Journal of Pollution Effects & Control
Open Access
ISSN: 2375-4397
ISSN: 2375-4397
Research Article - (2014) Volume 2, Issue 2
When macrophytes are growing in the eutrophicated aquatic ecosystem, the vegetation induces important effects to the water quality and phytoplankton concentrations in the water which affected by macroscopic physical, chemical and biological processes and the effects are the results of direct and indirect interactions of the aquatic plants and water body. The interactions between macrophytes, nutrients and phytoplankton blooms were examined in the water and sediments of a shallow, eutrophic and typical East Taihu, China. The importance of macrophytes as a sink for nutrients, and the inhibitory effect of macrophytes on phytoplankton bloom potential were assessed through three different seasons. Luxuriant aquatic plants growth in this system led to decrease available nutrients for phytoplankton and prevented bloom development. Uptake of N and P by aquatic plants accounted for a major portion of the observed N and P loss from the water column and sediments. Luxury uptake of N and P were indicated by high biomass and tissues N and P concentrations, indicating the capacity of macrophytes to act as a nutrient sink in midsummer. Dissolved inorganic N (DIN) and soluble reactive phosphorous (SRP) in water were reduced in midsummer in the presence of macrophytes. The use of macrophytes to reduce the nutrients in water system and thereby inhibit freshwater phytoplankton blooms should be considered as an effective management strategy in shallow eutrophicated lakes. As aquatic macrophytes also develops considerable indirect effects that could have a vital impact than the direct uptake the nutrients into the plant biomass.
Keywords: Eutrophication; Macrophytes; Nutrients; Algae blooms; East Taihu Lake
Submerged macrophytes are an important biotic component in freshwater ecosystems worldwide [1]. They provide refuge and food for various animals [2] and exert a strong influence on the water physical and chemical properties [3]. Macrophytes alter (generally increase) biodiversity in aquatic habitats [4]. They also provide shelter for zooplankton and young fish, reduce nutrient levels, serve as a habitat for macro-invertebrates [1,4]. Some field studies have demonstrated that the “clearing effect” increased with the macrophytes density and spatial extension of stands, but it was only restricted to a short distance outside the vegetation [5,6].
Freshwater phytoplankton blooms (mainly cyanobacterial) have become an increasingly problematic water quality issue worldwide [7-9]. They represent a health threat to domestic animals and human consumers of affected waters [10,11]. Blooms are primarily caused by excessive loading of nutrients [8] and global warming appears to enhance bloom potentials [12]. In recent years, periodic and widespread phytoplankton blooms have proliferated in the Taihu, China and this phenomenon was particularly serious in Taihu Lake in 2007 [13,14].
Submerged macrophytes can play an important role in the control of phytoplankton within enclosed shallow water bodies [15,16], and a possible negative feedback exists between shading provided by macrophytes and phytoplankton bloom development [17]. Enhanced nitrogen (N) and phosphorus (P) uptake and accumulation by summer biomass buildup of macrophytes leads to N limitation of the phytoplankton [15,18]. We investigated nutrient and macrophyte interactions with respect to their impacts on phytoplankton bloom potentials and water quality in East Lake Taihu, China (referred to as East Taihu). The lake has distinct regions where macrophytes form an important fraction of primary producers. In the relatively clear waters of East Taihu, macrophytes tend to dominate. Conversely, primary production in relatively turbid North Taihu is dominated by phytoplankton. So the purpose of the study was to investigate the role macrophytes played in nutrient cycling in the system by examining the partitioning of N and P between the macrophytes, water and sediment in East Taihu from March to December in 2009.
Study area
The Eastern portion of Taihu is largely comprised of a large (130 km2), shallow (mean depth 1.0 m) Bay, which was called Eastern Taihu. Submerged macrophytes have historically flourished in the lake region [19]. This region is also an important water source for the City of Shanghai [19]. The entire Taihu is currently eutrophic, with total phosphorus (TP) and total nitrogen (TN) concentrations of the water column rarely falling below the OECD threshold value of 0.02 mg/L TP and 0.2 mg/L TN for eutrophic lakes [20].
