Journal of Fertilization: In Vitro - IVF-Worldwide, Reproductive Medicine, Genetics & Stem Cell Biol
Open Access
ISSN: 2375-4508
ISSN: 2375-4508
Research Article - (2015) Volume 3, Issue 2
The superoxide dismutase 1 gene (Sod1) is one of the cell protective mechanisms to eliminate reactive oxygen species damage in order to protect cell components and avoid germinal failure or tumorigenesis. Mouse primordial germinal cells are a population of cells originating from the proximal margin of the epiblast in close proximity to the extra embryonic ectoderm, and from which some cells differentiate into male or female germinal cells. During cell migration across the genital ridge, primordial germinal cells display an extensive proliferation rate and they are exposed to ROS damage. In absence of cell antioxidant mechanisms, primordial germinal cells have increased risk to accumulate specific mutations by free radical damage. We hypothesize that during cell migration, Sod1 knockout primordial germinal cells accumulate microsatellite instability and this could be a signal for primordial germinal cells pool selection, premature ovarian failure or tumor transformation. We dissected and isolated primordial germinal cellsfrom Sod1+/+ (wild type), Sod1+/- (heterozygous), and Sod1-/- (homozygous) mouse strain (B6; 129S7-Sod1tm1Leb/J), from the genital ridge at 10. 5 and 18 days post coitum to determine and quantify microsatellite instability in 10 single tandem repeat markers by single cell PCR methods. Our results demonstrate that primordial germinal cells from Sod1knockout mice accumulated microsatellite instability in five markers (4-4-19844, D13mit16, D13mit78, D19mit68, and DXmit249) located near important genes involved in cell proliferation and differentiation. Taken together, this study shows that microsatellite instability is directly related to Sod1 gene disruption that could induce abnormal migration characteristics seen in primordial germinal cells such as germinal premature failure or tumor invasion and metastasis. In conclusion, microsatellite instability could be a signal for primordial germinal cells pool selection, or tumorigenesis.
<Keywords: Superoxide dismutase-1 gene; Tumorigenesis; Germ cell development; Microsatellite instability
Primordial germinal cells (PGCs) are the progenitors of spermatogonia and oogonia in the testis and ovary, respectively [1]. PGCs originate from the embryonic primary ectoderm (epiblast), and they are one of the cell types that migrate directly through the developing embryo to their site of histogenesis [2,3]. Proliferation and migration of PGCs have been widely studied in different animal models such as mouse, rabbit, sheep, and pig [4]. Of all mammals studied so far, the mouse has provided the most extensive data for the study of early stages of embryonic development [5,6]. There is a similarity in development between mouse and human germinal cells. Therefore, the mouse is an outstanding research model for the study of germ cell lineage [7,8]. Our study utilizes mouse PGCs as a model to evaluate the effects of oxidative stress in germ cell development that leads to genomic instability by accumulation of microsatellite instability (MSI). Elevated free radical damage through reactive oxygen species (ROS) accumulation can override DNA repair mechanisms, giving rise to increased cell mutations that induce infertility, cancer development, chronic diseases (including cardiovascular and neurodegenerative diseases), and aging [9,10].
In mammalian cells, protection against oxidative DNA damage caused by ROS accumulation is accomplished by a complex defense system composed of antioxidant enzymes such as the superoxide dismutase family (Sod1, Sod2, and Sod3) [11]. Sod1 is also known as copper/zinc superoxide dismutase (Cu/Zn Sod). Sod2 is also known as manganese superoxide dismutase (Mn-Sod), and Sod3 is also known as secreted extracellular superoxide dismutase (EC-Sod). All of these Sod genes play critical roles in order to maintain cell oxidative homeostasis and are expressed in the rodent ovary and testis [12]. The Sod1 gene codes for cytosolic superoxide dismutase, which is important for the catalyzation of superoxide ion (O2 -) into hydrogen peroxide (H2O2), and in presence of catalase, H2O2 is reduced to water [13]. The SOD1 enzyme has been isolated from a wide range of organisms such as yeast, bacteria, mice, chicken, and bovine [14]. Sod1 mutant mice display a pleiotropic phenotype that includes neurodegeneration, immunodeficiency, cancer predisposition, and hypersensitivity to ionizing radiation [15]. Although Sod1 knockout mice are born and develop normally, Sod1 deficient female mice are sub-fertile or infertile and display a marked increase in post-implantation embryonic lethality [16]. Altogether, Sod1 deficient mice are prone to infertility and cancer due to an increase of ROS within the ovary affecting its function. Recent observations related to fertility and Sod1 knockout mice are correspondent. Watanabe et al. [17] concludes that Sod1 knockout female mice show repression of plasma progesterone level compared to Sod1+/+ females during pregnancy. Also, Sod1 knockout oocytes could be normally fertilized and developed to the two-cell stage in vitro. However, Sod1 knockout embryos failed to divide to the four-cell stage under conventional culture conditions (20% O2). When Sod1 knockout embryos were cultured under hypoxic conditions (1% O2), they developed to the morula stage but could not develop into blastocysts, indicating that O2 stress inhibited the development of Sod1 knockout embryos at the two-cell stage. Also, Noda et al. [18] concludes that fertilization of the oocytes in vitro conditions was normal, however the development of the embryo was impaired by the deficiency of lutea support that ensure the progress of implantation and pregnancy [17]. This increased ROS to which the Sod1 knockout mice are exposed, results in inhibition of downstream genetic mechanisms that are responsible for the increased risk of ovarian follicular dysfunction and are at a higher risk for premature ovarian failure and cancer development [19].
Endogenous production of ROS from different cellular metabolic processes leads to persistent exposure to oxidative damage [14]. Protective mechanisms against this damage are needed in order to counteract harmful effects in vulnerable cell components such as mitochondria, nucleus, and membrane [9]. However, mutations in the genes responsible for removing superoxide radicals generated in the cytoplasm and nucleus can have a negative impact on genomic DNA integrity, giving rise to defects in cell functionality and cell malignancy transformation [14]. In order to analyze the role of the Sod1 gene in genomic homeostasis of mouse PGC development, we dissected and isolated primordial germinal cells from the genital ridges of Sod1+/+ (wild type), Sod1+/- (heterozygous), and Sod1-/- (homozygous) mice. Our results support the hypothesis that accumulation of genomic DNA instability due to oxidative damage, established by MSI, is a main factor that negatively influences cell development and cell migration. MSI was detected in different mouse single tandem repeats in Sod1 homozygous -/- mouse embryos at 10. 5 days post-coitum (dpc). This study suggest that these markers are important novel target sequences that could be used in humans as a screening method to evaluate the disruption of cell stress responses that lead to initiation and progression of premature ovarian failure or ovarian carcinoma.
