
| 8 | (4E,6Z)-2,6-Dimethyl-2,4,6-octatriene | 0.59 |
| 9 | Bicyclo[3,1,1]hept-3-en-2-ol,4,6,6-trimethyl | 0.83 |
| 10 | 1,3-Cycloheptadiene | 0.61 |
| 11 | Longifolene-(V4) | 3.67 |
| 12 | Caryophyllene | 1.45 |
| 13 | Tricyclene | 1.22 |
| 14 | Sabinene | 0.61 |
| 15 | Zingiberene | 7.63 |
| 16 | Campholenic aldehyde | 1.79 |
| 17 | Caryophyllene oxide | 0.59 |
| 18 | Apiole | 0.39 |
| 19 | Camphene | 0.78 |
| 20 | 9-Fluorenemethanol | 4.28 |
| 21 | 3- methylcyclohexene | 0.76 |
| 22 | 2-Amino-6-methylbenzothiazole | 1.20 |
| Total | 99.40 |
Table 1: Chemical composition of the EO isolated by hydrodistillation
from C. tinctoria flower.
The most abundant components of the EO were limonene (52.56%), pinene (11.70%), and zingiberene (7.63%). Several other components such as 9-fluorenemethanol (4.28%), longifolene-(V4) (3.67%), phellandrene (1.92%), myrcene (1.86%), 3-carene (1.44%) m and caryophyllene (1.45%) were less in amounts. Oxygenated sesquiterpenes, sesquiterpene hydrocarbons, hydrocarbons, and others were found in trace amounts only.
Anti-C. neoformans effects of the EO
The MIC and MFC values indicated that the EO was active against C. neoformans (Table 2) and exhibited strong antifungal activity. The MIC and MFC levels of the EO against C. neoformans were 0.781 and 1.563 μL/mL, respectively. The MIC levels of AmB and FCZ against the C. neoformanswere 3.906 and 50 μg/mL, respectively. In recent years, the increasing social and health implications of C. neoformans have suggested the constant endeavoring to develop safe and new natural antifungal agents to cure human fungal disorders. In this study, the EO showed a pronounced antifungal efficacy against C. neoformans and a MIC value lower than the positive control (FCZ and AmB).
| MIC | MFC | |
|---|---|---|
| AmB µg/mL | 3.906 | 7.812 |
| FCZ µg/mL | 50 | 100 |
| EO µL/mL | 0.781 | 1.562 |
| Limonene µL/mL | 0.781 | 1.562 |
| Pinene µL/mL | 1.562 | 3.124 |
Table 2: MIC and MFC values of EO, FCZ, and AmB against C. neoformans.
The germ tube formation is a transition state from budding to hyphal cells, and it represents an essential stage to virulence. Therefore, the effect of the EO on the germ tube formation was tested (Figure 1A).
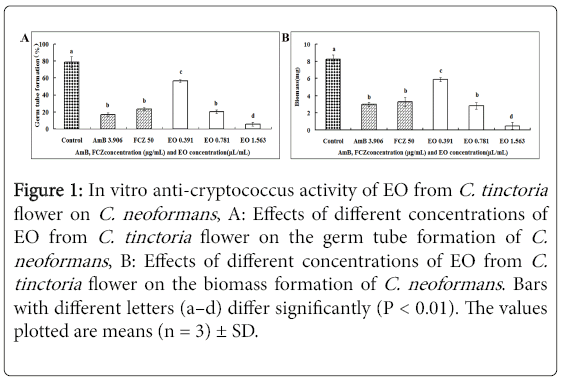
Figure 1: In vitro anti-cryptococcus activity of EO from C. tinctoria flower on C. neoformans , A: Effects of different concentrations of EO from C. tinctoria flower on the germ tube formation of C. neoformans , B: Effects of different concentrations of EO from C. tinctoria flower on the biomass formation of C. neoformans . Bars with different letters (a–d) differ significantly (P < 0.01). The values plotted are means (n = 3) ± SD.
Various concentrations of the EO and positive control showed inhibitory effects on the germ tube formation compared with the controls (P < 0.05). The inhibition calculated on the 8th hour was found to be equal to 56.33%, 20.31%, and 5.38 % for C. neoformans, at the EO final concentrations of 1.563, 0.781, and 0.391 μL/mL. The positive controls FCZ and AmB exhibited inhibition on the germ tube formation at 50 and 3.906 μg/mL, respectively, and the EO showed comparable activities at 0.781 μg/mL and higher activities at 0.391 μg/mL (Figure 1A).