Three sampling sites (1#, 2#, 3#) in eastern Taihu were selected for transects according to prior results from the Taihu Monitoring Program Stations (Figure 1). The three sites of 1# (E:120.41428°, N:30.98694°), 2# (E:120.4342°, N:31.03294°) and 3# (E:120.49765°, N:31.08131°) were located at the east suburb channel, near commercial crab culture operations, and the main channel of open water region, respectively. Sites were sampled in the March, August and December of 2009. These months represented the germination, maturation and senescence periods for aquatic macrophytes.
Figure 1: The sampling sites in the East Taihu Lake. (The three sites of 1#, 2# and 3# were located at the east suburb channel, near the crab culture and the main channel of open water region, respectively).
Sampling and analytical methods
Six quadrats were located along each transect crossing the studying area. Every sampling site had two quadrats. Water quality data and water samples were collected from all quadrats before collection of submerged macrophytes and sediment samples were undertaken to prevent contamination of the water with sediment. Secchi depth, temperature and dissolved oxygen (DO) were measured on sites. The suspended solids (SS), pH and EC of the water were measured in the laboratory.
Water samples were obtained using a water sampler deployed three times at different depths. Sub-samples were collected for chemical analyses. Surface sediment per quadrat was sampled and then frozen immediately in dry ice. The samples were analyzed for sediment chemical composition on the pooled samples of the top 2 cm of sediments. This is considered to be the portion of sediment where nutrients, P in particular, are enriched [21]. Lastly, the submerged macrophytes were collected from each quadrat, with either a submerged 0.28 m2 quadrat by hand or a circular rake for very small plants [22] or large plants, respectively. The plants were then dried at 60°C for 24 h and weighed.
Water samples were analyzed for total phosphorus (TP), total dissolved phosphorus (TDP), soluble reactive phosphorous (SRP), total nitrogen (TN), total dissolved nitrogen (TDN), ammonium (NH4 +-N), and nitrogen oxides (NOX-N). Concentrations of TDN, NH4-N and NOX-N were determined colorimetrically on a Skalar Autoanalyser (Skalar-SA 3000/5000, Netherlands). TN, TDN were pretreated by digestion with potassium sulphite. TDP and TP were determined after perchloric acid digest, and SRP was determined using the ascorbic acid:molybdate method [23]. Chlorophyll a was measured with hotethanol extraction method as the phytoplankton concentration in the sampling water [24,25]. Sediment TP was measured colorimetrically of Murphy and Riley [26].
Environmental conditions in the East Taihu
Surface water temperature increased from a March mean of 13°C to an August maximum average of 32.0°C (Table 1), and then declined to a December mean of 9.0°C. Bottom temperatures were similar to the surface water in spring, increasing more gradually over the summer time. The trend in surface DO shows a decline from March to August, then a rise to December, with some variability among sampling sites. The water pH and EC varied relatively little, except for some decrease in the aquatic macrophytes maturation period. Water Secchi depth of the 1# site was 0.15 m in March, and at the same time it were 0.90 m and 0.95 m at 2# site and 3# site, respectively. But the Secchi depth of 1# site reached 1.60 m in August, it was higher than that the other two sites and the water Secchi depth changed along with the season and macrophytes growth.
