Immunome Research
Open Access
ISSN: 1745-7580
ISSN: 1745-7580
Research Article - (2017) Volume 13, Issue 3
The aim of this study was to evaluate the modulatory effect of zymosan on human langerhans cells (LCs). The expression of Toll-like receptors (TLRs) and cytokines in LCs treated with zymosan was evaluated by determining the mRNA levels of TLR-1, TLR-2, TLR-4, TLR-9 and the mRNA levels of IL-1β, IL-6, IL-8, IL-10, IL-12, IL-23, TNF-α, and the protein level of bioactive IL-12p70. Furthermore, correlation coefficient of the mRNA levels of TLRs and cytokines was also evaluated to determine their mutual effects. The examined 10 samples showed significant increase in the mRNA level of those cytokines in all. As for TLRs, significant increase in TLR-2, decrease in TLR-1 and TLR-4 though the changing extent were slight and no significant difference in TLR-9 was shown. No significant increase of bioactive IL-12p70 protein was shown. All cytokines except for IL-12 were positively correlated to each other (r>0.3) at 24 h after zymosan treatment and to TLR-2 at both 8 h and 24 h. As our conclusion, zymosan may modulate LCs by increasing transcriptional level of cytokines and TLR-2 expression. Inflammatory and non-inflammatory cytokines in zymosan-treated LCs changed expression in a balanced style, while IL-12 was increased in all samples regardless of other cytokines and TLRs levels. All the tested cytokines except IL-12 and TLR-2 have a mutual promotion effect.
<Keywords: Cytokine; Langerhans cell; Toll-like receptor; zymosan
APCs: Antigen-Presenting Cells; CLRs: C-type Lectin Receptors; DCs: Dendritic Cells; GAPDH: Glycerldehyde 3-Phosphate Dehydrogenase; LCs: Langerhans Cells; LPS: Lipopolysaccharide; PAMPs: Pathogen-Associated Molecular Patterns; PRRs: Pattern Recognition Receptors; PBMCs: Peripheral Blood Mononuclear Cells; qRT-PCR: Quantitative Real-Time PCR; TLRs: Toll-Like Receptors
Langerhans cells (LCs) are unique subsets of dendritic cells (DCs) which are located in epidermis and mucosa, the very first immune barrier of body [1]. Because of the special distribution, LCs as predominate Langerhans cells (LCs) are unique subsets of dendritic cells (DCs) which are located in epidermis and mucosa, the very first immune barrier of body [1]. Because of the special distribution, LCs as predominate mucocutaneous antigen presenting cells (APCs) plays a crucial role in local immune responses. Except for antigen presenting, LCs secretes cytokines that affect whole immune system function to improve both innate and adaptive immune responses. DCs express pattern recognition receptors (PRRs) that recognize pathogen-associated molecular patterns (PAMPs) on invading products such as microorganisms. Among the PRRS, Toll-like receptors (TLRs) and C-type lectin receptors (CLRs) play important roles in DC-based immunity [2]. Therefore, it is predicable that LCs may recognize and bind to antigens through TLRs or CLRs and trigger some downstream cytokines production.
Fungal cell wall is most composed of polysaccharides, for instance, polymers of mannose, glucose (β-glucan) and N-acetylglucosamine, which have been demonstrated to possess immunostimulatory properties [3]. Zymosan is a kind of glucide compound extracted from cell wall of heat-killed yeast Saccharomyces cerevisiae, which is composed of β-(1,3) glucan, mannose and other substance [4]. Its immune activity has been extensively investigated by many researchers [5,6]. Jiang et al. reported zymosan can modulate inflammatory responses by increasing intracellular cAMP and activating protein kinase A pathway in macrophage cells from mice [5]. Human PBMCs incubated with zymosan were demonstrated to secrete relatively higher level of IL-1β, IL-6, IL-23 and TNF-α by Syk- and Src-dependent pathways [6]. Among various kinds of human immune cells, we have our study restricted to relationship between zymosan and LCs in this current study, which comes to a more precise conclusion whether zymosan can improve human epidermal or mucosal immunity. Zymosan is prepared from fungi cells and its component is similar with major PAMPs of some fungi, such as Candida albicans (C. albicans) [7,8]. Therefore, the investigation of the effect of zymosan on LCs may give hints for the effect of some fungi with similar PAMPs existing on the surface of skin or mucosa on local immunity.