Animal model
Uteri of pregnant female mice from the heterozygous superoxide dismutase 1 (Sod1) knockout strain (B6; 129S7-Sod1tm1Leb/J) were purchased from The Jackson Laboratory (Bar Harbor, ME). Details related to the generation of the Sod1 mouse model have been previously described. Briefly, target mutations in the Sod 1 gene were generated by replacement of exon 1 and 2 with a PGK-hptr expression cassette [12]. In the Jackson Lab facilities, male and female heterozygous mice with the superoxide dismutase 1 (Sod1tm1Leb) gene were bred. Females were checked for vaginal plugs and plug-positive females were transferred to necropsy at 10. 5 and 18 days post-coitum (dpc). Uteri were removed from euthanized females and shipped to our laboratory at Mississippi State University. A total of two pregnant uteri at 10. 5 dpc and two uteri at 18 dpc were dissected. From these uteri we obtained a total of 37 mouse fetuses: male (n=18) and female (n=19, with the three expected genotypes: wild type homozygous Sod1+/+(n=11), heterozygous Sod1+/- (n= 20), and homozygous Sod1-/-(n=6).
Isolation and maintenance of primordial germinal cells
From each mouse fetus, the placenta and extra embryonic tissues were removed. Mouse fetuses were transferred into a fresh petri dish filled with D-PBS and the length was measured. Fetus tails were removed for DNA purification, sexing, and genotyping by PCR. Each mouse fetus was dissected following the protocol previously standardized in our lab [20]. Briefly, gonadal ridges were removed by cutting away the tissues that support them. Gonadal ridges were transferred to a petri dish filled with fresh PBS to clean away the surrounding somatic cells. Dispersed suspensions of PGCs from the containing tissues were recovered for isolation of DNA. Characterization of PGCs was achieved using the alkaline phosphatase kit according to the manufacturer’s instructions (Millipore, Billerica, MA).
DNA isolation
DNA was prepared from each fetus tail and from primordial germinal cells harvested from gonadal ridges at 10. 5 and 18 dpc. DNA was isolated with the Purelink genomic DNA mini kit (Invitrogen Carlsbad, CA) following the manufacturer’s protocol. All DNA samples were quantified using a NanoDrop™ ND1000 spectrophotometer (Thermo Scientific, Wilmington, DE).
Sex determination of mouse fetus
Sex determination was achieved by PCR amplifications performed in a total reaction volume of 25μl containing 1X of buffer D (US DNA, Forth Worth, TX), 2. 5mM ofMgCl (US DNA, Forth Worth, TX), 10mM of dNTPs (Applied Biosystems, Foster City, CA) and 1. 25 U/μL of hot multi taq DNA polymerase (5 U/μl; US DNA, Forth Worth, TX). Primers were used as follows: Zfy forward 5’-AAGATAAGCTTACATAATCACATGGA and Zfy reverse 3’-CCTATGAAATCCTTTGCTGCA CATGT, 600bp. The m-DXNds3 forward: GAGTGCCTCATCTATACTTACAG and m-DXNds3 reverse: TCTAGTTCATTGTTGATTAGTTGC, 242 bp [21]. PCR was performed on PE 9600 thermocycler using following cycling protocol: 1 cycle of 95°C for 11 min, 30 cycles of (denaturation for 60 sec at 94°C, annealing for 150 sec at 60°C, and extension for 150 sec at 72°C) final step 1 cycle of 72°C for 2 min and hold at 4°C. Negative controls were included in each PCR. Amplified products were analyzed with gel electrophoresis at 4% nusieve agarose gel (Clonetics, Lonza, Walkersville, MD).
Genotyping of mouse fetus
Genotyping of mouse fetuses to determine presence of the Sod1 transgene was achieved by PCR amplifications performed in a total reaction volume of 25 μl containing 1X of buffer D (US DNA, Forth Worth, TX), 2. 5mM of MgCl (US DNA, Forth Worth, TX), 10 mM of dNTPs (Applied Biosystems, Foster City, CA) and 1. 25 U/μL of hot multi taq DNA polymerase (5 U/μl; US DNA, Forth Worth, TX). Primers used were: Mut781F 5’-TGTTCTCCTCTTCCTCAT CTCC-3’, Mut782R 5’-ACCCTTTCCAAATCCTCAGC-3’, Wt878F 5’TGAACCAGTTGT GTTGTCAGG-3’, and Wt888R 5’-TCCATCACTGGTACAT AGCC-3’ (The Jackson Lab, Bar Harbor, ME). PCR reactions were performed on PE 9600 thermocycler using cycle as follows: 1 cycle for 94°C for 3 min, 35 cycles of (94°C for 20 sec, 60°C for 1 min; 72°C for 1 min); 1 cycle 72°C for 2 min, and hold at 4°C. Negative controls were included in each PCR. Amplified products were analyzed with gel electrophoresis at 4% nusieve agarose gel (Clonetics-Lonza, Walkersville, MD).
Mouse single tandem repeat markers selection and standardization
A total of 21 single tandem repeats (STR) were selected from across the mouse genome. Fifteen markers were located on autosomes, and six markers were located on X chromosome (Table 1). Fluorescent primers were purchased from IDT (Coralville, IA) at 100μM/μL. The labeled dyes used were either 6-FAM (blue) or HEX (green). For initial testing, forward and reverse primers were combined for a final primer concentration of 25μM/μL to create a singleplex primer stock. Primers were tested at concentrations ranging from 0. 8-1. 5 μM/μL in standard PCR conditions and reagents. Each locus was standardized in separate PCR reactions for optimization and to ensure specificity and sensitivity of the marker.
| N. | Marker | Size (Bp) | Repeat Motif | Marker Location |
|---|---|---|---|---|
| 2 | M4-4 | 137 | (CTTTT)17 | Chrom 4 |
| 3 | M4-2 | 129 | (GAAAAA)9 | Chrom 4 |
| 4 | mBat37 | 121 | (A)37 | Chrom 6 |
| 5 | D8mit120 | 130 | (GT)28 | Chrom 8 |
| 6 | D8mit47 | 198 | (TG)14 | Chrom 8 |
| 7 | D8mit88 | 113 | (TG)14 | Chrom 8 |
| 8 | D13mit16 | 209 | (TG)24 | Chrom 13 |
| 9 | 4-4-13-45870 | 390 | (GAGAA)40 | Chrom 13 |
| 10 | D13mit78 | 225 | (CA)25 | Chrom 13 |
| 11 | D14mit5 | 178 | (GT)9 | Chrom 14 |
| 12 | MT2620 | 99 | (CA)13 | Chrom 15 |
| 13 | D17mit20 | 176 | (CATA)13 | Chrom 17 |
| 14 | 4-4-19844 | 280 | (CAGGCT)30 | Chrom 19 |
| 15 | D19mit68 | 132 | (TG)21 | Chrom 19 |
| 16 | DXmit54 | 192 | (CA)15 (GA)16 | Chrom X |
| 17 | DXmit249 | 114 | (TG)17 | Chrom X |
| 18 | DXmit116 | 102 | (CA)19 | Chrom X |
| 19 | DXmit192 | 123 | (CA)24 | Chrom X |
| 20 | mBat-24 | 98 | A(24) | Chrom X |
| 21 | MX-3 | 115-121 | (CA)19 | Chrom X |
Notes: Fifteen markers were located in various autosomes. Six markers were located in the X chromosome. This table depicts the size in base pairs (bp), repeat motifs, and the chromosome location
Table 1: Complete list of 21 microsatellite mouse markers.
