Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2012) Volume 3, Issue 1
Berried females of Portunus pelagicus (Blue swimmer crab) were collected from the wild and the zoea 1 obtained from a single berry were stocked in 200 litre tanks at the rate of 25 numbers/mL. In order to study the independent effects of temperature and salinity, the larvae were reared in three different temperatures (26°C, 28°C and 30°C) at a fixed salinity of 35 ppt and in three different salinities (25 ppt, 30 ppt and 35 ppt) at a fixed temperature of 28°C. Feeding was carried out with microalgae, rotifers, moina and formulated shrimp feed, which varied with stage. The survival rates at each stage in all the trials were assessed by random sampling. The most ideal temperature and salinity among those studied were calculated in relation to the overall survival rate and larval development period. The data pertaining to both temperature and salinity manipulated investigations were found to be significant (P < 0.01) when subjected to one-way ANOVA. Among the tested temperatures, the highest average survival (14.12 ? 0.31%) and the lowest average development period (14.67 days) were obtained at 30 ± 0.1°C. Among the salinity values tested, the highest mean survival rate (06.89 ± 0.06%) and the lowest average development period (14.33 days) were obtained at 35 ppt.
Keywords: Portunus pelagicus, larval rearing, larval survival, larval developmental period
Survival and longevity of marine invertebrate larvae are influenced by abiotic factors such as water temperature and salinity and by biotic factors such as food availability, food quality and predation [1]. Crustacean larval development occurs within a narrow range of environmental parameters such as temperature and salinity [2]. Many authors like Baylon et al. [3] have investigated the influence of salinity on the survival and developmental period of mud crab (Scylla serrata) larvae under controlled conditions and got the best results at 32 ppt, which was the highest among the values tried.
Laboratory studies conducted by Nagaraj [4], with notionally optimal food and water quality, have generally shown that temperature has the greatest effect on the development period of the larvae, while survival can be influenced by both temperature and salinity. Temperature is widely held to be a critical factor for the development of marine invertebrate larvae [5]. Temperature changes profoundly influence the development of decapod larvae. Many studies have demonstrated the effects of temperature change on the developmental period of instars, their morphology, feeding rate, individual size, incidences of deformity and survival rate [6]. Several studies have been carried out to understand the effect of temperature, both as a single factor and in combination with salinity. The combined effect of temperature, diet and stocking density was studied on the larval survival of the marine ornamental crab Mithraculus forceps [7]. The larval survival rate increased to over 500% when the temperature was increased by 10°C in the coconut crab Birgus latro [8]. A decrease in the larval development period with an increase in temperature was reported in the snow crab Chionoecetes opilio by Takayuki et al. [9]. Temperature changes also produce measurable alterations in the directional responses to light (phototaxis) and gravity (geotaxis), and the activity of crustacean larvae in general [10].
Wide fluctuations in salinity were found to be critical for the growth and survival of larvae of several marine crustaceans. Salinity changes always have stressful effects on aquatic organisms, especially crustaceans and tolerance to such changes is often erratic during ontogeny [11]. Salinity fluctuations were seriously found to affect moulting, metamorphosis time and larval survival rates in the mud crab Scylla paramamosain [12]. In the early juveniles of P. pelagicus, the survival rate was less at salinities less than 15 ppt and more than 45 ppt; bulk of the larval mortality occurred at these extreme salinities due to ‘molt death syndrome’ [13]. Investigations have been carried out on the combined influence of diet and salinity on the larval survival and development of several crabs like S. serrata [3] and S. paramamosain [14]. A complex relationship was found to exist between temperature and salinity in Artemis spp. where a change in one factor would lead to an alteration on the effects of the other [15].
The marine blue swimmer crab P. pelagicus is a candidate species for culture because of its fast growth, attractive appearance and taste. The species is available throughout the coast of India, especially in the south-east and the south-west regions and breeds round the year. As there are no commercial hatcheries for the species, the farmers are forced to depend on juvenile crabs from the wild for culture practices. Difficulties in obtaining juveniles from the wild, their non-uniform sizes and concerns of stock depletion due to overexploitation have necessitated research activities to frame out hatchery technology for the species. Knowledge on the influence of various abiotic parameters, especially temperature and salinity on the rate of survival of the larvae and the development period is of extreme significance in standardising the hatchery technology and developing a conservation management strategy for the species. However, studies on the effect of these parameters on the larvae of P. pelagicus from Indian waters are scanty. So the present work is taken up, to study the rates of survival and the periods of development of the larvae of P. pelagicus from the south-east coast of India in relation to temperature and salinity of the rearing water. The study also attempts to find out the optimal range of these parameters for each larval stage.