Inhibition of the biomass by the EO was also estimated by measuring the dry cell weight (Figure 1B). The EO significantly inhibited the growth of C. neoformans in a dose-dependent manner (P < 0.05). Similar to the results of MIC, MFC, and germ tube formation experiments, the EO at 0.391 μl/mL showed comparable activity with the positive controls FCZ (50 μg/mL) and AmB (1.563 μg/mL).
Acting mechanism of the EO against C. neoformans
Flow cytometry was used to investigate the membrane integrity of C. neoformanscells after treatment with the EO, with PI as the fluorescent marker. Substantial morphological changes were observed on the scattergrams of C. neoformans cells after 12 h incubation with 0.391, 0.781, and 1.563 μL/mL EO (Figure 2D-2F), and the percentage of PI-stained C. neoformans cells increased along with the concentration.
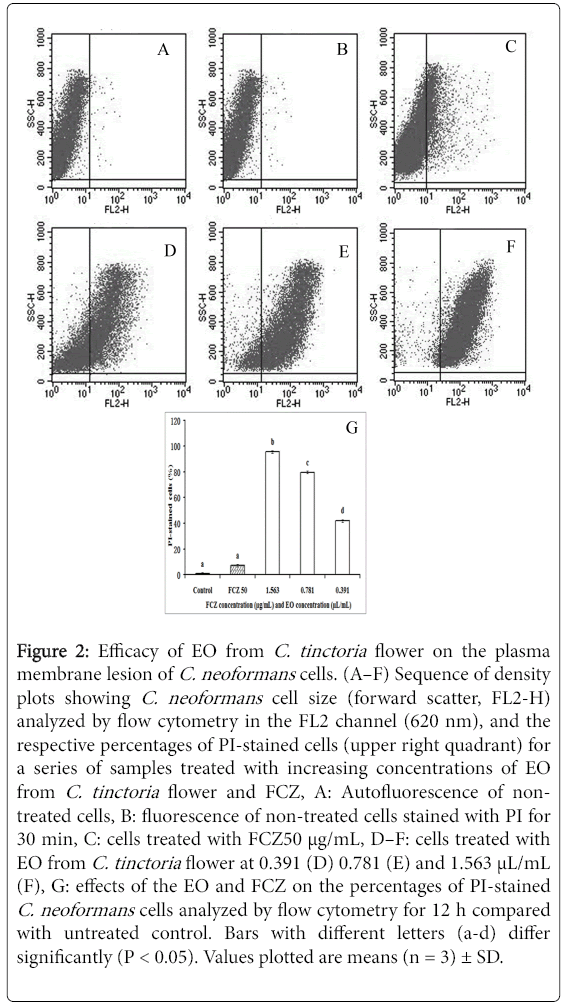
Figure 2: Efficacy of EO from C. tinctoria flower on the plasma membrane lesion of C. neoformans cells. (A–F) Sequence of density plots showing C. neoformans cell size (forward scatter, FL2-H) analyzed by flow cytometry in the FL2 channel (620 nm), and the respective percentages of PI-stained cells (upper right quadrant) for a series of samples treated with increasing concentrations of EO from C. tinctoria flower and FCZ, A: Autofluorescence of nontreated cells, B: fluorescence of non-treated cells stained with PI for 30 min, C: cells treated with FCZ50 μg/mL, D–F: cells treated with EO from C. tinctoria flower at 0.391 (D) 0.781 (E) and 1.563 μL/mL (F), G: effects of the EO and FCZ on the percentages of PI-stained C. neoformans cells analyzed by flow cytometry for 12 h compared with untreated control. Bars with different letters (a-d) differ significantly (P < 0.05). Values plotted are means (n = 3) ± SD.
The percentages of stained cells with 1.563, 0.781, and 0.391 μL EO were 95%, 79%, and 42%, respectively. The results suggested that the cell membrane structure of the C. neoformans was disrupted by the EO in a dose-dependent manner. Under the same experimental conditions, 50 μg/mL FCZ induced only 7.18% of cell death (Figure 2C). These results indicated that the mode of action of the EO involved a primary lesion of the cell membrane, thus leading to cell death. Furthermore, the effect of EO on the cell membrane might be different from that of FCZ.