Single cell PCR
PCR was performed for 10 markers out of 21 total markers because only these 10 markers amplified during standardization. Two markers came from the mouse X chromosome, one marker each from chromosomes 6 and 17, and two markers each from chromosomes 8, 13, and 19 (Table 2). DNA serial dilutions were made at 0. 1 ng/μL, 0. 05 ng/μL, and 0. 025 ng/μL. Using these serial DNA dilutions, Poisson analysis of the amplified alleles was performed to calculate the correct genomic equivalent value for each sample in order to obtain less than one diploid genome equivalent of the DNA sample (sample genome equivalents range between 12. 5-50 pg/μL) [22,23]. This concentration of DNA allowed sufficient sensitivity to distinguish between wild type and mutated alleles at their appropriate frequency.
| n. | MARKER | CHROMOSOMAL LOCATION (cM) | SIZE (bp) | REPEAT MOTIF | GENBANK NUMBER | PRIMER SEQUENCES | FLUORESCENT LABELS |
|---|---|---|---|---|---|---|---|
| 1 | mBat24 | ChrXq26.1 | 98 | (A)24 | 14734 | CATAGACCCAGTGCTCATCTTCGT | HEX |
| CATCGGTGGAAAGCTCTGA | |||||||
| 2 | D13mit78 | Chr13 (75.0) | 225 | (CA)25 | 62142 | ACAGCACGGGTTTATCATCC | HEX |
| TATGCCTGCCAGGCTTCTAT | |||||||
| 3 | DXmit249 | ChrX(60.5) | 114 | (TG)17 | 493105 | TTATGTGCTTATTAGCCAAGGTG | HEX |
| AAAATAGAACTTCAGCAGCATGC | |||||||
| 4 | D19mit68 | Chr19 (6.0) | 132 | (TG)21 | 16973 | CCAATACAAATCAGACTCAATAGTCG | HEX |
| AGGGTCTCCCCATCTTCCTA | |||||||
| 5 | D8mit47 | Chr8 (56.0) | 197 | (TG)14 | 109384 | AAGATGTGCTTACTCTGACTTCCC | HEX |
| GGATCTATCCACATGTGGTGC | |||||||
| 6 | D8mit88 | Chr8 (61.66) | 113 | (CA)14 | 94457 | GTCCCTTGTAAACACTCTTGCC | HEX |
| CTCTTTGCCCACGGTTATGT | |||||||
| 7 | D17mit20 | Chr17 (34.3) | 176 | (CATA)13 | 12266 | AGAACAGGACACCGGACATC | HEX |
| TCATAAGTAGGCACACCAATGC | |||||||
| 8 | D13mit16 | Chr13 I (10.0) | 207 | (TG)24 | 109406 | CCAGCTGAAGGCTTACTCGT | FAM |
| AAAGTTAGAATCAGCCATTCAAGG | |||||||
| 9 | mBat37 | Chr6qC3 (35.94) | 121 | (A)37 | 1350935 | TCTGCCCAAACGTGCTTAAT | FAM |
| CCTGCCTGGGCTAAAATAGA | |||||||
| 10 | 4-4-19-844 | Chr19 (42.0) | 280 | (CAGGCT)30 | R96357 | CAGTTCACAGGGTAGCCACA | FAM |
| AAGGTACTGCACCTGCTTGG |
Notes: This table shows the specific chromosome location, size of marker (bp), repeat motifs, genbank access number, primer sequences amplified and dye labels for each marker
Table 2: Microsatellite markers for detection of MSI in primordial germinal cells of mouse.
PCR amplifications were performed in a total reaction volume of 10 μl containing 1 X of buffer D [800 Mm Tris HCl, 200 mM (NH4)2SO4, 0. 2% w/v Tween 20](US DNA,Fort Worth, TX), 2. 5 mM MgCl2 (US DNA, Fort Worth, TX), 1 X of Solution L(US DNA,Fort Worth, TX),1. 25 U Hot-MultiTaq DNA polymerase (5 U/μl; US DNA,Fort Worth, TX), 4% DMSO (Sigma Aldrich, Saint Louis, MO), 0. 4 mg/ μLbovine albumin serum (Thermo Scientific, Rockford, IL), 300μM dNTPs (Applied Biosystems, Foster City, CA), and 0. 5 and 1. 8 μM mixed primer (forward and reverse) respectively. Solution L (1X) was used as an additive that facilitates amplification of difficult templates. PCR was performed on a PE 9600 thermocycler using a ramping cycling protocol: 1 cycle of 95 ºC for 11 minutes; 1 cycle of 96 ºC for 1 minute; 10 cycles of [94ºC for 30 seconds, ramp 68 seconds to 58 ºC (hold for 30 seconds), ramp 50 seconds to 70 ºC (hold for 60 seconds)]; 25 cycles of[90 ºC for 30 seconds, ramp 60 seconds to 58 ºC (hold for 30 seconds), ramp 50 seconds to 70 ºC (hold for 60 seconds)]; 1 cycle of 60ºC for 30 minutes for final extension; and hold at 4ºC. Negative controls and reaction mixtures were included in each PCR to monitor for contamination.
Products amplified by single genome equivalent PCR were separated and detected by fragment analysis on an AB 3130XL Genetic Analyzer (Applied Biosystems, Foster City, CA) in the presence of Gene Scan 500 LIZ Ladder (Applied Biosystems, Foster City, CA) following the manufacturer’s protocol. Wild type and mutated alleles were quantified with the Gene Mapper version 4. 0 software package (Applied Biosystems, Foster City, CA). Repeat motif shifts from wild type standardized alleles were considered mutant alleles [22,23]. Presence of wild type and/or mutant alleles was scored for each single cell replicate. An average of 48 replicates per sample were amplified and scored from different samples of mouse PGCs.
Genomic instability statistical analysis
We amplified less than a single diploid genome-equivalent of DNA with single cell PCR methods to estimate oxidative stressinduced mutation frequencies in specific microsatellite repeat markers. DNA concentrations were adjusted to obtain between 0. 5-2 genome equivalents, on average, per single cell PCR reaction. The average number of amplifiable DNA molecules (λ) in each PCR reaction was calculated using the Poisson distribution: λ= -ln (K1/K), where K1 = the total number of alleles expected minus the number of alleles observed, and K =the total number of alleles expected [22,24,25].
Mutation frequencies for each marker by sample genotype, sex, and age were calculated by SP-PCR software version 2. 0 (M. D. Anderson Cancer Center, Houston, TX). Maximum likelihood estimates of the mean number of mutant alleles and wild type alleles in each replicate, as well as their bootstrap standard deviations, were also calculated by SP-PCR software version 2. 0 (M. D. Anderson Cancer Center, Houston, TX). Differences in the estimation of mutation frequencies were calculated with a two tailed t-test using raw mutation frequencies. A three way cross class ANOVA, assuming no third factor interaction, and appropriate LSD multiple comparison (t-test) outputs were also produced. Results were considered statistically significant with a p<0. 05 (using Procedure GLM from statistical package SAS/win 9. 2, SAS Institute Inc. , Cary, NC).