In our previous study, it has been demonstrated that zymosan can induce higher expression level of CLRs including Dectin-2, Mincle and MRC-2 of LCs [9]. In this current study, we aimed to further investigate immune regulation effects of zymosan by determining mRNA transcriptional levels of TLRs including TLR-1, TLR-2, TLR-4, TLR-9 and a variety of cytokines including IL-1β, IL-6, IL-8, IL-10, IL-12, IL-23, IFN-γ and TNF-α in LCs. Also we examined protein expression level of IL-12p70 of LCs stimulated with zymosan. The correlations among TLRs, cytokines and between TLRs with cytokines were evaluated to study their mutual effects. To our knowledge, this is the first time to demonstrate the modulatory effect of zymosan on human LCs. The mutual effects of all the tested cytokines except IL-12 with TLR-2 in zymosan-treated LCs were also demonstrated for the first time.
Generation of monocytes-derived LCs and stimulation of LCs with zymosan
Mononuclear cells were collected from healthy blood donors (n=10) by apheresis (Key Biologics, LLC, Memphis, TN). We numbered the subjects in a chronological order. Peripheral blood mononuclear cells (PBMCs) were isolated using the ficoll density gradient centrifugation and CD14+ monocytes were negatively isolated from PBMCs using Monocyte Isolation Kit II (Miltenyi Biotec, Auburn, CA). The isolated monocytes (106 ml-1) were cultured in RPMI-1640 medium supplemented with GM-CSF (1000IU ml-1), IL-4 (1000IU ml-1), TGFβ- 1(10ng ml-1, Invitrogen, San Diego, CA, USA) and equal concentration of cytokines were replenished on third day. Seven days later, monocytes were successfully converted into MoLCs. The identification of LCs was performed by detecting the expression of CD1a (eBioscience, San Diego, CA) and Langerin (Beckman-Coulter, Brea, CA) using FACS Fortessa (University of Arkansas for Medical Sciences Microbiology and Immunology Flow Cytometry Core Laboratory) and FACS Diva software (BD Biosciences, San Jose, CA).
LCs were stimulated with zymosan (10 μg ml-1; InvivoGen) [10]. Meanwhile, the cells without stimulation were set as negative control. After 8 h and 24 h stimulation, the cells were harvested for RNA extraction separately.
RNA extraction and cDNA synthesis
Total RNA was extracted with an RNeasy kit (Qiagen, Valencia, CA). All the RNA samples were treated with DNase I (Promega, Madison, WI) to remove potential residual DNA before RNA samples were reverse transcribed into cDNA using SuperScript III first-strand synthesis system (Invitrogen) for Quantitative PCR. The reaction system consisting of 10 × buffer 2 μl, dNTP (10 mM) 2 μl, MgCl2 (25 mM) 2.8 μl, Oligo dt (50 mM) 0.25 μl, R hex (50 μM) 0.5μl, RNase OUT (40 U μl-1) 0.2 μl, DNase (1 U μl-1) 0.75 μl, double distilled water (DDW) 1.5 μl, RNA 10 μl (0.1 μg). After all the RNA samples were performed to remove RNase and DNA under condition: 37ºC for 15 min: 70ºC for 12 min, 0.5 μl SuperScript III reverse transcriptase (200 U μl-1) was added to synthesize cDNA. The conditions for reverse transcription are as follows: 25ºC for 10 min; 50ºC for 50 min; 70ºC for 15 min.
Quantitative Real-time PCR and the sequencing of PCR products
Quantitative real-time PCR (qRT-PCR) was performed to determine the mRNA levels of TLR-1, TLR-2, TLR-4, TLR-9 and cytokines including IL-1β, IL-6, IL-8, IL-10, IL-12, IL-23, IL-27, IFN-γ and TNF-α. All the primers were described by Wang et al. [11]. MRNA was considered to be detectable when amplification of the corresponding cDNA was demonstrated. The results of threshold cycle numbers were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH), a human housekeeping gene. Relative quantification of the target gene relative to GAPDH was determined using 2-ΔΔCt Calculation method as described by Arocho [12]. In brief, ΔΔCt was calculated as ΔΔCt=[Ct (target gene)-Ct (GAPDH)] treated LCs-[Ct (target)-Ct (GAPDH)] not treated LCs at 8 h. The numerical value of ΔΔCt was then used in the 2-ΔΔCt Calculation. The final result represents the expression of target gene in treated LCs relative to the untreated LCs.