The total ergosterol content of the plasma membrane of C. neoformans was determined after treatment with different concentrations of EO (0.391 μL/mL-1.563 μL/mL) (Figure 3). The resultant ergosterol contents were 0.037 ± 0.01, 0.023 ± 0.09, 0.017 ± 0.06, and 0.013 ± 0.02, respectively, and the total ergosterol content of the positive control (FCZ at 50 μg/mL) was 0.024 ± 0.03. This finding demonstrated that the ergosterol content of the plasma membrane of C. neoformanswas significantly reduced when treated with different concentrations of the EO in a dose-dependent manner. The decrease in ergosterol content after treatment with 0.781 μL/mL EO (decreased by 46.74%) was comparable with that of the positive control of 50 μg/mL FCZ (decreased by 45.78%), and it was further decreased when treated with 1.563 μL/ml EO (decreased by 73.26%) (Figure 3).
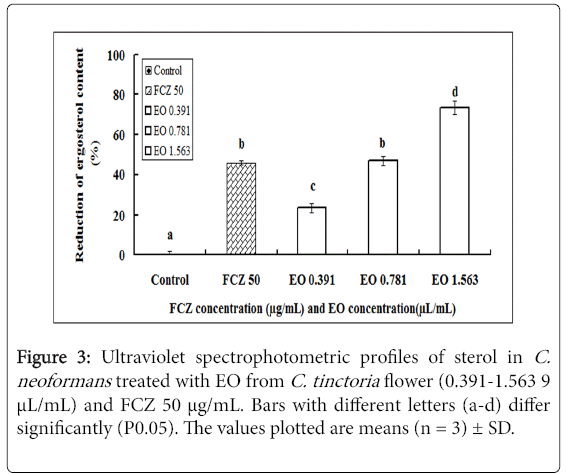
Figure 3: Ultraviolet spectrophotometric profiles of sterol in C. neoformans treated with EO from C. tinctoria flower (0.391-1.563 9 μL/mL) and FCZ 50 μg/mL. Bars with different letters (a-d) differ significantly (P0.05). The values plotted are means (n = 3) ± SD.
The effect of EO on mtΔψ of the cells was first measured by flow cytometry using Rh123, a potential-dependent distributional probe. The results are shown in Figure 4. The mtΔψ values (calculated from the mean fluorescence of all measured cells) of the cells untreated or treated with EO at concentrations of 0.391, 0.781, and 1.563 μL/mL were 84.03 ± 1.24, 183.3 ± 5.13, and 278.76 ± 6.64, respectively (Figure 4C-4E). The mtΔψ value of FCZ (50 μg/mL) (Figure 4B) without the increased intensity of fluorescent cells was similar to the control (Figure 4A). The results showed that the EO from C. tinctoria flower increased the intensity of fluorescent cells in a dose-dependent manner. In addition, EO increased mtΔψ in C. neoformans in a dosedependent manner.
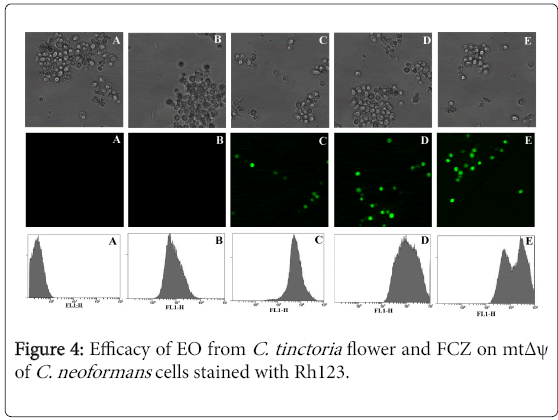
Figure 4: Efficacy of EO from C. tinctoria flower and FCZ on mtΔψ of C. neoformans cells stained with Rh123.
The effect of the EO from C. tinctoria flower on the mitochondria of C. neoformanswas assessed. The activities of the total mitochondrial dehydrogenases (Figure 5A), MDH (Figure 5B), SDH (Figure 5C), and A-TPase (Figure 5D) of C. neoformans were significantly reduced (P < 0.05) when treated with the EO at all concentrations.