| Sampling site | 1# | 2# | 3# | ||||||
| Sampling time | Mar | Aug | Dec | Mar | Aug | Dec | Mar | Aug | Dec |
| Secchi depth (m) | 0.15 | 1.60 | 1.00 | 0.95 | 1.22 | 0.90 | 0.45 | 0.85 | 1.30 |
| Water depth(m) | 1.00 | 1.60 | 1.50 | 0.95 | 1.22 | 0.90 | 1.00 | 1.60 | 1.40 |
| Temperature(m) | 13.0 | 32.0 | 9.0 | 13.0 | 32.0 | 9.0 | 13.0 | 32.0 | 9.0 |
| pH | 7.92 | 8.33 | 8.02 | 8.72 | 7.80 | 7.79 | 8.09 | 7.85 | 7.87 |
| EC(μs/cm) | 560.0 | 520.0 | 460.0 | 420 | 500.0 | 520.0 | 510.0 | 420.0 | 500.0 |
| DO(mg/l) | 10.10 | 9.85 | 4.50 | 7.56 | 4.87 | 9.70 | 8.30 | 7.65 | 9.60 |
| SS(mg/l) | 118.64 | 2.16 | 7.77 | 0.80 | 2.61 | 9.30 | 20.88 | 6.56 | 8.93 |
Table 1: The water environmental characteristic in East Lake Taihu in 2008
(Integrated samples of the water column were obtained using a water sampler deployed three times at different depths. Then water samples were brought back to the laboratory with the ice-bag protected and analyzed as soon as possible. The each sampling time was chosen at the same 9:00 am ; and all values presented are mean values).
Nutrients changes in water and sediment
The selected three sampling sites exhibited similar water depth profiles and fluctuations (Figure 2). However, the three sites showed some variability among different forms of nutrients in the water. They had the higher concentration in March at 1# site, and then dropped to the minimum in August, but then increased again in December. The other sites showed an inverse pattern. Dissolved nitrogen and phosphorus concentrations in water column were sufficiently high to be available to macrophytes and phytoplankton. Sites 2# and 3# had the lower concentration of the TDN and TDP, and then increased from March to August. During the period from August to December, only the concentration of TDP in 1# sampling site rose (Figure 3). The PO4 3--P and NO3 --N in water appeared to be directly available to macrophytes and phytoplankton. Though the NO3 --N was higher in March, it decreased quickly in summer.
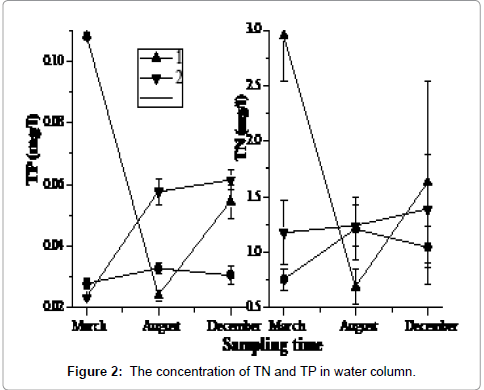
Figure 2: The concentration of TN and TP in water column.
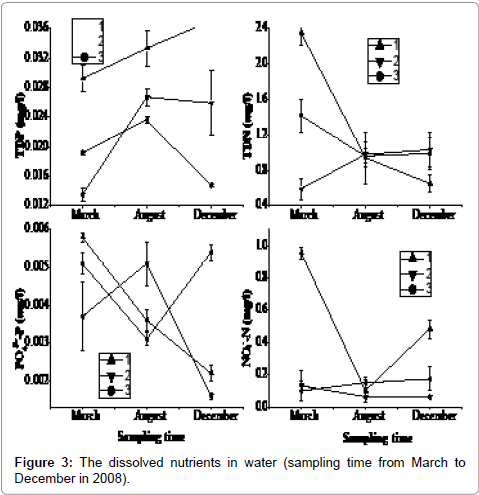
Figure 3: The dissolved nutrients in water (sampling time from March to December in 2008).
The TN and TP concentration in the sediments were similar to those in the water column. However, the 2# site had the higher concentration in March, and it was the time of the germination period of aquatic macrophytes, after which the nutrient concentrations declined during the summer (Figure 4). Nutrient accumulations appeared largely attributable to macrophytes decomposition.
