Advanced Techniques in Biology & Medicine
Open Access
ISSN: 2379-1764
ISSN: 2379-1764
Mini Review - (2015) Volume 3, Issue 3
Genetic toxicology testing has initially concentrated on induction of gene mutations and chromosome aberrations because these genetic changes are considered of great importance for it induces heritable diseases as well as the initiation of cancer. The classical studies on the genetic events underlying the development of tumors proved the importance of phenomenon termed “loss of heterozygosity”. Gene conversion and, mitotic recombination are potent mechanisms leading to loss of heterozigosity. The most important is that the step of induced loss of heterozygosity may be an early or a late step in carcinogenesis. During early embryonic development, groups of cells (imaginal discs) are set apart. They proliferate mitotically during the larval development until they differentiate during metamorphosis into structures of the body of the adult fly (eyes, wings). If a genetic alteration occurs in one of these imaginal disc cells, this alteration will be present in all the descendant cells and will form a clone of mutant cells. The wts (warts) gene was identified based on its ability to act as a tumor suppressor in Drosophila. Deletion, recombination of this gene leads to the formation of cells clones that are rounded and greatly overgrown, and literally generate “warts” (tumors) on their body. This system test will be a useful additional genetic endpoint in toxicological studies in general, as well as in studies to evaluate carcinogenic agents.
Keywords: Drosophila melanogaster, Carcinogens agents, Tumor, Gene wts, Warts
The use of insects, specifically Drosophila, to monitor genetic damage by chemical agents has a tradition of more than 50 years. Drosophila has been used intensively for mutation research and also in short-terms tests for identifying carcinogens [1]. Drosophila melanogaster is a well-established insect model for human diseases and toxicological research due to its well-documented genetics and developmental biology. In this context, there are several advantages that make the fruit fly D. melanogaster a better choice for developing fast and assay systems for detection of chemical with genotoxic activity. The main points are: it is a eukaryotic organism with a short generation time (approx. 10 days at 25°C); it has been easily detectable genetically controlled morphological characters; large number of mutants and genetically characterized strains are available; culture media are inexpensive and allow the breeding of large numbers of animals using simple facilities; it is capable of activating enzymatically promutagens and procarcinogens in vivo.
Although homologous recombination is an important process for DNA repair, there is growing evidence that deleterious genomic rearrangements may result from homologous recombination, which means that homologous recombination events may be determinant in carcinogenesis [2]. The transformation of normal cells into cancer cells is a multistep process. Mitotic recombination is a mechanism involved in bringing about such transformation [3]. In heterozygous cells bearing both mutant and normal alleles for a tumor suppressor gene, the somatic recombination may be a promoter of neoplasms by inducing homozygosis of the mutant tumor suppressor allele [4,5].
During the D. melanogaster embryonic development, the imaginal disc cells proliferate mitotically to form the body of the adult fly. Genetic alterations in some of these cells lead to the formation of mutant cells that are detected as wing-hair mutation spots on the adult fly [6]. In the same way that there are tests used in the mutagenic assessment of chemical, natural and synthetic agents, there are also tests that are applied to assess the carcinogenic potential of these agents, including the test to detect epithelial tumor clones (wts), also performed on D. melanogaster [7,8].
Justice et al. [9] described a new tumor suppressor gene, warts (wts), identified by its clone phenotype in Drosophila. Loss of the wts gene not only results in overproliferation but also in apical hypertrophy of epithelial cells, leading to abnormal deposition of extracellular matrix (cuticle) during adult development. According to Justice et al. [9] the wts gene function is important in controlling cell growth and shape as well as proliferation. For them, the wts gene shows highly significant homology to a gene in Neurospora that is required for the control of cell shape and to the human gene in which mutation causes myotonic dystrophy. Mitotic recombination in somatic cells induces homozygous loss of the warts (wts) gene of D. melanogaster, leads to the formation of cell clones that are fragmented, rounded, and greatly overgrown compared with normal cells [9].
Nishiyama et al. [10] reported the isolation of a putative human protein kinase, termed h-warts, which is highly related to the Drosophila warts protein (serine/threonine kinase domain), which is highly homologous to the human myotonic dystrophy protein, and describes its possible involvement in cell cycle regulation. According to authors, h-warts is phosphorylated in the mitotic phase and hyperphosphoryled when the cell cycle is arrested by disruption of the spindle with antineoplastic agent, which exerts its effect in cells by interfering with the polymerization of microtubules.
The mechanism associated with the formation of tumors is related to Hippo pathway that was identified to orchestrate proliferation and apoptosis to control organ size, initially in Drosophila melanogaster from genetic screens is seeking tissue growth regulators.