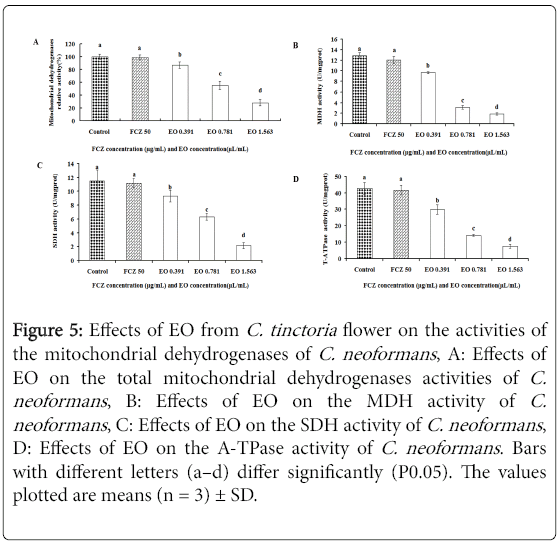
Figure 5: Effects of EO from C. tinctoria flower on the activities of the mitochondrial dehydrogenases of C. neoformans , A: Effects of EO on the total mitochondrial dehydrogenases activities of C. neoformans , B: Effects of EO on the MDH activity of C. neoformans , C: Effects of EO on the SDH activity of C. neoformans , D: Effects of EO on the A-TPase activity of C. neoformans. Bars with different letters (a–d) differ significantly (P0.05). The values plotted are means (n = 3) ± SD.
All of these reductions in the EO activity were dose dependent, and the positive control FCZ (50 μg/mL) without reduction was similar to the control.
The fluorescent dye DCFH-DA, a well-established compound used to assay and quantify ROS production, was used to investigate the changes in the intracellular ROS level of C. neoformans cells.
As shown in Figure 6C-6E, an increase in fluorescence intensity was detected in C. neoformanscells incubated with different concentrations of the EO for 12 h in a dose-dependent manner. Compared with that in the control (Figure 6A) and the positive control FCZ (Figure 6B), the promotion of the fluorescence intensity of the EO corresponded to 7.19, 9.06, and 12.03 times at a concentration of 0.391, 0.781 and 1.563 μL/mL, respectively. The percentage of ROS-positive cells clearly increased with the concentration of EO (Figure 6F). Compared with the control (2.89 ± 1.45 %) and positive control FCZ (3.51 ± 2.03%), the ROS-positive cells of the EO was 82.8 ± 5.45%, 49.20 ± 3.64%, and 24.41 ± 2.87%, respectively. The EO was observed to induce ROS accumulation in C. neoformans cells.
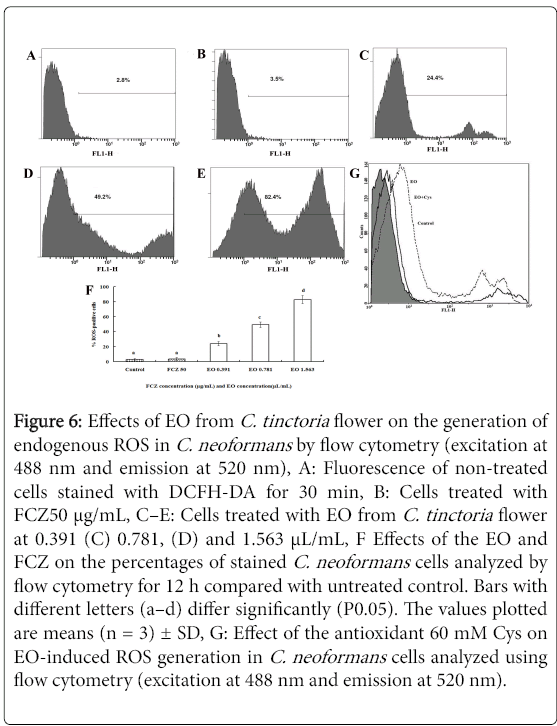
Figure 6: Effects of EO from C. tinctoria flower on the generation of endogenous ROS in C. neoformans by flow cytometry (excitation at 488 nm and emission at 520 nm), A: Fluorescence of non-treated cells stained with DCFH-DA for 30 min, B: Cells treated with FCZ50 μg/mL, C–E: Cells treated with EO from C. tinctoria flower at 0.391 (C) 0.781, (D) and 1.563 μL/mL, F Effects of the EO and FCZ on the percentages of stained C. neoformans cells analyzed by flow cytometry for 12 h compared with untreated control. Bars with different letters (a–d) differ significantly (P0.05). The values plotted are means (n = 3) ± SD, G: Effect of the antioxidant 60 mM Cys on EO-induced ROS generation in C. neoformans cells analyzed using flow cytometry (excitation at 488 nm and emission at 520 nm).