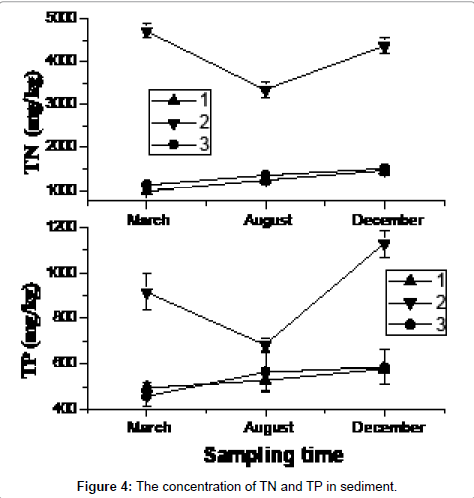
Figure 4: The concentration of TN and TP in sediment.
Macrophytes and phytoplankton in the water column
The three sampling sites showed similar environmental conditions in the water and the macrophytes occurred in shallow regions to a maximum depth of 1.60 m in August and a maximum depth of 0.95 m in March. Macrophytes community composition also varied with sampling times and sites. The main species of the macrophytes included: Potamogeton malaianus Miq.; Vallisneria natans L.; Elodea nuttallii; Hydrilla verticillata Royle; Ceratophyllum demersum with other species comprising only a small percentage of the total composition (data not shown).
Site 1# is located at the east suburb channel, the flood discharge channel of Taihu and main water supply for the City of Shanghai. The macrophytes could obtain amounts of nutrients and grow well as the water velocity became lower at the 1# site (Figure 5). The 2# site had the highest biomass of aquatic plants during the selected sampling sites in March. But the 1# site had the highest biomass among three sampling sites in summer (fresh weight reaching 2,255.48 g/m2). The 3# site had low biomass relatively except in August. Though the species composition and the macrophyte biomass showed little difference during the sampling time, in general there was an increase in aquatic plant biomass at sampling sites except some decreases at the 3# sampling site.
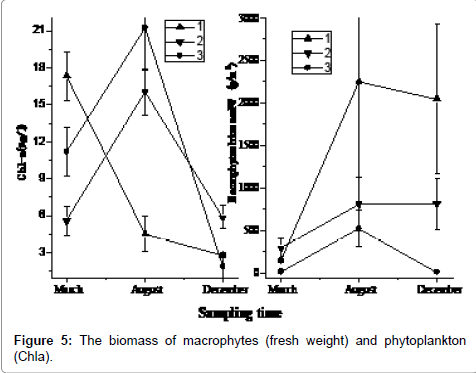
Figure 5: The biomass of macrophytes (fresh weight) and phytoplankton (Chla).
Phytoplankton biomass (dominated by cyanobacterial), estimated as Chl-a concentrations, varied among sampling times and sites (Figure 5). The concentration of phytoplankton biomass at 1# site was higher than that at the others sampling sites in March, but it was the lowest among the three sampling sites in August, only 5.58 μg Chla L-1. The phytoplankton biomass generally increased in the summer, except site 3# showed a decrease. The biomass in 2# site became progressively higher over summer, reaching a peak in December (21.2 μg Chla L-1).
Conditions within the water column
The increase in water temperature in early summer at the germination period of aquatic macrophytes would favor growth of these plants. The early summer increase in biomass of photosynthetic plants would explain the increase DO concentrations in water in March. Differential photosynthetic use of carbon dioxide would also explain the fluctuation in pH at the three sampling sites. Higher water temperatures could also cause a decrease in the oxygen solubility in the water and therefore the August DO concentrations were reduced at all sampling sites (Table 1). As some macrophytes decomposed during this senescent period, DO consumption would also increase. As mentioned above, the macrophytes grew most vigorously from March to October, as temperature favored the aquatic plants and phytoplankton growth in this period. The results of Wang [27] indicate that water temperature and total phosphorus (TP) played dominant roles in controlling phytoplankton growth dynamics in most seasons; COD (chemical oxygen demand) and BOD (biological oxygen demand) presented significant positive relationships with phytoplankton biomass in spring, summer and autumn.