The PCR reaction system included RNA free H2O 3.25 μl, SYBR Green 6.25 μl, forward primer 0.25 μl, each reverse primer 0.25 μl, 2.5 μl diluted cDNA sample and was under following conditions: 95ºC for 3 min; 95ºC for 10 s; 60ºC for 30 s, total 40 cycles. PCR testing for each cDNA sample was performed in duplicate.
After gel-purification, the amplifications of the desired products were confirmed by DNA sequencing technique. Data are not shown.
IL-12p70 protein detection by ELISA
Cell-free supernatants of LCs treated with zymosan for 24 h were collected and the concentration of cytokine IL-12p70 protein was determined by an IL-12p70 high sensitivity ELISA kit (eBioscience). Values from media only wells were subtracted from experimental wells. The assay for each samples were performed in duplicate.
Correlation coefficient of cytokines and TLRs
We studied correlation coefficient of mRNA expression levels of cytokines and TLRs between 8 h and 24 h, and between cytokines to study mutant effects by formula below.
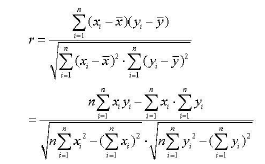
Statistical analysis
All statistical analyses were performed using SPSS for Windows version 19 (SPSS Inc., IBM Company,USA). ANOVA analysis was performed to analyze the differences in the mRNA level of TLR and cytokines among groups. Differences with p-values of less than 0.05 were considered to be significant.
Phenotypic analysis of LCs
The CD14+ cells induced using cytokines were shown expression of CD1a and Langerin, which demonstrated successful conversion of monocytes to LCs (data not shown).
The mRNA level of TLRs in LCs treated with zymosan
To investigate the effects of zymosan on TLRs in LCs, the mRNA level of 4 TLRs (TLR-1, 2, 4, 9) were examined by qRT-PCR. LCs from ten subjects were treated with zymosan and examined. All TLRs examined were detectable in LCs from all subjects. TLR-2 was up-regulated at the mRNA level (p<0.05) at 8 h (Figure 1A), while TLR-1 and TLR-4 were significantly down-regulated (p<0.05) at 8 h and 24 h (Figures 1B and 1C). The changing extent was slight. There was no significant difference in TLR-9 at mRNA level (Figure 1D).

Figure 1: The mRNA level of TLR-2 in LCs; B) The mRNA level of TLR-1 in LCs; C) The mRNA level of TLR-4 in LCs; D)The mRNA level of TLR-9 in LCs.
The mRNA level of cytokines in LCs treated with zymosan
To investigate the effects of zymosan on cytokines in LCs, the mRNA level of 7 cytokines (IL-1β, IL-6, IL-8, IL-10, IL-12, IL-23 and TNF-α) were examined. Total 10 samples of LCs were treated with zymosan and mRNA levels of 7 cytokines were examined by qRT-PCR. After gel-purification, the amplifications of the desired products were confirmed by DNA sequencing technique. Among all cytokines detected, some showed significantly increased mRNA level, while others were almost not affected. IL-1β was the most induced cytokine since the expression in all samples were increased dramatically with statistically significant difference between control and treatment group (p<0.05) (Figure 2A). IL-6, IL-8, IL-10 and IL-12 were increased significantly at 8 h and 24 h (Figures 2B-2E). IL-23 and TNF-α were shown enhanced in samples at 8 h only (p<0.05) (Figures 2F and 2G).

Figure 2: The mRNA level of IL-1β in LCs; B) The mRNA level of IL-6 in LCs; C) The mRNA level of IL-8 in LCs; D) The mRNA level of IL-10 in LCs; E) The mRNA level of IL-12 in LCs; F) The mRNA level of IL-23 in LCs.