To investigate whether ROS production is directly involved in the antifungal effect of the EO and is not merely a secondary effect of the EO action, the effect of the antioxidant Cys on the net level of ROS production and on the antifungal activity in EO-treated C. neoformans cells was measured. As shown in Figure 6G, the net ROS production in cells induced by EO treatment was evidently reduced by the addition of the antioxidant Cys. With the presence of Cys, the quantity of ROS in cells increased only slightly with the MIC concentration (0.781 μL/ mL). This finding suggests that Cys can arrest the induced ROS production by the EO. Thus, ROS formation plays a critical role in dill oil-induced killing of C. neoformans cells.
EO, which is aromatic volatile products of the secondary metabolism of plants, can be applied in every aspect of daily living [8,31]. EOs has a great antifungal activity due to its synergistic effect with some active components, and thus they are more promising in commercial application than single compounds.
The chemical compositions of the EO from the C. tinctoria flower used in the present study were previously determined by our laboratory. Using GC-MS, the major components detected in the EO from C. tinctoriaflower are limonene (52.56%), pinene (11.70%), and zingiberene (7.63%). Compared with the reported composition of the EO from C. tinctoria flower by other researchers, the limonene and pinene were in agreement but had different contents. Pinene was detected but 9-fluorenemethanol and longifolene-(V4) were not in the EO presented in this paper. A literature search revealed limonene (63.5%), 3-carene (7.0%), p-cymene (5.2%), and zingiberene (2.4%) to be the main constituents of EO from the wild-growing C. tinctoria flower native to the Kunlun Mountain area, Xinjiang, China [19]. Gaspar et al. [12] reported that the major components of C. tinctoria flower EO were α-pinene (45.2%) and trans-carve (28.6 %). These variations in the chemical constituents of the EO may be attributed to the various agro climatic (geographical, climatical, and seasonal) conditions of the regions, the adaptive metabolism of plants, and the stage of maturity.
The anti-cryptococcus activity of the EO from C. tinctoria flower was studied in vitro. In sum, the EO from C. tinctoria flower significantly restricted the germ tube formation and the biomass of the tested C. neoformans in a dose-dependent manner. In vitro studies on EO indicated its potential as an antifungal agent againstC. neoformans. The EO exhibited antifungal activities that could be attributed to the presence of limonene, pinene, and zingiberene [32,33]. To the best of our knowledge, this report is the first on the anti-C. neoformans activity of the EO from C. tinctoria flower.
The plasma membrane plays a vital role in maintaining a homeostatic environment, transferring energy, and exchanging materials and information in the cell to keep cells healthy and alive. PI is one of the most popular fluorescent probes used in assessing the effect of drugs on plasma membranes [34]. Ergosterol is specific to fungi and is the major sterol component of the fungal cell membrane. It is also responsible for maintaining cell function and integrity [35]. The observed dose-dependent antifungal activity of the EO from C. tinctoria flower on C. neoformans cells with a severe lesion of the plasma membrane significantly reduced the ergosterol content. Our results indicated that the plasma membrane is a major target of the EO from C. tinctoria flower, and ergosterol is known to be the major sterol component of the fungal cell membrane and it maintains the cell function and integrity [36]. Other studies have also shown that plantderived EO can cause a considerable reduction in the quantity of ergosterol [34,37], consistent with this study. Thus, the plasma membrane of C. neoformans should be an important antifungal target of the EO from C. tinctoria flower.