Macrophytes and nutrients
N and P values in the Taihu sediment are significantly different from those found in East Taihu sediments [28-30]. TN and TP concentration in the water was higher in 2# site in March and then decreased through the summer period. As in the water column, the TN in sediment also had higher content in December than that in August, although TP varied little during this period. The N and P concentration changes in the sediments indicate that nutrient losses from the top 2 cm of sediment were similar to the nutrient gains of the macrophyte community (Figure 5), of which the major species was the submerged macrophyte Potamogeton malaianus. The N data showed an increase in macrophyte N which was not matched by loss from the sediment, a phenomenon that has been observed by others [31,32]. TDN decreases in water column, although considerable (Figure 3). The additional N may have been derived from deeper sediments or from western Taihu water, although P data do not support this (Figure 3).
The macrophytes biomass progressively increased from March to August (Figure 5), a period when temperature and irradiance are favorable for macrophyte growth. Macrophyte species diversity was also enhanced during the same period (data not shown). Thriving aquatic plants during this period will optimally adsorb dissolved nutrients from the water, and increase the depth of the photic zone (Table 1). In a word, these effects would promote a positive-feedback, environment friendly ecosystem [33].
The Inhibition Effects of Macrophytes on Phytoplankton
The use of macrophytes for reducing N and P availability and thereby reducing phytoplankton blooms potential has been used in diverse aquatic ecosystems [18,34,35]. Possible inhibition of phytoplankton by allelochemicals released by submerged macrophytes has been proposed as one of the mechanisms that contribute to the maintenance of clear-water states in shallow lakes [36,37]. Although direct proof of allelopathy remains elusive, several authors have suggested possible involvement of allelopathy to explain phytoplankton successional patterns in whole-lake studies of vegetated, shallow lakes [38,39]. During this survey, the Chl-a concentration was low at the sampling sites except for 1# in March, when it was somewhat higher (Figure 5). The reason have been that this sample was taken from a site that was influenced by open Taihu water which passed through the 1# site, which was the main flood discharge channel of the Taihu. Following the summer macrophyte growth period, phytoplankton concentrations rapidly decreased. The macrophytes decayed at higher temperature in summer and caused oxygen deficiency in water column at the 2# site, as proven the nutrients concentration variety (e.g. higher concentration of PO4 3--P, TDP in water). The apparent inhibitory effects of macrophytes on phytoplankton are consistent with the other results [40-42]. Another explanation is salinity (conductivity) control of nutrient uptake by macrophytes [43], as the conductivity changed during surveying time (ranged from 560 to 42 μs/cm during the March to August time). Decreased salinity would promote uptake of N and P by these macrophytes, resulting in decreased availability of nutrients for phytoplankton.
Aquatic macrophytes have been shown to be a significant sink for nutrients and they increased clarity in East Taihu. Higher summer macrophyte biomass was responsible for uptaking large amounts of N and P. Macrophytes incorporation of N and P accounted for most of the observed nutrient loss from the sediment and water column. Surficial sediment N loss did not account for all the macrophyte N gain. Therefore other N sources, including cyanobacterial N2 fixation and atmospheric N deposition, are postulated. Water column TDN and NO3 --N significantly decreased during the P. crispus growth period. Decreased salinity (conductivity) maybe have also promoted N and P incorporation by macrophytes and hence the reduction of available nutrients for phytoplankton, explaining a reduction in phytoplankton biomass and blooms during the summer. The inhibitory effect of macrophytes on phytoplankton blooms has significant ramifications for management of phytoplankton blooms.
Thanks to the anonymous reviewer for his hard work in reviewing the article and he put forward many constructive suggestions and pointed out much minor mistakes in manuscript, his work is important for the authors to improve the paper quality.
This research was supported financially by the National Natural Science Foundation (NO. 41101525) and Independent Innovation Capital of Jiangsu province (NO. CX(12)5057).
No competing financial interests exist.