Source of the larvae
Five female P. pelagicus of size range 140 - 160 mm carapace width with light yellow berry were collected from the Gulf of Mannar off Mandapam (9°09’ N, 78°44’ E). Animals were brought live to the laboratory, given a prophylactic dip in 200 ppm formalin for 30 minutes [16] and stocked in 1000 L FRP (Fibreglass Reinforced Plastic) tanks with black interior and sandy bottom (hundred millimeters thickness) for acclimatization (2 days). Three of the most active animals were weighed (to the nearest 1 g) and stocked separately in 500 litre FRP tanks for the study. Feeding was done with raw clam (Meritrix meritrix) and cuttlefish (Sepia pheraonis) meat (1 : 1), ad libitum and the excess feed was removed early morning every day. Water exchange (70%) and water quality monitoring were carried out daily. The temperature, salinity, pH and photoperiod were maintained at 28 ± 0.1°C, 35 ppt, 8.1 ± 0.1 and 12hL:12hD (hL/hD = hours light/dark), respectively. A sandy substratum of four inches thickness was provided in all the tanks for refuge and to prevent egg loss [17]. The pH of water was adjusted by adding 1 normal solution (N) of sodium carbonate or 1N hydrochloric acid when required. Tanks were covered with black sheets after the photophase. The berry colour was examined daily and when the berry became dark grey in colour, the animals were transferred to cylindro-conical FRP hatching tanks of 500 L capacity with white interior.
Larval stocking and maintenance
Once the eggs were hatched, the zoeae 1 (identified according to Shinkarenko [18]) were collected using 150 μm mesh sieve and the most active ones (identified by their vigorous swimming activity in opposite direction in a whirl), from the same brood were stocked after rinsing in fresh seawater in 200 L rectangular FRP tanks at a density of 25 numbers/L (i.e. 5000 larvae/tank, as this stocking density gave better results in earlier trials) in triplicate with controls, for each experimental value of a parameter. Dead larvae and feed waste were siphoned out prior to the daily water exchange (70%). Abiotic parameters were monitored daily and continuous aeration was provided in all the tanks. The survival rates at each of the five larval stages such as zoea 1 (Z1), zoea 2 (Z2), zoea 3 (Z3), zoea 4 (Z4) and megalopa (M) (identified as per Shinkarenko [18]) were calculated by daily random sampling (mortality due to any reason other than temperature or salinity variation is assumed to be the same in all the replicates of the values tested under each parameter). At first crab instar (C1), the larvae were harvested using 500 μm mesh nets for estimating the final survival rate. The periods of development of the larvae (from Z1 to C1) for each replicate under the temperature and salinity treatments were also recorded.
Larval feeding protocol
Zoeae 1 and 2 were fed twice daily with Skeletonema costatum (50000 numbers/mL) and Brachionus plicatilis (25 numbers/mL). The larvae were also fed with formulated prawn feed (Frippak 2 CD-INVE, Belgium) at a rate of 0.5 g/1000 L of water (four times daily). Zoeae 3 and 4 were also fed with S. costatum, B. plicatilis and formulated prawn feed (at densities 50000 cells/mL, 30 numbers/mL and at a rate 0.5 g/1000 L of water, respectively). The frequency of feeding was maintained as in the case of Z1 and Z2. Megalopae were fed daily once with Moina macrura and newly hatched Artemia nauplii maintaining cell densities at 3 numbers/mL and 5 numbers/mL, respectively and twice daily with freshly prepared egg custard at the rate of 0.5 g/1000 L of water. Rotifers and M. macrura were collected from the wild and mass cultured in 1000 L FRP tanks, feeding on microalgae. Microalgal (Chlorella sp. and S. costatum) inoculums were obtained from the live feed laboratory of the Central Marine Fisheries Research Institute, Mandapam and scaled up in 1000 L FRP tanks.
Experimental layout
Two parameters viz. temperature and salinity were investigated separately. One set of experiment investigated the effect of three different temperatures and another set investigated the effect of three different salinities.
The values selected for studying the effect of different temperatures were: Treatment 1 (T1)/control = 26 ± 0.1°C, Treatment 2 (T2) = 28 ± 0.1°C and Treatment 3 (T3) = 30 ± 0.1°C (T3/treatment 3). Temperatures were maintained by keeping the tanks in chilled room and then increasing the water temperature to the desired levels using aquarium heaters with thermostats (AZOO, USA 100 W). The salinity, pH and photoperiod were maintained at 35 ppt, 8.1 ± 0.1 and 12hL : 12hD throughout the experiment.