Following the discovery of the four tumor suppressors that constitute the core kinase cassette, candidate gene-based approaches and forward genetic screens have implicated at least seven additional tumor suppressors whose activities converge on Hpo and/or wts [11]. Over the past decade, discoveries on Hippo signaling have revealed a complex signaling network integrating various signaling pathways to modulate tissue homeostasis, organ size control, tissue repair, and regeneration. Malfunction of the Hippo pathway is associated with tumor and cancer development [12]. Hippo signalling is regulated by diverse inputs and subject to extensive crosstalk by other pathways, but major factors regulating the pathway includes cellular organization and contacts with neighbouring cells [13]. According to authors Hippo signalling controls organ growth and cell fate by regulating the activity of the kinase Warts. Sidorov et al. [14] examined the impact of genetic manipulations of three Drosophila genes involved in Ado signaling on the incidence of somatic mosaic clones formed by the loss of heterozygosity (LOH) of tumor suppressor and marker genes. Adenosine (Ado) is a ubiquitous metabolite that plays a prominent role as a paracrine homeostatic signal of metabolic imbalance within tissues. It quickly responds to various stress stimuli by adjusting energy metabolism and influencing cell growth and survival. Ado is also released by dead or dying cells and is present at significant concentrations in solid tumors. Ado signaling is mediated by Ado receptors (AdoR) and proteins modulating its concentration, including nucleoside transporters and Ado deaminases [15]. The authors showed that genetic manipulations with the AdoR, equilibrative nucleoside transporter 2 (Ent2), and Ado deaminase growth factor-A (Adgf-A) cause dramatic changes in the frequency of hyperplastic outgrowth clones formed by LOH of the warts (wts) tumor Suppressor. Therefore, Hippo represents an important pathway in Drosophila tumor development.
Some authors (7-8, 14, 17-21), chose the warts (wts) gene for creation of model system, in D. melanogaster, to assessment carcinogenic compounds. Induction of epithelial tumors in D. melanogaster, heterozygous for the suppressor gene wts, can be done using some strains and crosses.
Eeken et al. [7] used a variation of the well-known Drosophila somatic mutation and recombination test (SMART), where mostly recessive markers have been used leading to visible phenotypes in the eyes and wings of the fly. In this case, they used the following strains: 1) wild-type; 2) a strain carrying the X-linked eye color marker white (w), and 3) a strain carrying on the third chromosome wtsMT4-1, a lethal warts allele, balanced over TM3, characterized by multiple inversions and marked by the dominant mutation stubble (Sb). They used three types of crosses: 1) wtsTM3 females with wild-type males (and the reciprocal cross; 2) wtsTM3 females with w males; and 3) wtsTM3 females with In(3)C/M(3)99B males. Wild-type females were mated to wtsTM3 males resulting in F1 females and males with two phenotypes, wts/+ and TM3, Sb wts+/+.
Another hand, Sidorov et al. [14] used different strains and crosses. They used w; P{lacZ w+mC = lacW}wtsP2/TM6B, Hu Tb and wild type Oregon R Drosophila melanogaster stocks. The symbols are: w: white, 1–1.5, recessive, determines the white eye color; wtsP2: wartsP2, 3-100, recessive lethal, tumor suppressor gene; P{lacZ w+mC=lacW}wtsP2: the insertion of the P{lacZ w+mC=lacW} transposon into the wts locus vicinity causing the wtsP2 mutation (this P-element contains lacZ gene from E. coli and w+ allele); TM6B: balancer for chromosome III, homozygous lethal; Hu: humeral, 3-51, additional macrochaetae on the humeri; Tb: Tubby, 3-90.6, larvae, pupae and adults shorter and thicker than wild-type; Hu and Tb are the dominant makers of the TM6B balancer chromosome. To obtain the wts/+ heterozygotes free from balancer chromosome they performed two crosses — P ? w; P{lacZ w+mC=lacW} wtsP2/TM6B, Hu Tb X ? Oregon R (cross B) and reciprocal one (cross R).
However, other authors [8,16-21] used a similar strain and crosses performed by Eeken et al. [7]. They used two mutants D. melanogaster strains for the experiments: [1] wtsTM3, Sb1. This strain presents one lethal wart (wts) on chromosome 3, balanced by the TM3 chromosome, with multiple inversions characterized by the dominant stubble (Sb) mutation, phenotypically identified by short bristles. According to authors, this strain was provided by Bloomington Drosophila Stock Center of the University of Indiana, USA: (Bloomington/7052); [2] multiple wing hairs (mwh/mwh). Flies of the mwh strain have a marker gene on chromosome-3 (3–0.3) in a distal position, with three or more hairs in each cell. The strain is kept in homozygosis since it is a viable mutation. It was provided by Dr. Ulrich Graf (Physiology and Animal Husbandry, Institute of Animal Science, ETH Zurich, Schwerzenbach, Switzerland). The stocks containing these strains are also kept in ¼ L flasks containing culture medium for D. melanogaster at 25°C and 65% relative humidity. To obtain wts +/+ mwh heterozygotic larvae, virgin females wtsTM3, Sb1 [7] are crossed with mwh/mwh males [17]. The eggs of the descendants are collected during an 8 h period. After 72 ± 4 h, third-instar larvae are washed in reverse osmosis water and collected using a fine mesh sieve. The larvae from this cross are treated with the different compounds with carcinogenic potential. The larvae are placed in glass flasks containing 1.5 g of instant mashed potatoes (HIKARI® brand, São Paulo, Brazil) culture medium, and 5 mL of the compound test at the different concentrations. The larvae are submitted to a chronic treatment for approximately 48 h. However, only adult flies, without the chromosome balancer (TM3, Sb1) are analyzed. It is found that they did not have truncated bristles.
Following metamorphosis, the adult flies are transferred to recipients containing 70% ethanol. Later the males and females of the (wts +/+ mwh) genotype, which had wild hairs (long and thin) are analyzed for tumor (wart) presence. The flies are observed using a stereoscopic magnifying glass and entomological tweezers. Only tumors that were large enough to be unequivocally classified are recorded. The tumor frequency is calculated as the number of tumors/number of wts +/+ mwh flies [7].