Furthermore, the mitochondria play a vital role in the cell, such as in apoptosis, pH homeostasis, redox [38], and ATP production. In the mitochondria, ATP is derived from mitochondrial oxidative phosphorylation, in which mtΔψ plays an essential role [39]. mtΔψ is a sensitive indicator of the energetic state of the mitochondria and the cell [40]. This indicator can also be used to assess the activity of mitochondrial proton pumps, electrogenic transport systems, and mitochondrial permeability transition [41]. The mitochondria of healthy cells maintain mtΔψ across their inner membranes, and this maintenance is the result of an electrochemical gradient maintained through the electron transport chain [42]. Inhibitors of mitochondrial electron transport, such as antimycin A and potassium cyanide, decrease mtΔψ by inhibiting the proton-pumping function of the respiratory chain that leads to decreased ATP production and cell death. A previous study showed that the antifungal action of EO is involved in mtΔψ hyperpolarization in yeast and Aspergillus [43,44]. To the best of our knowledge, the present study is the first to show that the antifungal activity of EO from C. tinctoria flower is causally linked to mtΔψ hyperpolarization in C. neoformans . In the present research, Rh123 was used to examine the effect of EO on mitochondrial potentials. Rh123 is a cationic and lipophilic dye that permeates the negatively charged mitochondria and reflects mtΔψ. Exposure to different concentrations of EO for 12 h showed a significant hyperpolarization of C. neoformans mtΔψ in a dose-dependent manner, thus suggesting mitochondrial dysfunction.
Mitochondria are also the main organelles for ATP synthesis. Mitochondrial dehydrogenases, including MDH and SDH, are important enzymes in the biosynthesis of ATP [45]. MDH catalyzes the interconversion of malate and oxaloacetate in the TCA cycle, and SDH catalyzes the oxidation of succinate to fumarate in the TCA cycle and transfers the electrons from succinate to ubiquinol [46]. In this study, the mitochondrial MDH and SDH activities were measured using assay kits, and the results showed that the mitochondrial MDH and SDH activities of C. neoformans were reduced after the treatment with EO from C. tinctoria flower. This result suggests that the mitochondria of C. neoformans are another important antifungal target of EO from C. tinctoria flower. Moreover, EO could damage the plasma membrane of C. neoformans and affect the enzyme activities of mitochondria; this process could be the antifungal mechanism of EO from C. tinctoria flower.
Mitochondria are organelles inside the cells of higher organisms that represent a major source of ROS. ROS lead to enzyme inactivation, cell dysfunction, and then cell death, and they also play physiological roles in the regulation of apoptosis [47,48]. In the assay for the detection of ROS, an increase in the amount of ROS was apparent after treatment with the EO from C. tinctoria flower. The addition of antioxidant Cys readily prevented ROS production, thus indicating that ROS may be an important mediator in the exhibition of the antifungal effects of the EO from C. tinctoria flower. Recent research has demonstrated that the antifungal action of many antifungal agents is involved in the induction of ROS formation in C. neoformans [49,50]. To the best of our knowledge, the present study is the first to show that the antifungal activity of the EO from C. tinctoria flower is causally linked to an intracellular process in C. neoformans , which is the induction of endogenous ROS. ROS accumulation is a common pathway underlying cellular damage induced by different types of stresses including exposure to EO. Normally, higher mtΔψ is accompanied by a more efficient electron transportation process, which in turn leads to the leakage of more electrons to generate ROS. Previous studies have indicated that ROS generation is exponentially dependent on mtΔψ [51], consistent with the results of our study. That is, both mtΔψ and endogenous ROS were obviously augmented in C. neoformans after treatment with the EO from C. tinctoria flower.
In summary, EO from C. tinctoria flower was successfully extracted and analysed. Its major components are limonene, pinene, and zingiberene. The EO from C. tinctoria flower showed anticryptococcus activity in vitro. It could inhibit the germ tube formation and biomass increase in the C. neoformans . Its mechanism of action was to disrupt the permeability barrier of the plasma membrane and the mitochondrial dysfunction-induced ROS accumulation in C. neoformans . With better antifungal activity than the commonly used antifungal agents and less likelihood of inducing drug resistance, the EO from C. tinctoria flower is a potential natural anti-cryptococcus agent.
The authors wish to acknowledge Rui Zhang for her help with flow cytometry, and Prof. Jianwei Hu for kindly identifying C. tinctoria . This work was supported by the National Natural Science Foundation of China (31301585), the Key Program of Protection and Utilization of Biological Resources of Xinjiang Province of China (BRZD1502).
Conceived and designed the experiments: HZ JT. Performed the experiments: TL HP. Analyzed the data: JT TL. Contributed reagents/ materials/analysis tools: TL HP HZ. Wrote the paper: HZ JT.