The values selected for studying the effect of different salinities were: T1 = 25 ppt, T2 = 30 ppt and T3/control = 35 ppt. Salinity of water was adjusted by mixing freshwater and seawater. The temperature, pH and photoperiod were maintained at 28.0 ± 0.1°C, 8.1 ± 0.1 and 12hL : 12hD throughout the study. Observations on feeding, growth, cannibalism, activity and survival were recorded every morning and evening.
Statistical analysis
The overall (Z1 to C1) survival rates and development period under different temperature and salinity treatments were statistically analyzed with one-way ANOVA. The data with significant difference (P < 0.01) were then subjected to Post Hoc Test of multiple comparisons using ‘Fisher's Least Significant Difference’.
Complete larval development took place in all the replicates under the parameters studied. The survival rate fell almost gradually with time and stages of the larvae at all the temperatures and salinities studied.
Larval survival rate and development period in relation to temperature
The average overall survival rates at 26 ± 0.1°C, 28 ± 0.1°C and 30 ± 0.1°C were 6.85±1.02%, 11.88±0.14% and 14.12±0.31%, respectively. The overall survival rates at 28 ± 0.1°C and 30 ± 0.1°C were significantly (P < 0.01) different from that at 26 ± 0.1°C. The survival rates at each stage also differed with temperature (Figure 1). The average period of larval development was found to be 18, 17 and 15 days at 26 ± 0.1°C, 28 ± 0.1°C and 30 ± 0.1°C, respectively. The development periods at 28 ± 0.1°C and 30 ± 0.1°C were significantly (P < 0.01) different from that at 26 ± 0.1°C. The development period of each stage also varied slightly with temperature (Figure 2).
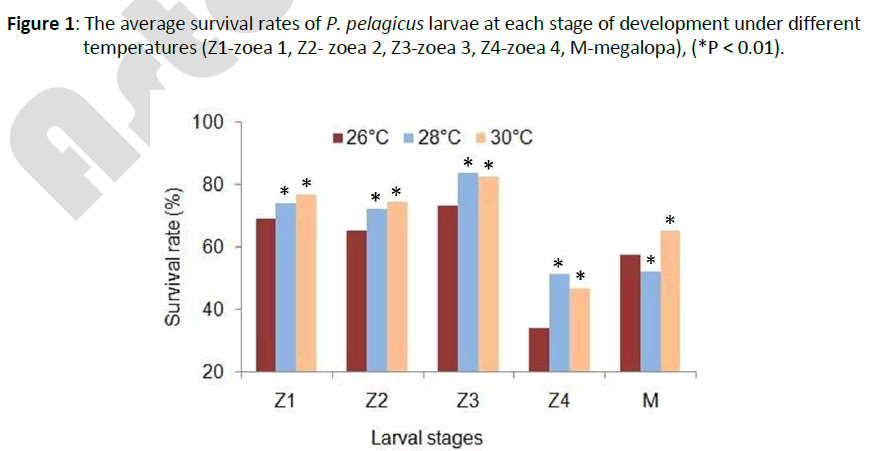
Figure 1: The average survival rates of P.pelagicus larvae at each stage of development under different temperatures (Z1-zoea 1, Z2- zoea 2, Z3-zoea 3, Z4-zoea 4, M-megalopa), (*P < 0.01).
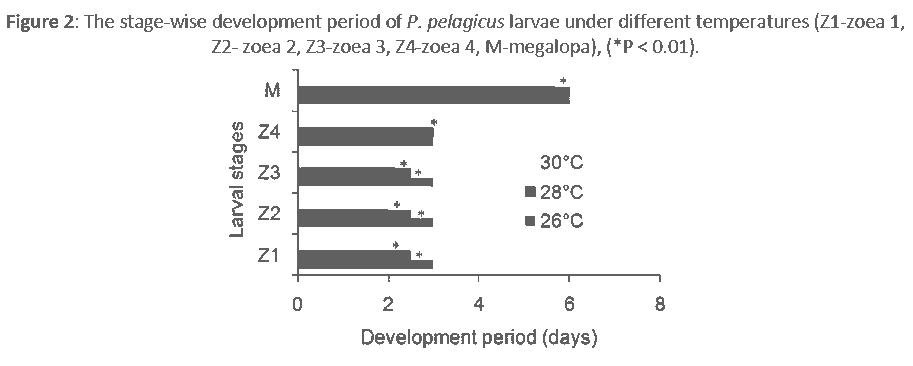
Figure 2: The stage-wise development period of P. pelagicus larvae under different temperatures (Z1-zoea 1, Z2- zoea 2, Z3-zoea 3, Z4-zoea 4, M-megalopa), (*P < 0.01).