The carcinogenic potential from compounds is identified by the Mann, Whitney and Wilcoxon nonparametric U test, using α=0.05 level of significance.
As already stated, the test for detection of epithelial tumor clones (warts) in Drosophila melanogaster has been widely used with many different objectives. Here we only present some selected results and discuss important aspects of that assay.
In the treatment procedure of that assay, the larvae are treated chronically in vials containing medium with the test compound at a certain concentration. The larvae feed on this medium until completion of their development when they leave it and putate. Normally, it is not possible to determine the dose of the compound reaching the target cells in the imaginal disc of the larvae. However, methods are available for a quantitative measurement of the chemical compound taken up by individual larvae.
Sidorov et al. [14] evaluated ten chemicals for blastomogenic activity in adult wts/+ heterozygotes of D. melanogaster. To obtain the wts/+ heterozygotes free from balancer chromosome the authors performed two crosses - P ? w; P{lacZ w+mC =lacW} wtsP2/TM6B, Hu Tb X ? Oregon R (cross B) and reciprocal one (cross R). All experimental procedures were carried out at 24 ± 1.5ºC. Parents (10 females and 4 males) were placed into standard cultural vials for 36-40 h for breeding and egg laying, and after 36-40 h transferred onto the fresh medium. The age-heterogeneous population of eggs and first-instar larvae in each vial was treated with 0.3 ml of solutions or suspensions of the substances tested. The controls were treated with distilled water and 10% DMSO water solution.
They have studied 10 compounds of five chemical classes: polycyclic aromatic hydrocarbon (PAH), aflatoxins (AFB1, AFB2 and AFG2), aromatic amides (2-AAF, 4-AAF), heterocyclic aromatic derivatives of N-nitrosamines (NNK) and the complex platinum compound. The blastomogenic and toxic effects of the substances tested are summarized in Tables 1 and 2.
| Substance concentration | Cross-direction | Flies | wts tumor frequency | Survival | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Head | labium | Notum | Wings | Humeri | Thoraicpleura | First leg pair leg pair |
Second leg pair | Third leg pair | Halteres | Abdomen | Total | ||||
| B(a)P su 1.98mM | B | 1673 | 0.54 | 0.00 | 4.36 | 6.22 | 0.00 | 0.84 | 1.44 | 2.21 | 4.30 | 1.14 | 0.00N | 21.04* | 51.64* |
| R | 803 | 0.12 | 0.00 | 2.61 | 5.71 | 0.00 | 0.62 | 0.75 | 0.37 | 0.25 | 0.50 | 0.25 | 11.18* | 58.54* | |
| B(e)P su 1.98 mM | B | RS | 0.14 | 0.00 | 1.56 | 2.13 | 0.00 | 0.28 | 0.43 | 0.57 | 0.99 | 0.43 | 0.00 | 653* | 67.41* |
| R | 468 | 0.21 | 0.00 | 1.28 | 2.35 | 0.00 | 0.21 | 0.21 | 0.00 | 0.64 | 0.00 | 0.00 | 4.92 | 57.42* | |
| Pyrenesu 2.48mM | B | 623 | 0.16 | 0.00 | 1.28 | 1.44 | 0.00 | 0.32 | 0.64 | 0.48 | 0.80 | 0.16 | 0.16 | 5.44 | 53.27* |
| R | 574 | 0.00 | 0.00 | 1.57 | 2.27 | 0.00 | 0.52 | 0.00 | 0.52 | 0.35 | 0.17 | 000 | 5.40* | 55.04* | |
| AFB1su 0.32mkM | B | 1044 | 1.05 | 0.00 | 6.13 | 18.20 | 0.00 | 0.86 | 0.77 | 2.49 | 3.64 | 2.11 | 0.00 | 35.25* | 71.50 |
| R | 742 | 0.40 | 0.00 | 2.16 | 10.78 | 0.00 | 0.67 | 0.27 | 0.67 | 0.81 | 0.40 | 0.00 | 16.17* | 54.68* | |
| AFB2su 0.32 mkM | B | 611 | 0.16 | 0.00 | 2.29 | 1.31 | 0.00 | 0.16 | 0.33 | 0.66 | 2.62 | 0.49 | 000 | 8.02* | 73.68 |
| R | 775 | 0.13 | 0.26 | 2.07 | 2.32 | 0.00 | 0.39 | 0.39 | 0.13 | 1.55 | 0.00 | 0.00 | 7.23* | 67.35 | |
| AFG2 su 0.32 mkM | B | 603 | 0.17 | 0.00 | 2.49 | 3.15 | 0.00 | 0.50 | 0.17 | 0.50 | 3.98 | 0.17 | 0.00 | 11.11* | 83.16* |