IL-12p70 protein detection by ELISA
IL-12p70 as biologically active form of IL-12 directs Th1 the polarization in response to microorganism. To investigate the potential of zymosan to polarize Th1 response, its effect on active form of IL-12 in LCs, the level of IL-12p70 protein was examined by ELISA. Cell-free supernatants from LCs treated with zymosan for 24 h were analyzed for the presence of IL-12p70 protein. The results of 24 h protein concentration showed there was no significant difference in IL-12p70 expression (Figure 3).

Figure 3: The IL-12p70 protein secreted by LCs.
Correlation coefficient of cytokines and TLRs
Correlation coefficient of mRNA expression levels of cytokines and TLRs between 8 h and 24 h (Table 1) were determined. Among TLRs, TLR-9 was showed highly positively correlated (r>0.8) with time, which means the higher expression level at 8 h, the higher level at 24 h. Other TLRs (1, 2, 4, 6) showed moderately positively correlated (0.8>r>0.3). As for cytokines, IL-8 was highly positively correlated (r>0.8) with time. IL-10, IL-12, IL-23 and TNF-α were moderately positively correlated (0.8>r>0.3). While IL-1β, IL-6 and IFN-γ were not significantly correlated (0.3>r>-0.3), which indicates their expression levels may depend on interactions with other cytokines instead of time.
| Cytokines | correlation coefficient (r) |
|---|---|
| IL-1β | -0.14743 |
| IL-6 | 0.10027 |
| IL-8 | 0.81944 |
| IL-10 | 0.73948 |
| IL-12 | 0.77243 |
| IL-23 | 0.60360 |
| IFN-γ | -0.20940 |
| TNFα | 0.52340 |
| TLR-2 | 0.65587 |
| TLR-6 | 0.64386 |
| TLR-1 | 0.71932 |
| TLR-4 | 0.33267 |
| TLR-9 | 0.82665 |
Table 1: Correlation coefficient of mRNA expression levels of cytokines and TLRs between 8 h and 24 h.
Further calculation of correlation coefficient of cytokines at 8 h and 24 h were in matrix to study mutual effects of cytokines (Tables 2 and 3). At 8 h, the expression level of IL-1β was not correlated with the expression of other cytokines. The moderately or no correlation was shown for the expression of other cytokines (Table 2). At 24 h, nearly all cytokines were positively correlated (r>0.3), except for IL-12. This result suggested that inflammatory and non-inflammatory cytokines changed expression in a balanced style, while IL-12 was increased in all samples in our test regardless of other cytokine levels.
| IL-1β | IL-6 | IL-8 | IL-10 | IL-12 | IL-23 | IFN-γ | TNF-α | |
| 1 | 0.20637 | 0.19280 | 0.19206 | 0.13779 | -0.11830 | 0.22617 | 0.23447 | IL-1β |
| 1 | 0.09958 | 0.60274 | 0.50172 | 0.04107 | 0.66800 | 0.68804 | IL-6 | |
| 1 | 0.80755 | -0.0297 | 0.77742 | 0.08087 | 0.49694 | IL-8 | ||
| 1 | 0.38541 | 0.60900 | 0.50806 | 0.79088 | IL-10 | |||
| 1 | -0.01328 | 0.80151 | 0.42012 | IL-12 | ||||
| 1 | 0.24483 | 0.30817 | IL-23 | |||||
| 1 | 0.42858 | IFN-γ | ||||||
| 1 | TNF-α |
Table 2: Correlation coefficient of mRNA expression levels among cytokines at 8 h.
| IL-1β | IL-6 | IL-8 | IL-10 | IL-12 | IL-23 | IFN-γ | TNF-α | |
| 1 | 0.93609 | 0.92446 | 0.82513 | -0.20791 | 0.90683 | 0.67991 | 0.92621 | IL-1β |
| 1 | 0.95903 | 0.91724 | -0.09620 | 0.95204 | 0.77213 | 0.96050 | IL-6 | |
| 1 | 0.94201 | -0.15813 | 0.96628 | 0.69433 | 0.93747 | IL-8 | ||
| 1 | 0.02604 | 0.90542 | 0.66811 | 0.88062 | IL-10 | |||
| 1 | -0.09604 | 0.10961 | -0.06469 | IL-12 | ||||
| 1 | 0.66654 | 0.92351 | IL-23 | |||||
| 1 | 0.87520 | IFN-γ | ||||||
| 1 | TNF-α |
Table 3: Correlation coefficient of mRNA expression levels among cytokines at 24 h.