Sod1-/- knockout genotype could be responsible for embryo growth retardation or loss of pregnancy
We obtained two mouse uteri from The Jackson Laboratory at 10. 5 dpc and two uteri at 18dpc. Fetal genotypes were determined with PCR using purified DNA samples of mouse fetus tails dissected during the isolation protocol. We were able to genotype the following 37 fetuses: 19 females including 8 Sod1+/+, 9 Sod1+/-, and 2 Sod1-/-, and 18 male fetuses including 3 Sod1+/+, 11 Sod+/-, and 4 Sod1-/- (Figures 1-3). Drastic reduction in the physical size of female homozygous Sod1-/- mouse fetuses was observed. These results suggest that the reduced number of the female Sod1-/- phenotype might be the result of sub-fertility due to early embryonic reabsorption or spontaneous miscarriage (Figure 3). However, this observation was not statistically significant. The lengths of all dissected fetuses were measured, and we observed that at 10. 5 dpc of gestational age, male and female Sod1+/+ (wild type) and heterozygous Sod1+/- mouse fetuses showed an average size of 14mm. In contrast, female homozygous Sod1-/- mouse fetuses showed a slight decrease in average size at 10 mm, and male homozygous Sod1-/- mouse fetuses showed an average size of 13 mm. At 18dpc, all fetuses showed an average size of 21 mm for all genotypes, and no differences between male and female fetuses were observed (Figure 4).

Figure 1: Determining genotype of mouse fetuses with STS markers: MR0781 and MR0878
Notes: Amplifications from STS markers: MR0781 and MR0878 were analyzed through gel electrophoresis a 4% NuSieveagarose gel.1000bp ladder (EZ BioResearch LLC, Saint Louis, MO) (line1). Sod1 heterozygous (+/-) samples showed both 240 bp and123 bp bands (lines 2 and 3). Sod1 homozygous (-/-) samples showed only 240 bp band (line 4). Sod1 homozygous (-/-) wild type samples showed only 123 bp band (line 5).
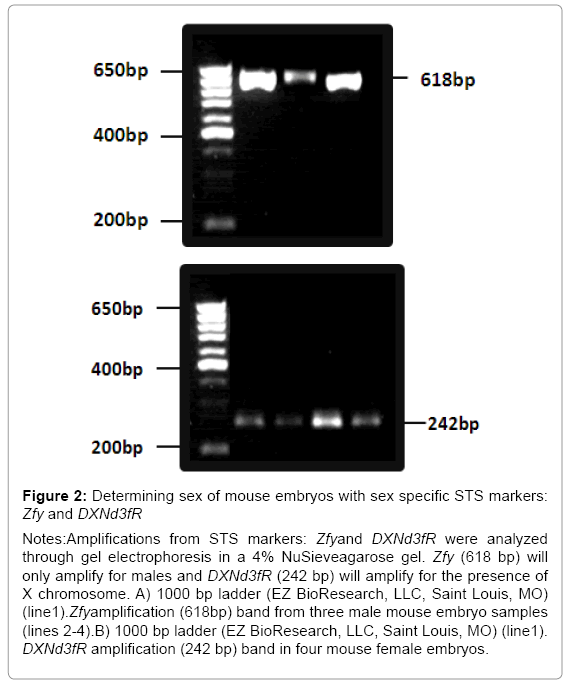
Figure 2:Determining sex of mouse embryos with sex specific STS markers: Zfy and DXNd3fR
Notes:Amplifications from STS markers: Zfyand DXNd3fR were analyzed through gel electrophoresis in a 4% NuSieveagarose gel. Zfy (618 bp) will only amplify for males and DXNd3fR (242 bp) will amplify for the presence of X chromosome. A) 1000 bp ladder (EZ BioResearch, LLC, Saint Louis, MO) (line1).Zfyamplification (618bp) band from three male mouse embryo samples (lines 2-4).B) 1000 bp ladder (EZ BioResearch, LLC, Saint Louis, MO) (line1). DXNd3fR amplification (242 bp) band in four mouse female embryos.
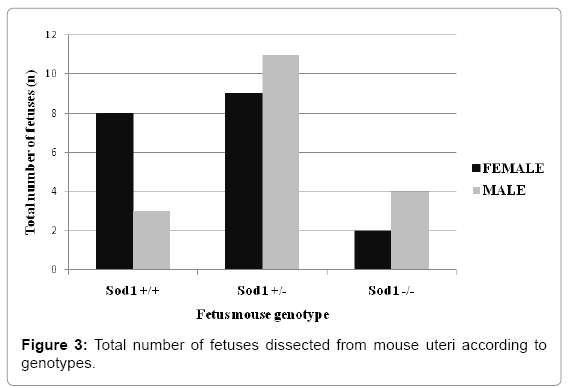
Figure 3: Total number of fetuses dissected from mouse uteri according to genotypes.
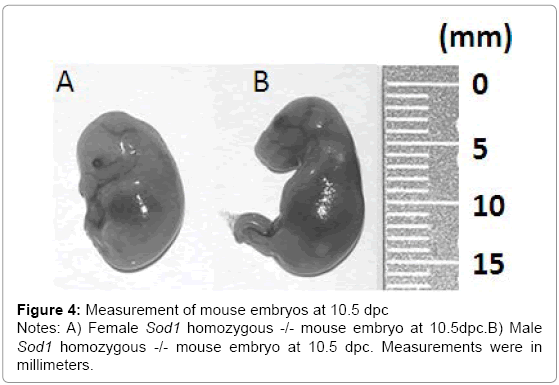
Figure 4: Measurement of mouse embryos at 10.5 dpc
Notes: A) Female Sod1 homozygous -/- mouse embryo at 10.5dpc.B) Male Sod1 homozygous -/- mouse embryo at 10.5 dpc. Measurements were in millimeters.
Genomic instability in single tandem repeats was present in Sod1 knockout mice
Cancer and other disorders such as infertility have been correlated with accumulation of spontaneous mutations due to oxidative stress in different organs such as brain, testes, ovary, and liver [26]. Mechanism of cellular defense against oxidative stress includes Sod1 which is a key molecule in the redox system [14]. We asked whether disruption of the Sod1 function induces accumulation of the spontaneous mutations in PGCs early in embryonic development at 10. 5 dpc and later on at 18 dpc. We determined genomic instability by MSI analysis at two different times during mouse embryonic germinal cell development (10. 5 dpc and 18 dpc). DNA samples from female and male PGCs of Sod1+/+, Sod1+/-, and Sod1-/- genotypes were analyzed. We observed statistically significant high frequencies of mutations in five out of 10 microsatellites evaluated (4-4-19844, D13mit16, D13mit78, D19mit68, and DXmit249) (Figure 5) (Table 3). These results show that spontaneous mutations can accumulate in microsatellite sequences during embryonic development due to a deficiency in mouse PGCs to oxidative stress response.