| R | 802 | 025 | 0.00 | 1.87 | 2.87 | 0.00 | 0.13 | 0.00 | 0.13 | 0.75 | 0.13 | 0.00 | 6.11* | 68.01* | |
| 2-AAF su 2.24mM | B | 887 | 0.00 | 0.00 | 3.16 | 2.48 | 0.00 | 0.45 | 1.24 | 2.93 | 3.72 | 0.34 | 023 | 14.54* | 55.44* |
| R | 631 | 0.16 | 0.00 | 2.85 | 2.22 | 0.00 | 0.32 | 0.63 | 0.79 | 0.95 | 0.16 | 0.00 | 8.56* | 61.17* | |
| 4-AAF su 2.24mM | B | 967 | 0.10 | 0.00 | 2.69 | 2.48 | 0.00 | 0.21 | 0.21 | 1.14 | 1.66 | 0.00 | 0.00 | 8.48* | 53.76* |
| R | 677 | 0110 | 0.00 | 3.69 | 4.14 | 0.00 | 0.00 | 0.15 | 0.30 | 0.44 | 0.15 | 0.00 | 8.86* | 66.15 | |
| NNK su 0.193M | B | 871 | 0.00 | 0.00 | 1.95 | 1.95 | 0.00 | 0.34 | 0.00 | 0.23 | 2.76 | 0.23 | 0.00 | 7.46* | 62.21 |
| R | 770 | 1.00 | 0.00 | 0.78 | 1.82 | 0.13 | 0.26 | 0.13 | 0.65 | 0.52 | 0.00 | 0.00 | 4.29 | 51.25 | |
| NNK su 0.966M | B | 814 | 0.00 | 0.12 | 1.11 | 3.69 | 0.00 | 0.12 | 0.12 | 2.49 | 0.49 | 0.12 | 0.00 | 5.90* | 63.94 |
| R | 7143 | 0.00 | 0.00 | 0.77 | 2.68 | 0.00 | 0.00 | 0.00 | 0.67 | 0.51 | 0.00 | 0.00 | 3.96 | 57.02 | |
| 10% DMSO (control) (conical | B | 674 | 0.15 | 0.00 | 1.19 | 1.63 | 0.00 | 0.00 | 0.15 | 0.15 | 0.30 | 0.00 | 0.00 | 3.56 | 71.17 |
| R | 1005 | 0.00 | 0.00 | 0.40 | 1.49 | 0.00 | 0.20 | 0.00 | 0.20 | 0.50 | 0.20 | 0.00 | 2.99 | 65.42 | |
*The value is considerably different from the control one (P<0.01)
**The value is considerably differen1 from the Control one (P<0.05)
Data from Sidorov et al. (2001)
Table 1: Total and organ-specific frequencies (%) of wts tumorous clones induced by the organic mutagens of different classes in the wts/+ flies and the survival (%) of the flies of all genotypes up to the adult stagea.
| Solution concentration (mg/ml) | Cross. direction | Flies | wts tumor frequencies | Survival | |||||||||||
| Head | Labium | Notum | Wings | Humeri | Thoracic pleura | First leg pair |
Second leg pail | Third leg pair | Halteres | Abdomen | Total | ||||
| 2.00 | B | 420 | 0.95 | 0.00 | 3.57 | 16.19 | 0.24 | 1.91 | 024 | 024 | 0.48 | 0.24 | 0.00 | 24.05* | 75.71 |
| R | 948 | 1.69 | 0.11 | 2.74 | 8.86 | 0.00 | 1.58 | 0.21 | 011 | 0.11 | 1.16 | 0.00 | 16.67* | 59.98* | |
| 1.00 | B | 1389 | 0.50 | 0.36 | 4.97 | 14.26 | 0.07 | 2.45 | 213 | 2.02 | 2.16 | 1.66 | 0.07 | 30.74* | 7738 |
| R | 445 | 0.23 | 0.00 | 5.62 | 11.01 | 0.00 | 0.90 | 0.45 | 1.35 | 0.90 | 0.00 | 0.23 | 20.67* | 70.69* | |
| 0.20 | B | 575 | 0.00 | 000 | 0.87 | 5.91 | 0.00 | 1.22 | 0.70 | 0.17 | 1.39 | 0.70 | 0.00 | 10.96* | 79.13 |
| R | 646 | 0.46 | 000 | 1.39 | 537 | 0.16 | 0.00 | 0.00 | 0.46 | 0.31 | 0.62 | 0.00 | 8.98* | 62.93* | |
| 0.02 | B | 428 | 0.47 | 020 | 1.17 | 4.44 | 0.00 | 0.23 | 0.00 | 0.23 | 0.47 | 0.23 | 0.00 | 7.24* | 77.01 |
| R | 1492 | 0.00 | 0.00 | 0.87 | 2.15 | 0.00 | 0.00 | 027 | 0.07 | 0.13 | 0.07 | 0.00 | 3.55* | 75.64* | |
| 0.01 | B | 650 | 0.31 | 0.15 | 0.62 | 4.77 | 0.00 | 0.46 | 0.31 | 0.46 | 1.23 | 0.62 | 0.00 | 8.92* | 71.19* |
| R | 1091 | 0.28 | 0.00 | 2.20 | 4.58 | 0.000.00 | 0.83 | 0.18 | 0.18 | 0.37 | 0.18 | 0.00 | 8.00* | 69.67* | |
| Water (control) | B | 1048 | 0.10 | 0.00 | 0.95 | 1327 | 0.00 | 0.10 | 0.19 | 0.10 | 0.19 | 0.38 | 0.09 | 3.53* | 77.42 |
| R | 2598 | 0.08 | 0.00 | 1.00 | 1.58 | 0.00 | 000 | 0.19 | 0.04 | 0.08 | 0.08 | 0.00 | 3.12* | 81.95 | |
aValues in bold pairs: wts clone frequency is considerably higher in the B-crossing compared to the R-crossing (P<0.01)
*The value is considerably different from the control one (P<0.01)
Data from Sidorov et al. (2001)
Table 2: Total and organ specific frequencies (%) of ten tumorous clones induced by the oxoplatin aqueous solutions of various concentrations in the ma+ flies and the survival (%) of the flies of all genotypes up to the adult stagea.