Correlation coefficient of mRNA expression levels between cytokines and TLRs at 8 h and 24 h were calculated at same time (Tables 4 and 5). At 8 h, positive relations included: TLR-6 and TLR-1 with IL-6 and IFN-γ; TLR-4 with IL-23; TLR-2 with all cytokines tested except for IL-12; TLR-9 with IL-1β (highly). At 24 h, positive relations included: TLR-4 with IL-23; TLR-2 with all cytokines tested except for IL-12. This result shows two things: first, cytokines and TLRs have a mutual promotion effect; second, the effect is possibly short-term cause less positive relations at 24 h than at 8h.
| IL-1β | IL-6 | IL-8 | IL-10 | IL-12 | IL-23 | IFN-γ | TNF-α | |
| -0.15615 | -0.00918 | 0.13371 | 0.03366 | -0.02409 | 0.67524 | 0.37227 | -0.11218 | TLR-4 |
| 0.09862 | 0.50761 | -0.11235 | 0.04876 | -0.09243 | 0.21423 | 0.30146 | 0.05245 | TLR-6 |
| 0.21713 | 0.70602 | -0.12523 | 0.23009 | 0.21197 | 0.06957 | 0.54218 | 0.27254 | TLR-1 |
| 0.32212 | 0.47876 | 0.40828 | 0.57159 | 0.19588 | 0.49438 | 0.54683 | 0.44411 | TLR-2 |
| 0.89244 | -0.01303 | 0.10332 | 0.03373 | -0.01553 | -0.32068 | -0.01798 | 0.05131 | TLR-9 |
Table 4: Correlation coefficient of mRNA expression levels between cytokines and TLRs at 8 h.
| IL-1β | IL-6 | IL-8 | IL-10 | IL-12 | IL-23 | IFN-γ | TNF-α | |
| 0.14606 | 0.26305 | 0.27474 | 0.11712 | -0.04271 | 0.32691 | 0.26795 | 0.26713 | TLR-4 |
| -0.21255 | -0.14008 | -0.33606 | -0.26959 | -0.16154 | -0.27825 | -0.03599 | -0.22643 | TLR-6 |
| 0.00099 | 0.08148 | -0.11844 | -0.21949 | -0.06316 | -0.09932 | 0.23964 | 0.00596 | TLR-1 |
| 0.35030 | 0.49744 | 0.41950 | 0.39813 | -0.25531 | 0.43091 | 0.39950 | 0.38130 | TLR-2 |
| 0.16362 | 0.04981 | 0.17781 | 0.24427 | -0.21232 | -0.04473 | -0.07387 | 0.04902 | TLR-9 |
Table 5: Correlation coefficient of mRNA expression levels between cytokines and TLRs at 24 h.
Membrane-bound PRRs including TLRs and CLRs on cells of the innate immune system provide a link between innate and adaptive immunity. The interaction of PRRs and the corresponding PAMPs can induce the release of activation signals and trigger the synthesis and secretion of cytokines and co-stimulatory molecules necessary for the initiation of adaptive immune responses. TLRs usually recognize and bind with ligands such as lipopolysaccharide (LPS) [13], zymosan [14,15], Neospora caninum [16] and then induce APCs activation and Th1 programming during infection [17]. Among all TLRs, TLR-2 plays a significant role in macrophages activation and works as homodimer [18] or heterodimer with TLR1 or TLR6 [19] to recognize diverse PMAPs. A cascade of events occurs after PAMP recognition by TLR- 2, which activate host defense mechanisms and initiate an adaptive immune response in MyD88-dependent signaling pathway [20]. In the wide array of TLR-2 ligands, zymosan was proved to be a strong stimuli in diverse recent studies [21,22]. Similarly, in our experiment, zymosan induced significant increase in TLR-2 of human LCs though the increasing fold of TLR-2 is less than 5 fold. Also, it demonstrated positive relations with nearly all cytokines at 8 h and 24 h, which indicate TLR-2 has done a positive effect in this process. Although mRNA levels of other TLRs didn’t rise in this test and some of them even decrease, it can’t be ruled out that TLRs may also participate in response of LCs to zymosan by higher affinity or more synthesis of second messengers in LCs to help with downstream immune modulation. Though the effect of zymosan with low concentration (10 μg ml-1) on LCs was investigated in this paper, a dose-response needs to be evaluated in the future study.