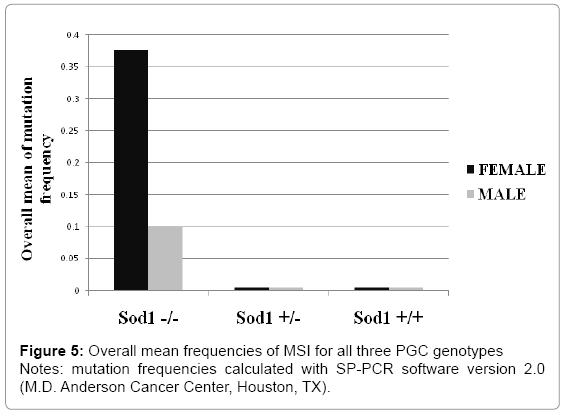
Figure 5: Overall mean frequencies of MSI for all three PGC genotypes
Notes: mutation frequencies calculated with SP-PCR software version 2.0 (M.D. Anderson Cancer Center, Houston, TX).
| Gender | Genotype | Embryo Age (Days post-coitum) | LOCI* | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4-4-19844 | D13mit16 | D13mit78 | DXmit249 | D19 mit68 | |||||||||||||
| n | m | f | n | m | f | n | m | f | n | m | f | n | m | f | |||
| Male | Mutated | 11 dpc | 37 | 0 | 0 | 35 | 2 | 0.031 | 28 | 0 | 0 | 28 | 2 | 0.062 | 33 | 2 | 0.055 |
| 18 dpc | 45 | 0 | 0 | 34 | 0 | 0 | 28 | 0 | 0 | 38 | 0 | 0 | 44 | 0 | 0 | ||
| Heterpzygous | 11 dpc | 14 | 0 | 0 | 44 | 0 | 0 | 36 | 0 | 0 | 46 | 0 | 0 | 47 | 0 | 0 | |
| 18 dpc | 27 | 0 | 0 | 44 | 0 | 0 | 28 | 0 | 0 | 40 | 0 | 0 | 42 | 0 | 0 | ||
| Wild type | 11 dpc | 41 | 0 | 0 | 44 | 0 | 0 | 41 | 0 | 0 | 30 | 0 | 0 | 41 | 0 | 0 | |
| 18 dpc | 46 | 0 | 0 | 91 | 0 | 0 | 42 | 0 | 0 | 43 | 0 | 0 | 44 | 0 | 0 | ||
| Female | Mutated | 11 dpc | 26 | 3 | 0.075 | 26 | 2 | 0.051 | 22 | 5 | 0.15 | 39 | 7 | 0.086 | 41 | 0 | 0 |
| 18 dpc | 36 | 0 | 0 | 42 | 0 | 0 | 39 | 1 | 0.012 | 48 | 0 | 0 | 48 | 0 | 0 | ||
| Heterpzygous | 11 dpc | 45 | 0 | 0 | 46 | 0 | 0 | 39 | 0 | 0 | 47 | 0 | 0 | 48 | 0 | 0 | |
| 18 dpc | 74 | 0 | 0 | 82 | 0 | 0 | 45 | 0 | 0 | 42 | 0 | 0 | 42 | 0 | 0 | ||
| Wild type | 11 dpc | 40 | 0 | 0 | 42 | 0 | 0 | 35 | 0 | 0 | 43 | 0 | 0 | 41 | 0 | 0 | |
| 18 dpc | 30 | 0 | 0 | 46 | 0 | 0 | 44 | 0 | 0 | 40 | 0 | 0 | 40 | 0 | 0 | ||
Notes: *Number of estimated alleles (n), number of mutant alleles (m), and mutation frequency (f) were calculated using SP-PCR software version 2.0 (M.D. Anderson Cancer Center, Houston, TX)
Table 3: Frequencies of MSI at five significantly informative microsatellite markers in male and female mouse primordial germinal cells.
Microsatellite instability occurs early in the mouse PGC development
Mouse PGCs migrate out of the hindgut through the dorsal mesentery at 9. 5 dpc. PGCs arrive at the genital ridge at 10. 5-11. 5 dpc. The complete colonization of the genital ridge by PGCs is achieved at 13 dpc. After PGC migration and colonization, the percentage of PGCs gradually decline from 13. 5 dpc to 16. 5 dpc of mouse embryonic development [27]. We suggest that accumulation of MSI could be the signal on the genital ridge for PGC selection before meiotic arrest takes place. Our results report high mean frequencies of MSI in Sod1 knockout mouse PGC samples, whereas wild type Sod1 and heterozygous Sod1+/- mouse PGC samples did not show MSI over time. Specifically female PGCs Sod1 knockout -/- at 10. 5 dpc showed a high overall mean frequency of MSI in four unstable markers (f= 0. 362) in comparison with the MSI frequency observed just in one unstable marker at 18 dpc (f= 0. 012). Our results suggest that MSI could be a signal for PGC pool selection during early mouse gonad colonization in embryos with abnormal redox mechanisms (Table 3).
Oxidative stress response due to superoxide dismutase 1 deficiency differs between sexes
Previous studies have reported that female Sod1 knockout mice are infertile in comparison with male homozygous mice that are reproductively normal [12,16]. We were interested in determining if MSI accumulation is common in female or male PGCs from Sod1 knockout mouse embryos. Our results showed high MSI frequency in female and male Sod1 knockout mouse PGCs. Female Sod1 knockout PGCs showed statistically high MSI frequencies in four markers at 10. 5 dpc (4-4-19844, D13mit16, D13mit78, and DXmit249) and only one marker showed instability at 18 dpc (D13mit78). In contrast, male Sod1 knockout PGCs showed instability frequencies in three markers at 10. 5 dpc (D13mit16, D19mit68, and DXmit249) and no instability was detected at 18 dpc. Female Sod1 knockout PGCs displayed a significant 3-fold increase in the overall mean mutation frequency compared with male Sod1knockout PGCs (p<0. 05) (Figure 6). Mutation frequencies in wild type and heterozygous Sod1 were stable in both female and male PGCs samples. These observations confirm that MSI could be the cause of female Sod1 reproductive deficiency.
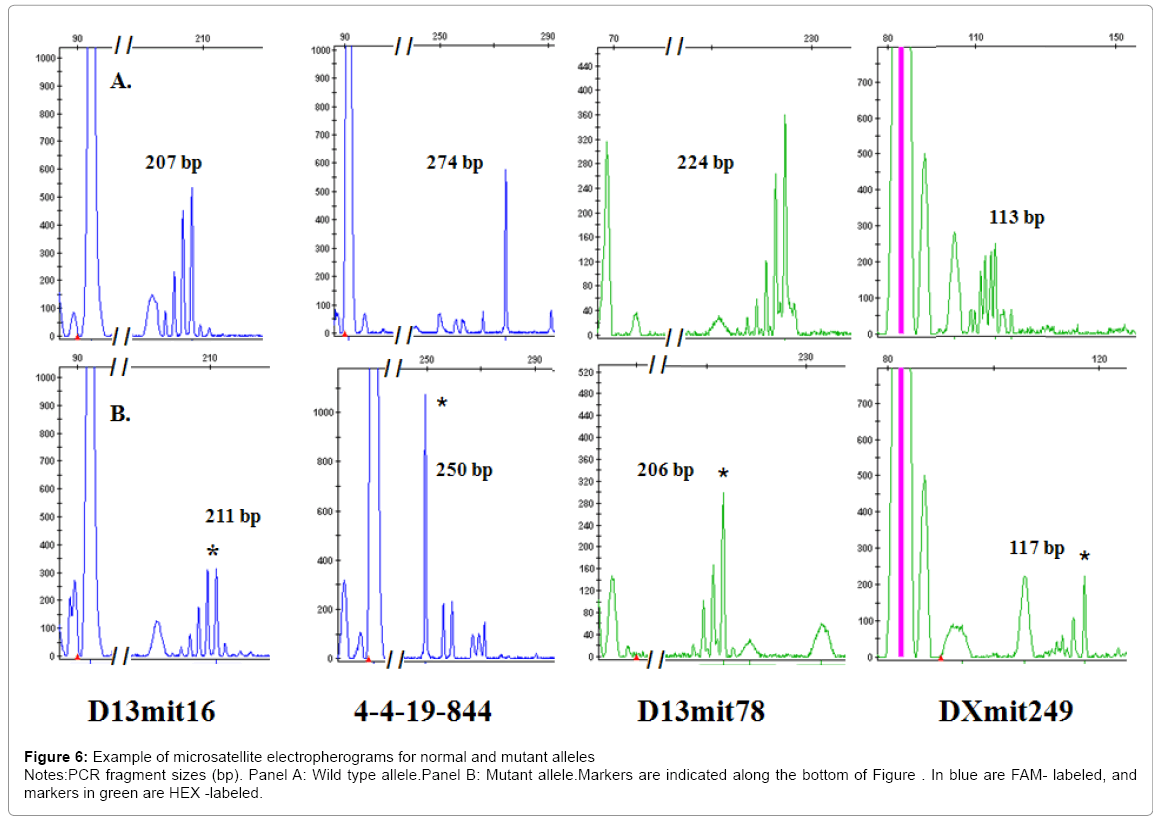
Figure 6: Example of microsatellite electropherograms for normal and mutant alleles
Notes:PCR fragment sizes (bp). Panel A: Wild type allele.Panel B: Mutant allele.Markers are indicated along the bottom of Figure . In blue are FAM- labeled, and markers in green are HEX -labeled.