According to the authors, the AFB1 was the most active, inducing two to four times more tumors then it’s less carcinogenic analogues — AFB2 and AFG2. The PAH group the maximal tumor frequency was induced by B(a)P. This frequency was considerably higher than those obtained from its less carcinogenic analogues, B(e)P and pyrene. Tumor frequencies induced by B(e)P and pyrene were closely similar to that in the control and to each other (Table 1).
The authors showed that aromatic amides, 2-AAF displayed the maximal activity. However, its activity strongly depends on the direction of the cross. While in cross B 2-AAF produced tumors 1.5 times more frequently than its non-carcinogenic isomer, 4-AAF, in cross R the activities of both AAFs did not show any differences.
NNK demonstrated relatively low blastomogenity and the tumor frequencies induced by five-fold different concentrations did not significantly differ from each other (Table 1).
The oxoplatin was studied in five concentrations. For this substance an obvious non-linear doze effect was demonstrated. Though the frequencies in cross R were lower than in cross B, their concentration dependency was of the same S-like shape in both reciprocal crosses (Table 2).
Eeken et al. [7] described the effects of the alkylating agent methyl methanesulfonate (MMS) and X-rays on developing flies, heterozygous for a recessive lethal allele of wts, and compare the results with those observed in the SMART using the eye color marker white. In this paper, Wild-type females were mated to wtsTM3 males resulting in F1 females and males with two phenotypes, wts/+ and TM3, Sb wts+/+. Three groups of crosses were started, a control group and two groups that after 2 days of egg-laying received MMS (0.03% and 0.06%). In the reciprocal cross (wtsTM3 females mated to wild-type males) exactly the same results were obtained. The data, from authors, of both series were pooled and are shown in Table 3.
| Total no.of F1 flies | No. of F1 fliesscored | No. of warts scored | Frequency (No. of Warts/fly ± S.D.) |
|||||||
| Eye | Head | Wing | Body | Leg | Halter | Total | Total | |||
| Control | ||||||||||
| wts/+males | 359 | 200 | 2 | 1 | 2 | 2 | 0 | 0 | 7 | 0.04±0.01 |
| wts/+females | 360 | 200 | 0 | 4 | 2 | 2 | 0 | 0 | 8 | 0.04±0.01 |
| TM3/+males | 359 | 200 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| TM3/+ femalesMMS0.03% | 347 | 200 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0.01 |
| wts/+males | 403 | 253 | 116 | 55 | 102 | 56 | 59 | 15 | 403 | 1.59±0.08 |
| wts/+females | 465 | 259 | 148 | 72 | 113 | 101 | 47 | 26 | 507 | 1.96±0.09 |
| TM3/+males | 432 | 258 | 0 | 1 | 0 | 0 | 1 | 0 | 2 | 0.01±0.01 |
| TM3/+ femalesMMS0.06% | 465 | 257 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| wts/+males | 319 | 220 | 136 | 96 | 120 | 109 | 97 | 30 | 588 | 2.67±0.11 |
| wts/+females | 364 | 240 | 245 | 137 | 159 | 148 | 110 | 44 | 833 | 3.47±0.12 |
| TM3/+males | 457 | 235 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0.004 |
| TM3/+females | 463 | 272 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
wts (warts) is located at chromosome-III, TM3 is a chromosome-III balancer preventing meiotic recombination, + defines wild-type chromosomes.
Data from Eiken et al. (2002).
Table 3: Offspring of the Cross Between wts/TM3 and Wild-Type (Summary of Both Reciprocal Crosses), Larval Treatment with MMS.
According to authors, after treatment with MMS, these females showed a higher frequency of clones of white cells in the eye than their brothers. Most of the extra white spots in these females are due to mitotic recombination. The frequency of mitotic recombination is related to the distance of the white gene to the centromere. This distance is approximately similar to the distance of the wts gene to the centromere (on the third chromosome). Both F1 males and F1 females are in addition either wts/+ or TM3/+. The induction of warts clones in these F1 flies is given in Table 4. When they analyzed the offspring of the cross between wtsTM3 females with white males verified at higher magnification for white spots in the eye (Table 4). To demonstrate the validity of the warts system, Eeken et al. [7] they carried out a pilot experiment that was performed using X-rays. Similar to the experiments with MMS, the induction of warts tumors was detected in wts/+ flies.
| Frequency of warts clones per fly ± S.D. | Frequency of white and/or warts spots per fly (i.e., per 2 eyes) ± S.D. |
||||
| No. fliesscored | Total | In eyeonly | No. fliesscored | White/wartsspots | |
| Contro | |||||
| +/Y; wts/+males | 100 | 0.06±0.02 | 0.02±0.01 | 100 | 0.17±0.04 |
| w/+; wts/+females | 100 | 0.09±0.03 | 0.04±0.02 | 100 | 0.28±0.05 |
| +/Y; TM3/+males | 100 | 0 | 0 | 100 | 0.10±0.03 |
| w/+;TM3/+femalesMMS0.03% | 100 | 0 | 0 | 100 | 0.06±0.02 |
| +/Y; wts/+males | 151 | 2.44±0.13 | 0.66±0.07 | 128 | 0.95±0.09 |
| w/+; wts/+females | 155 | 2.66±0.13 | 0.86±0.07 | 147 | 1.52±0.10 |
| +/Y; TM3/+males | 153 | 0 | 0 | 135 | 0.40±0.05 |
| w/+;TM3/+femalesMMS0.06% | 141 | 0.01 | 0.01 | 139 | 0.54±0.06 |
| +/Y; wts/+males | 164 | 3.79±0.15 | 1.13±0.08 | 134 | 1.66±0.11 |
| w/+; wts/+females | 164 | 4.29±0.16 | 1.48±0.10 | 154 | 2.27±0.12 |
| +/Y; TM3/+males | 151 | 0.03±0.01 | 0.01±0.01 | 144 | 0.62±0.07 |
| w/+; TM3/+females | 171 | 0.01 | 0.01 | 166 | 1.07±0.08 |
w (white) is X-linked, Y: Y-chromosome, wts (warts) located at chromosome-III, TM3 is a chromosome-III balancer preventing meiotic recombination, + defines wild-type chromosomes.