CLRs on dendric cells, such as Dectin-1, Dectin-2 and Mincle, can recognize carbohydrate moieties such as β-(1, 3) glucan and mannose [23,24] on antigens in a Ca2+-dependent way and trigger a series of downstream immune responses [25,26]. A recent study indicated DCs treated with zymosan showed significant increase of TNF-α secretion mediated by Dectin-1 receptor, one type of CLRs [27]. In our previous study, it was demonstrated that zymosan can induce higher expression level of CLRs including Dectin-2, Mincle and MRC-2[9], which indicated CLRs on LCs can timely response to zymosan not only by increasing number of receptors, but also activating cytokines secretion. It has been proved that Dectin-1 (a kind of CLRs) can cooperate with TLRs system to generate inflammatory cytokines production in response to fungal antigens [28].
ENK et al. proved LCs of mice can respond to several cell-contact antigens and induce evidently increased IL-1β mRNA transcription [29]. Dearman reported that murine DCs cultured with zymosan can produce marked expression of cytokine IL-1β, IL-6, TNF-α and IL-10 [30]. Similarly, our results demonstrated zymosan can induce significantly higher level of IL-1β mRNA in human LCs, which may finally contribute to early differentiation of DCs and influence their later functions [31]. As pro-inflammatory cytokines, IL-6 and IL-8 mRNA were increased in all samples in our study. Functionally, IL-6 is responsible for quick induction of granzyme B production in CD8+ T cells [32], while IL-8 participates in leukocyte recruitment and activation, especially neutrophils [33]. Combined with correlation coefficient calculation in our study, cytokines have mutual promotion effect not only among themselves, but also with TLRs-a subset of membrane-bound PRRs. This suggests zymosan can effectively enhance the activity of several types of immune cells.
Dillon et al. reported zymosan is a potent stimulus for IL-10 production through a Syk-dependent signaling pathway [34] that conveys signals different from those triggered by TLR2, which may lead to IL-12p70 induction. As the only anti-inflammatory cytokine, the ascending expression of IL-10 mRNA was demonstrated in 8 of all 10 samples in this study. On one hand, the result means zymosan can induce a balanced cytokine network reaction since IL-10 involves in inhibiting pro-inflammatory cytokines synthesis and preventing distensible tissue lesions related to immune responses [35,36]. On the other hand, it may also indicate a way of immune escape when fungi invades body environment because it has been shown that immune evasion mechanism of invaders like HIV and mycobacterium tuberculosis has strong relationship with a higher IL-10 expression [37,38].
IL-12 and IL-23 belong to one family due to a common subunit IL-12p40 and both play key role in cell-mediated immunity by enhancing cytotoxicity of natural killer cells and promoting Th1 cell differentiation [39,40]. In our test, all samples showed higher IL-23 and IL-12p40 mRNA level, which indicated that zymosan can strengthen T cell-mediated immunity in human. However, the detectable IL- 12p70 protein didn’t increase in any group, which was similar with the reports by Dillon [34] and Wei et al. [41]. It may be explained in the way: zymosan can’t induce IL-12p35 mRNA expression and this led to failure of combination of IL-12p40 and IL-12p35 subunits to form active IL-12p70 protein [40]. TNF-α mRNA level was measured higher in 9 samples. This may lead to improvement of host defence to bacterial, viral and parasitic infections as TNF-α works [42].
In preliminary work of Ainai A et al. it was proved that zymosan facilitates the uptake of HA vaccines prepared from influenza virus by BM-DCs [43]. And zymosan has been used as an adjuvant of nasal influenza vaccine and DNA vaccine for HIV-1 respectively in reliable medical researches [43].Overall, these studies demonstrate that zymosan has been applied successfully as an injection adjuvant. The results of this study further explain the mechanisms of cutaneous or mucosal immune reaction against fungal infection and the mechanisms of zymosan to improve human epidermal and mucosal immunity. Considering our results, it is indicated to be used as an adjuvant widely in intracutaneous vaccines since LCs can be activated. In clinical work, it also can be used and be used to enhance immunity of patients if necessary.
This study was supported by grants from the National Natural Science Foundation of China (No.81472439, No.81101989).