Microsatellite instability was detected in markers located near genes responsible for cellular stress responses
Upon migration and colonization of genital ridge by PGCs, several molecular pathways are responsible for the establishment of germinal cell pool during the mouse embryonic development [28]. Deregulation of Sod1 gene is a perfect model to evaluate oxidative stress damage as a cause of disease [29,30]. We determined MSI frequencies in two markers in both female and male knockout Sod1 mice. One marker is located near a gene involved in counteracting the oxidative damage mechanism (Dxmit249) and the other marker has been linked to migration and colonization status of PGCs (D13mit16). Significant differences in MSI frequencies were obtained with these two markers at 10. 5 dpc between female and male PGC samples. High MSI frequencies in both markers were observed in female and male Sod1 knockout mice compared to female and male wild type and heterozygous +/- Sod1 mice (p<0. 0001). Dxmit249 marker was highly unstable compared to D13mit16 at 10. 5 dpc in female and male Sod1 knockout mice (Figure 7). At 18 dpc both markers were stable. Our results indicate that MSI detected in markers located near genes responsible for the downstream steps in the antioxidant Sod1 pathway could be the signal for oxidative stress damage.
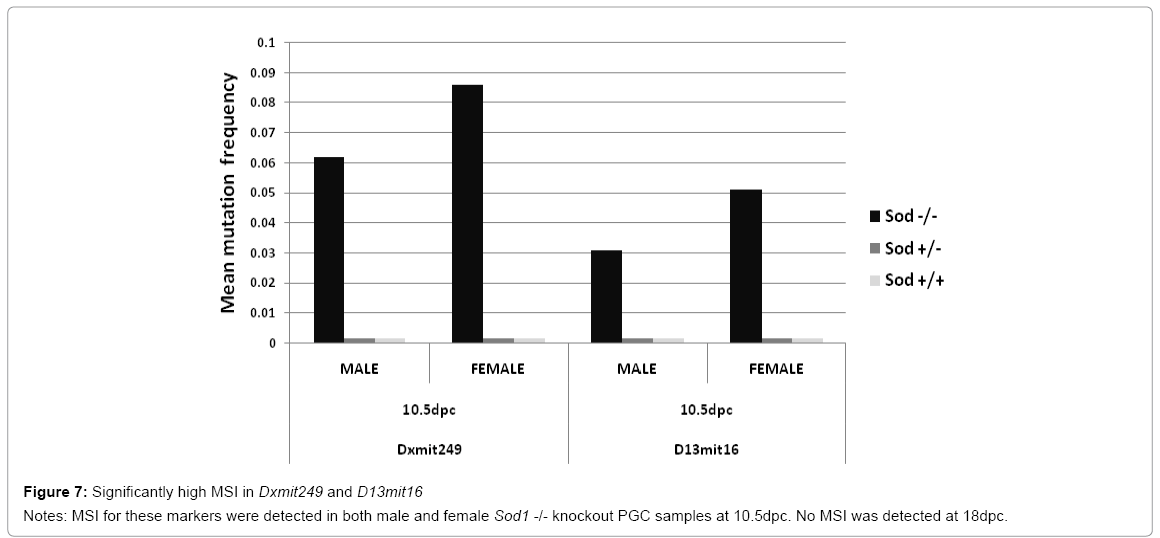
Figure 7: Significantly high MSI in Dxmit249 and D13mit16
Notes: MSI for these markers were detected in both male and female Sod1 -/- knockout PGC samples at 10.5dpc. No MSI was detected at 18dpc.
Embryonic development is disrupted by the presence of MSI in genes responsible for cell lineage commitment
Early during mouse embryonic development, primordial germinal cells are derived from the embryonic primary ectoderm (epiblast) [2]. Developmental genes responsible for cell lineage commitment are targets for the accumulation of spontaneous mutations generated by deregulation of antioxidative pathways [31]. We wanted to demonstrate that MSI is present near genes involved in embryonic differentiation pathways. We found that MSI was present in two unstable markers related to key genes that play a role in the morphogenesis stages of the embryo (D13mit78 and 4-4-19844). Significant differences in MSI frequency were observed in female Sod1 knockout PGCs when compared to male Sod1 knockout PGCs, which were stable for these two markers. The MSI detected in the D13mit78marker was highly significant at 10. 5 dpc in the female Sod1 knockout PGC sample (f=0. 14 p=0. 0001) compared to the MSI frequency of marker 4-4-19844 (f=0. 074 and p=0. 0002) (Figure 8). Unstable alleles were not detected in either wild type Sod1+/+ PGC or heterozygous Sod1+/- PGC samples. Taken together, these findings demonstrate that MSI could be a signal that triggers deregulation of genes involved in germ cell lineage commitment.