Data from Eiken et al. (2002)
Table 4: Offspring of the Cross Between wts/TM3 Females and white Males; Comparison between Induction of Warts Clones with Simultaneously Induced White Spots in the Eyes; Larval Treatment with MMS.
In conclusion, these authors showed that the alkylating agent MMS, as well as X-rays, can induce these tumors in a dose dependent manner. According to them, Warts tumors are detected in the head, the eyes, the legs, and in fact every part of the adult body. For them, this system will be a useful additional genetic endpoint in toxicological studies in general, as well as in studies of the mechanisms leading to LOH, as, for example, DNA repair processes.
Costa et al. [17] used Drosophila melanogaster heterozygous for the tumor suppressor gene wts to evaluate the effect of lapachol on epithelial tumors. Lapachol (4-hydroxy-3-(3-methylbut-2-enyl) naphthalene-1, 2-dione) is a natural quinone that can be isolated from many species of Bignoniaceae found in Brazil, commonly known as “ipê amarelo” [22]. A number of studies have examined this natural quinine, including its anticancer, antiviral, antimicrobial, anti-inflammatory, and antimalarial effects, as well as its significant effect on Trypanosoma cruzi [23,24]. In this experiment, we used the lapachol in combination with doxorubicin, to evaluate the reduction of tumors induced by this chemotherapeutic agent [17].
The following Drosophila melanogaster strains were used: 1) wtsTM3, Sb1 , this strain presents one lethal alleles warts (wts) in chromosome-3, balanced by a TM3 chromosome, having multiple inversions characterized by dominant stubble (Sb) mutation, phenotypically identified by the presence in the entire body of the fly, having short and longer bristles [17]. To obtain wts +/+ mwh heterozygotic larvae, virgin female wtsTM3, Sb1 were mated bred to mwh/mwh males. The larvae from this crossbreeding were treated with the tested chemical compound. However, only adult flies, without the chromosome balancer (TM3, Sb1) were analyzed and they did not have truncated bristles.
All of the compounds were tested in two different experiments, and the results were gathered for the purpose of statistical analyses, after checking that there was no heterogeneity in the samples. A total of 1,600 flies were analyzed, 200 flies from each control (positive or negative) 200 flies from each isolated concentration and 200 flies from each associated concentration. The results demonstrated that descendants treated with different LAP concentration levels (20, 40 or 60 μg/mL), did not show any statistically significant changes in the frequency of tumors when compared to the negative control (5% ethanol) (P>0.05). Therefore, no statistically significant carcinogenic effect of LAP was found in the experimental condition shown here. However, doxorubicin (DXR), used as a positive control, showed an unequivocal enhanced frequency of tumors when compare to negative control (P<0.05) (Figure 1). Evaluating the effect of LAP (20, 40 or 60 μg/mL), associated, simultaneously (co-treatment), with doxorubicin (0.125 mg/mL), a statistically significant reduction (P<0.05) in the frequency of tumors in all concentrations tested was verified (Table 5) [17].
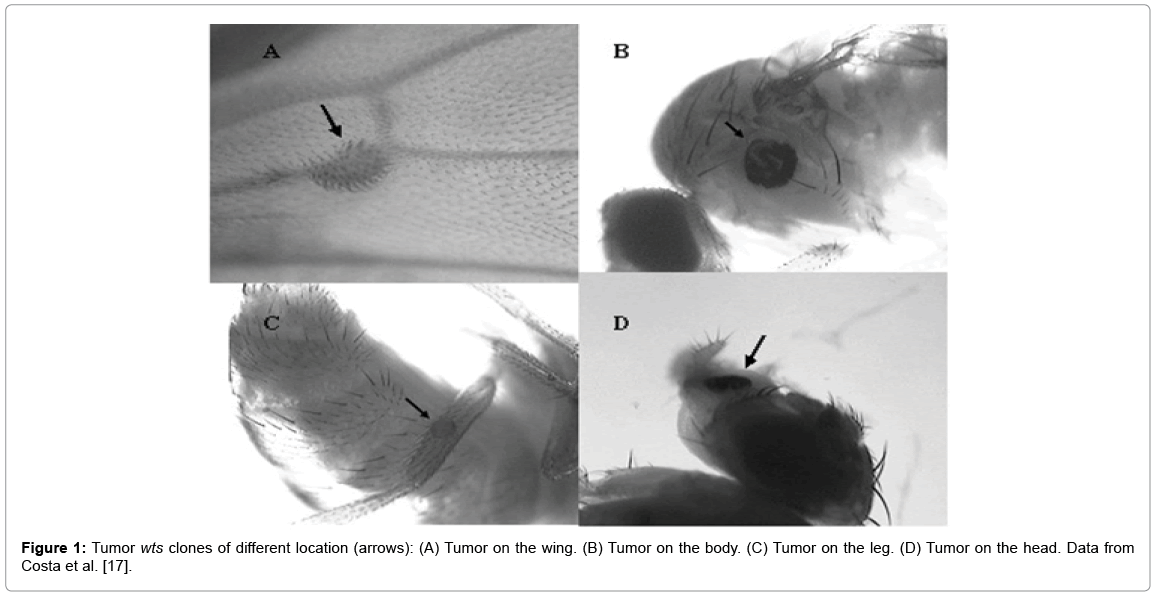
Figure 1: Tumor wts clones of different location (arrows): (A) Tumor on the wing. (B) Tumor on the body. (C) Tumor on the leg. (D) Tumor on the head. Data from Costa et al. [17].
| Treatments | Number of flies scored |
Number of warts scored | Frequency (Nº of warts/fly) | |||||||
| Lapachol (μg/mL) |
DXR (mg/mL) |
Eyes | Head | Wing | Body | Leg | Halter | Total | Total | |
| 0 | 0 | 107 | 0 | 0 | 9 | 5 | 3 | 0 | 17 | 0.16 |
| 0 | 0.125 | 145 | 6 | 12 | 64 | 30 | 20 | 2 | 134 | 0.93* |
| 20 | 0 | 119 | 0 | 7 | 7 | 0 | 0 | 0 | 14 | 0.12 |
| 40 | 0 | 81 | 0 | 0 | 4 | 3 | 2 | 0 | 09 | 0.11 |
| 60 | 0 | 105 | 0 | 0 | 5 | 3 | 2 | 0 | 10 | 0.10 |
| 20 | 0.125 | 93 | 2 | 1 | 11 | 11 | 5 | 1 | 31 | 0.33** |
| 40 | 0.125 | 100 | 0 | 0 | 13 | 11 | 3 | 0 | 27 | 0.27** |
| 60 | 0.125 | 125 | 0 | 2 | 14 | 9 | 0 | 2 | 27 | 0.22** |
Statistical diagnoses according to Mann-Whitney Test. Significance level α=0.05
* The value is considerably different from the negative control (Ethanol 5%) (P<0.05).
** The value is considerably different from the positive control (DXR 0.125 mg/mL) (P<0.05).
DXR, doxorubicin. Data from Costa et al. (2011)
Table 5: Frequency of tumors clones observed in Drosophila melanogaster heterozygous for the tumor suppressor gene wts treated with lapachol and doxorubicin.
Orsolin et al. [8] verified the carcinogenic and anti-carcinogenic potential of orlistat using the epithelial tumor detection test (wts). For verification, larvae from the wts test, descendants of the wts/ TM3 virgin female and mwh/mwh male cross, were treated with the same three orlistat concentrations separately and in association with mitomicin C (0.1 mM). All of the compounds were tested in two different experiments. A total of 1,600 flies were analyzed according to Costa et al. [17]. The results did not, however, provide evidence that orlistat has carcinogenic potential nor was it associated with the reduction of tumors induced by mitomicin C in D. melanogaster (Table 6) [8].
| Treatment | Number of tumors analyzed | ||||||||||
| Orlistat (mg/mL) | MMC (mM) | Number of flies | Eye | Head | Wing | Body | Leg | Halter | Total | Frequency (N° of tumors/fly) | |
| 0 | 0 | 200 | 0 | 10 | 11 | 7 | 17 | 1 | 46 | 0.23 | |
| 0 | 0.1 | 200 | 15 | 89 | 261 | 223 | 121 | 14 | 723 | 3.62* | |
| 2.4 | 0 | 200 | 1 | 14 | 13 | 13 | 14 | 2 | 57 | 0.29 ns* | |
| 4.8 | 0 | 200 | 1 | 6 | 19 | 12 | 19 | 1 | 58 | 0.29 ns* | |
| 9.6 | 0 | 200 | 0 | 6 | 19 | 11 | 14 | 1 | 51 | 0.25 ns* | |
| 2.4 | 0.1 | 200 | 11 | 74 | 270 | 201 | 178 | 12 | 746 | 3.73 ns** | |
| 4.8 | 0.1 | 200 | 19 | 86 | 277 | 185 | 164 | 17 | 748 | 3.74 ns** | |
| 9.6 | 0.1 | 200 | 34 | 93 | 264 | 194 | 153 | 19 | 757 | 3.78 ns** | |
Statistical diagnosis according to the Mann-Whitney Test. Level of significance P ≤ 0.05
* Value consider different from the negative control (P ≤ 0.05).
ns*, values considered not significant when compared to the negative control.
ns**, values considered not significant when compared to the positive control.
MMC, mitomicin C. Data from Orsolin et al. (2012)
Table 6: Tumor clone frequency observed in Drosophila melanogaster, heterozygote for the wts tumor suppressor gene, pre-treated with mitomicin C (six hours) and subsequently treated with different orlistat concentrations.