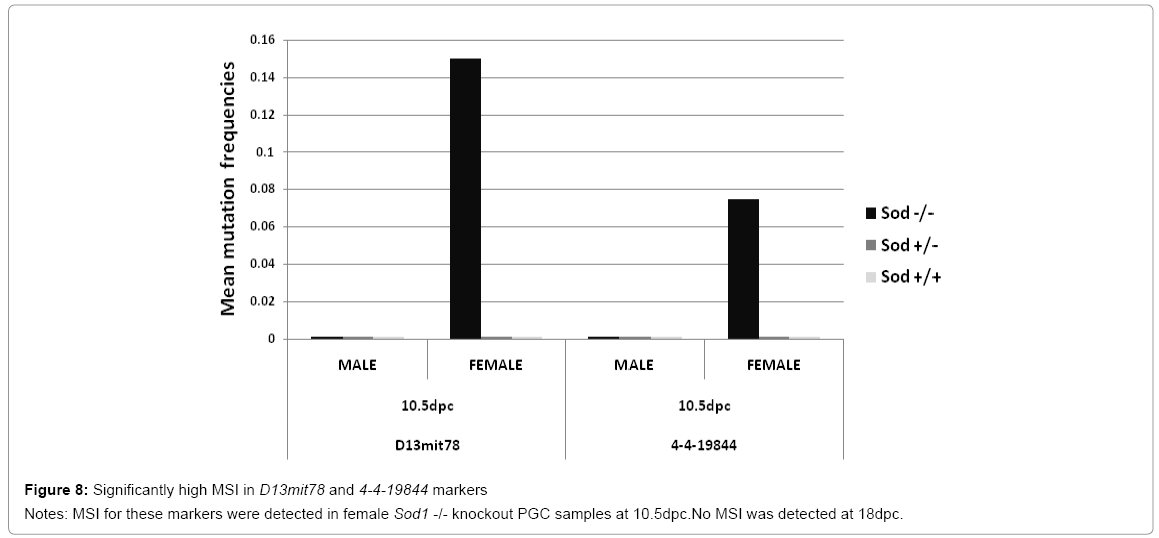
Figure 8: Significantly high MSI in D13mit78 and 4-4-19844 markers
Notes: MSI for these markers were detected in female Sod1 -/- knockout PGC samples at 10.5dpc.No MSI was detected at 18dpc.
Male PGCs are sensitive to microsatellite instability in specific markers related to Wnt cofactor genes
The Wnt signaling pathway participates in embryogenesis. This pathway controls cell proliferation and migration of mouse PGC and is thought to be related to specific functions in gonad and germ cell development [32]. We detected MSI in one marker (D19mit68) that is associated with Wnt cofactor responsible for the intracellular signaling cascade in mouse embryonic development. However, only male Sod1 knockout PGCs were unstable for this marker, and significant MSI frequency was observed in D19mit68 at 10. 5 dpc (p=0. 0067) (Figure 9). This marker was stable in both Sod1 wild type and Sod1 heterozygous male and female mice of all genotypes. Some non-significant instability was detected in PGCs at 18 dpc in both Sod1 wild type and Sod1 heterozygous male and female mice of all genotypes. This result is congruent with the importance of the Wnt signaling cascade function reported by others [33]. MSI detection in this marker could be a signal to deregulate the initializing cofactor for this important developmental pathway due to accumulation of mutations in mouse PGCs.
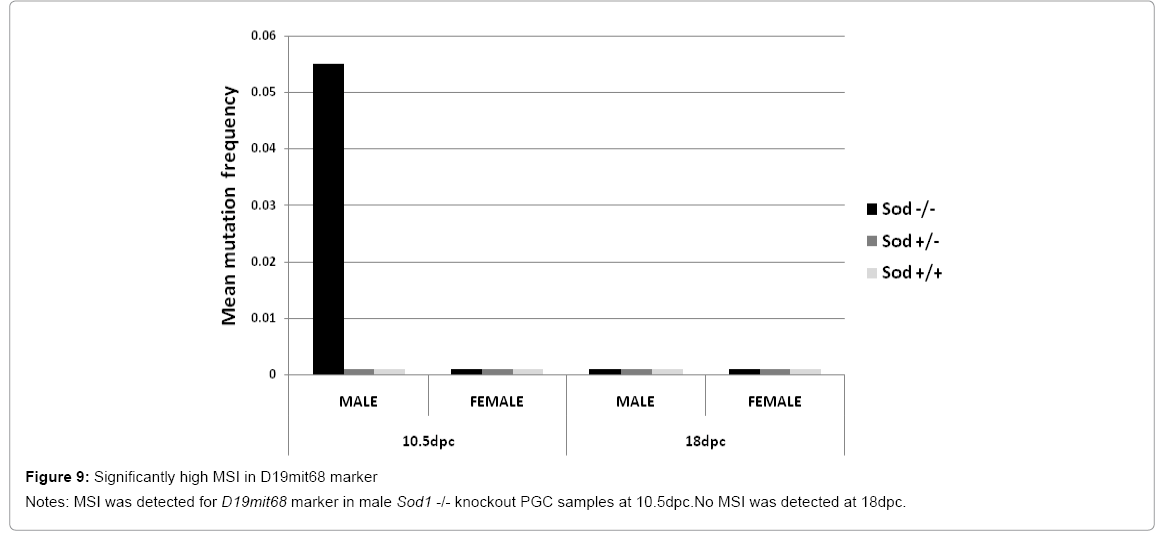
Figure 9: Significantly high MSI in D19mit68 marker
Notes: MSI was detected for D19mit68 marker in male Sod1 -/- knockout PGC samples at 10.5dpc.No MSI was detected at 18dpc.
Characterization of embryonic germinal cells as precursors to male or female gonads in mammals is essential for designing research related to infertility, gametogenesis, and cancer during early development. The numerous possible uses of these cell lines for further basic studies made them an invaluable research tool. Investigations of oxidative stress as a cellular mechanism involved in accumulation of mutations in tumorigenesis, disease development, or cell aging have increased our understanding about different pathways of cell pathogenesis. It is well understood that cells increase the rate of MSI due to accumulation of DNA damage that override DNA repair mechanisms and tumor suppressor pathways, leading to a best-case scenario of apoptosis or a worst-case scenario of tumor transformation. Many mouse models have been developed to study cellular stress response signals [12]. Knockout mice for the Sod1 gene, an antioxidant molecule responsible for the cellular stress response that defends against damage accumulation from oxidative stress, is a notable research model used in our study. Associated with the aging process, Sod1 knockout mice have displayed increases in DNA damage and MSI that are involved in the induction of spontaneous tumors such as in lymphoma, liver, and spleen cancer [12,26,34]. Additionally, these knockout Sod1 mice have developed neurodegenerative disorders such as amyotrophic lateral sclerosis and Parkinson’s disease that have been linked to the oxidative damage [35].
This mouse knockout strain (B6; 129S7-Sod1tm1Leb/J) is an ideal model for our study on the etiology of early ovarian cancer signals. This model has led us to understand and clarify early molecular signals implicated during ovarian development, ovarian tumor transformation, and ovarian cancer progression due to the accumulation of DNA damage by oxidative stress. Genomic instability in PGCs might be the first step of impairment for cell migration and early differentiation in the genital ridges. Our focus is the study of possible signals of genomic instability that induce germinal mutations during the first germ cell commitment in embryonic development. It is known that these signals have been linked to the origin of different cancers such as breast and endometrial carcinomas.
Our results indicate that PGCs accumulate MSI in particular markers located near important genes involved in cell proliferation, differentiation, and migration. This event could be a signal that modulates gonadal colonization by PGCs and selection of the germ cell pool that will be arrested for future gametogenesis in adult mice. We found that PGCs on Sod1 knockout mice have increased spontaneous mutations as determined by detection of high frequencies of MSI in five out of ten markers we tested. Female and male Sod1 knockout mice were significantly unstable in contrast to the complete stability detected for PGCs samples from Sod1 wild type and Sod1+/- heterozygous mice.