The “avelós” (Euphorbia tirucalli) is a plant of African origin widely used in popular medicine in the treatment of cancers, ulcers, inflammations and warts. It is considered a toxic plant, because its latex is corrosive in contact with the skin and mucosa. Alves and Nepomuceno [18] used the test for detection of epithelial tumor clones in Drosophila melanogaster, with the objective of knowing the anti-carcinogenic potential of the “avelós”. This way, wts+/+mwh larvae were pre-treated with the chemotherapy drug mitomycin C (0.1 mM), known as a tumor inductor, and afterwards with the aqueous extract of “avelós” (0.33; 0.5 e 1μL/mL). All of the compounds were tested in two different experiments. A total of 1,600 flies were analyzed according to Costa et al. [17]. The “avelós” did not induce increase in the tumor frequency. However, in the association with mitomycin C was verified statistically significant reductions in the tumor induced by this chemotherapy drug. The aqueous extract of “avelós” showed indications of answer dose in association with mitomycin C. Therefore, in the experimental conditions proposed by this study, the latex of “avelós” was able to reduce tumor (Table 7) [18]
| Treatment | Number of flies | Number of tumors analyzed (total tumors) | |||||||
| Avelos (concentration) µL/mL |
MMC (mM) |
Eye | Head | Wing | Body | Leg | Halter | Total | |
| 0 | 0 | 200 | 0.00 (0) | 0.13 (26) | 0.04 (9) | 0.10 (21) | 0.01 (3) | 0.00 (0) | 0.30 (59) |
| 0 | 0.1 | 200 | 0.11 (23)* | 0.54 (108)* | 0.50 (100)* | 0.58 (117)* | 0.17 (35)* | 0.03 (6)* | 1.94 (389)* |
| 0.33 | 0 | 200 | 0.00 (0) | 0.10 (20) | 0.03 (7) | 0.04 (8) | 0.01 (3) | 0.01 (2) | 0.20 (40) |
| 0.50 | 0 | 200 | 0.00 (0) | 0.07 (14) | 0.03 (6) | 0.10 (21) | 0.02 (5) | 0.01 (2) | 0.24 (48) |
| 1.00 | 0 | 200 | 0.00 (0) | 0.06 (13) | 0.02 (5) | 0.05 (10) | 0.00 (0) | 0.00 (0) | 0.14 (28)* |
| 0.33 | 0.1 | 200 | 0.02 (5)** | 0.10 (20)** | 0.31 (62)** | 0.18 (36)** | 0.03 (6)** | 0.01 (3) | 0.66 (132)** |
| 0.50 | 0.1 | 200 | 0.03 (6)** | 0.03 (7)** | 0.31 (62)** | 0.19 (38)** | 0.03 (7)** | 0.01 (2) | 0.61 (122)** |
| 1.00 | 0.1 | 200 | 0.02 (4)** | 0.07 (14)** | 0.25 (50)** | 0.14 (29)** | 0.06 (12) | 0.01 (2) | 0.55 (111)** |
Statistical diagnosis according to the Mann-Whitney Test. Level of significance P ≤ 0.05
* Value consider different from the negative control (P ≤ 0.05).
** Value consider different from positive control (MMC 0.1 mM) (P<0.05).
MMC, mitomycin C. Data from Alves and Nepomuceno (2011).
Table 7: Tumor clone frequency observed in Drosophila melanogaster, heterozygote for the wts tumor suppressor gene, pre-treated with mitomicin C (six hours) and subsequently treated with different avelos aqueous extract.
Rutin is a flavonoid abundantly present in native Dimorphandra mollis of the cerrado, popularly known as field bean or bean-d'anta, and has been the focus of many current studies due to its numerous therapeutic properties. The aim of this study was to examine the anticarcinogenic effect of flavonoid, by testing for the detection of clones of epithelial tumors (warts) in Drosophila melanogaster. A total of 1,600 flies were analyzed according to Costa et al. [17]. The results showed that the larvae which were induced to tumor formation, by MMC (Mitomycin C,) and, subsequently treated with rutin showed a statistically significant reduction in the number of tumors, regarding the positive control. At the concentration of 37.5 mM rutin, there was a reduction of 71% in the overall frequency of tumors induced by MMC; in treatment with a concentration of 75 mM, the reduction was 78%, and the concentration of 150 mM was decreased by 64% in tumors induced in Drosophila [20].
Garlic (Allium sativum L.) since ancient times was used as a condiment and as a medicinal plant, due to its ability to lower cholesterol and blood pressure, to have antiviral activity, antifungal and antibacterial, contribute to blood glucose and strengthen immune system. The test for the detection of epithelial tumor clones in Drosophila melanogaster was used for the purpose of understanding the modulating effect of garlic oil on the carcinogenicity of doxorubicin. For this purpose, larvae wts+/+mwh, were treated with the chemotherapeutic doxorubicin (0.4 mM), known to be tumor-inducing and subsequently with garlic oil (0.5%, 1% and 5%). A total of 1,600 flies were analyzed according to according to Costa et al. [17]. The garlic oil did not induce tumors frequency increase. Moreover, in combination with doxorubicin were verified statistically significant reductions in this chemotherapyinduced tumor. Therefore, in the experimental conditions in this study, garlic oil was capable of reducing tumor [21].
Although many papers using Drosophila as a test organism in the detection of tumors haven not been published, the test for detection of epithelial tumor clones (warts) represent a rapid , very sensitive to different classes of agents, an inexpensive assay to evaluate the carcinogenic activity of single compounds as well as of complex mixtures. For the application of the test materials, various protocols for single or combined as well as for sequential treatments of larvae by feeding are available.