The induction of MSI in mouse PGC samples from female and male Sod1 knockout embryonic mice are directly related to the increased sensitivity of spontaneous mutations due to antioxidant defense disruption. Furthermore, our results were in agreement with the observation that Sod1 knockout females are reproductively deficient while Sod1 knockout males are fertile [12]. PGC samples from female Sod1 knockout mice showed instability in four markers located near genes previously linked to neuroectoderm cell lineage commitment and ovarian cancer development. These genes are responsible for normal migration and colonization abilities of the embryonic germinal tissue early during development as well in abnormal invasion and metastasis abilities seen during cell transformation and tumorigenesis.
Female and male Sod1 knockout PGCs were significantly unstable for markers Dxmit249 and D13mit16 in contrast with female and male Sod1 wild type and Sod1+/- heterozygous mice who were stable. Marker Dxmit249 is near the carbonic anhydrase 5b, mitochondrial (Car5b) gene that is involved in the conversion of pyruvate to lactic acid in the presence of oxygen. The Car5b gene catalyzes the reversible hydratation of CO2 to HCO3 in order to maintain tissue pH homeostasis [36]. In cancer cells, high levels of intracellular lactate give rise to over expression of acid-regulating proteins like carbonic anhydrases [37] (Table 4). Our results infer that MSI of marker Dxmit249 could be a signal that deteriorates pH cell defense mechanisms implicated in ovarian cancer disease. Marker D13mit16 is located near engulfment and cell motility protein 1(Elmo1) gene that is responsible for stimulation of normal cell migration. Although its role in different cell processes has not been clearly defined, it is known that Elmo1 protein links to Dock180 protein to form a protein complex (Dock180-Elmo1complex) that plays a critical role in cell adhesion, proliferation, dissemination and invasion of ovarian cancer [38] (Table 4). The Dock180-Elmo1complex activates Rac1, a member of the Rho GTPase family, which participates in the modulation and regulation of the actin cytoskeleton that is essential in cell migration during cancer cell invasion [39]. Our results suggest that MSI in marker D13mit16 could be an early manifestation of the invasive potential that ovarian cancer cells display at the adult stage. However, further studies are needed to conclusively determine the role, if any, that MSI in marker D13mit16 has.
| MARKER | CHROMOSOME LOCATION (cM) | REPEAT MOTIF | MARKER LOCATION | GENE | GENE LOCATION | GENE FUNCTION |
|---|---|---|---|---|---|---|
| 4-4-19844 | Chr19 (42.0) | (CAGGCT)30 | chr19:4288135-4288414 | Adrbk1 | Intron 18-19 | (Adrenergic receptor kinase, beta 1) cell signals involved in inflammation |
| D13mit16 | Chr13 I (10.0) | (TG)24 | chr13:20385311-20385519 | Elmo1 | 5'Upstream 0.2Mb | (Engulfment and cell motility protein 1) protein binding, apoptosis |
| D13mit78 | Chr13 (75.0) | (CA)25 | chr13:119618032-119618260 | Fgf-10 | 5'Upstream 0.1Mb | (Fibroblast growth factor10 precursor); cellprocesses during embryogenesis, adult tissue homeostasis, and carcinogenesis |
| DXmit249 | ChrX(60.5) | (TG)17 | chrX:160413766-160413877 | Car5b | 5'Upstream 0.01Mb | (Carbonic anhydrase 5b, mitochondrialgene);catalysing the interconversion of carbon dioxide and bicarbonate |
| D19mit68 | Chr19 (6.0) | (TG)21 | chr19:3645155-3645286 | Lrp5 | 5'Upstream 0.1Mb | (Low-density lipoprotein receptor-related protein5 precursor) act as a coreceptor of Wnt-pathway |
Notes: This table shows the specific chromosome location, repeat motifs, nucleotide marker location in regards to the chromosome, related genes, marker location of adjacent genes, and gene function information from NCBI, NLM, and NIH databases or from reference papers.
Table 4: Summary list of 5 informative markers and related genes.
Significantly high frequencies of MSI in markers D13mit78 and 4-4-19844 were detected in female Sod1 knockout PGC samples, but both markers were stables in male Sod1 knockout PGCs, as well as in male and female wild type Sod1 and heterozygous Sod1+/- PGCs. Marker D13mit78 is located near the fibroblast growth factor 10 precursor (Fgf-10) gene that has been linked to cell development (Tables 1.3 and 1.4). The Fgf10 gene has been detected in normal human ovarian theca, stroma and endometrial cells that play a role in stimulating the growth of activated follicles [40]. Impairment of Fgf10 gene expression influences paracrine signaling of cell survival, showing its importance for maintenance of follicles and follicular growth [40]. The results of this study denote that MSI on marker D13mit78 could negatively affect early follicular development of female primordial gonads, resulting in increased risk of infertility. Marker 4-4-19844 was unstable and it is located near the adrenergic receptor kinase, beta 1(Adrbk1 also known as Grk2) gene which has a key role in cell signal transduction through the process of receptor desensitization and internalization, avoiding excessive stimulation [41] (Tables 1.3 and 1.4). The follicle stimulating hormone receptor (FSHr) is a clear example of a Grk2transmembrane receptor which is expressed in the ovarian granulosa cells. It has been reported that Grk2 protein translation is affected by the presence of oxidative stress, giving rise to uncontrolled receptor signaling [42]. As a consequence of this, decreased expression of the Grk2 gene has been involved in human granulosa cell tumors probably due to improper receptor desensitization and prolonged activation of FSHr [41]. Taken together, our findings suggest that high risk of female infertility and ovarian cancer could be a manifestation from MSI of markers D13mit78 and 4-4-19844 in PGCs.
Male Sod1 knockout PGCs were significantly unstable in one marker, D19mit68. MSI instability was not detected in male Sod1 knockout PGCs or in female and male wild type Sod1 and heterozygous Sod1+/- PGC samples. The marker D19mit68 is located near low density lipoprotein receptor related protein 5 precursor (Lrp5) gene that acts as a cofactor of the Wnt pathway. The importance of Wnt pathway lies in many cell processes such as cell development, proliferation, fate, and death. It is known that specific mutations of genes that encode this pathway contribute to different types of cancer such as breast, ovarian, and prostate [33]. Our results suggest that MSI on marker D19mit68 could be responsible for malignant cell transformation in male mouse PGCs in the presence of oxidative stress.
In conclusion, our results indicate that accumulation of DNA damage is present in Sod1 knockout mice. PGC samples from Sod1 mice at the time of migration and colonization of the gonad acquired spontaneous mutations due to oxidative stress response deficiencies by Sod1 gene disruption. The presence of MSI during specific times in mouse embryonic development could be a signal for PGC pool selection in the female and male genital ridges and, additionally, to be the on-switch for cell transformation into premature ovarian failure and/or ovarian tumorigenesis when aging deterioration occurs during adult life. Extrapolation of the instability of these markers and genes in the mouse model has also been described in the human genome. Therefore, identification of target mouse genes involved during cell stress responses might shed some light toward determining mechanisms that control oxidative stress consequences in human PGCs. These novel markers might allow for useful screening methods to detect initiation, determination, and progression of premature ovarian failure or human ovarian carcinomas. Supplementary characterization of these genes and their expression status needs to be done in the future to determine the specific functions they have in the evolution of human ovarian failure or tumorigenesis.
Isolation and derivation of mouse embryonic germinal cells (http:// www. jove. com/index/Details. stp?ID=1635)