Larval survival rate and development period in relation to salinity
The average overall survival rate at 25 ppt, 30 ppt and 35 ppt were 5.42 ± 0.35%, 6.54 ± 0.25% and 6.89 ± 0.06%, respectively. The average overall survival rate at 25 ppt was significantly (P < 0.01) different from those at 30 ppt and 35 ppt. The survival rate at each stage also differed with salinity (Figure 3). The average period of larval development was found to be 18, 16 and 15 days at 25 ppt, 30 ppt and 35 ppt, respectively. The average period of larval development at 25 ppt was found to be significantly different (P < 0.01) from that at 35 ppt and the remaining pairs were not significant. The development period of each stage also varied slightly with salinity (Figure 4).
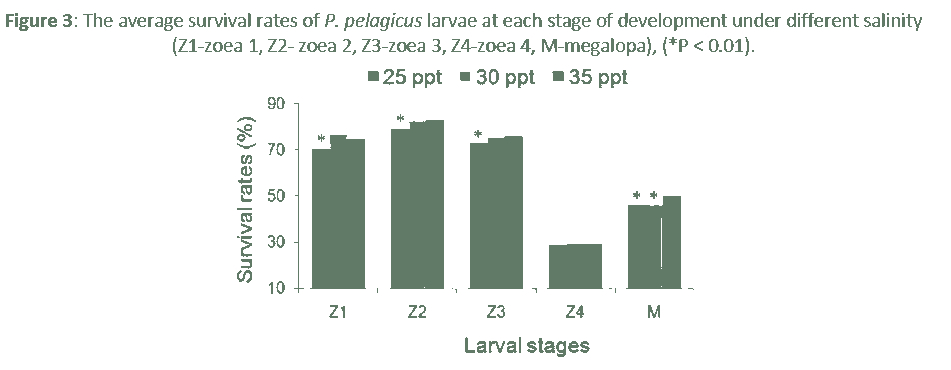
Figure 3: The average survival rates of P. pelagicus larvae at each stage of development under different salinity (Z1-zoea 1, Z2- zoea 2, Z3-zoea 3, Z4-zoea 4, M-megalopa), (*P < 0.01).
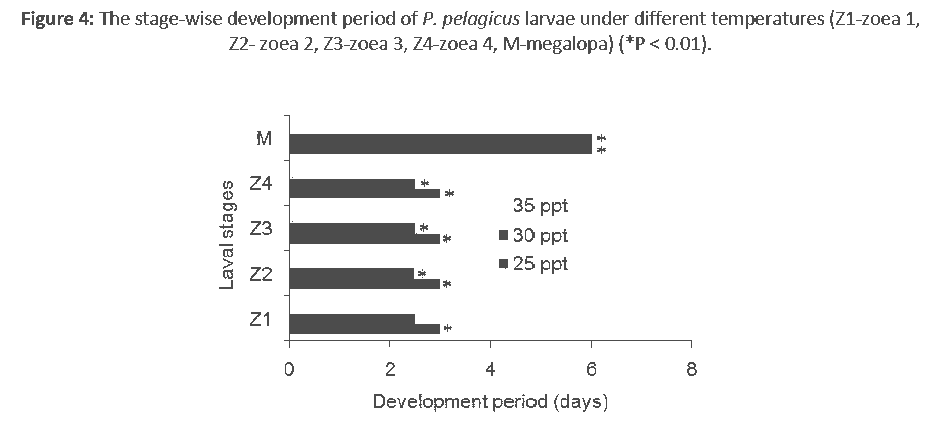
Figure 4: The stage-wise development period of P. pelagicus larvae under different temperatures (Z1-zoea 1, Z2- zoea 2, Z3-zoea 3, Z4-zoea 4, M-megalopa) (*P < 0.01).
According to Sorgeloos and Leger [19], rearing techniques, disease and nutrition are the three main areas of research that have supported commercial production of marine fish and crustacean larvae. The physico-chemical characteristics of the larval rearing medium are of extreme significance since this form the environment of the larvae. Any intervention causing adverse alterations to the larval environment will badly affect the instar development rate and consequently the overall survival.
In the present study, each of the temperatures regimes tried influenced the overall larval survival rate differently. The three temperatures tested here were not in the extreme range and the maximum difference among the treatments was only 4°C. This shows that the larvae are very sensitive to even small differences in temperature of the larval rearing medium. The overall survival rate was found to increase with temperature. Similar increase in survival rate with the increase of temperature was reported in many other crabs like S. serrata [3] and S. paramamosain [20]. Since maximum overall survival rate was observed at 30°C, that temperature seemed to be the most suitable temperature for larval rearing of P. pelagicus among the temperature regimes selected in the present study. Similarly, 30°C or nearby values were reported to be the optimum temperature by several authors like Hoang et al. [20] for portunid crab larval rearing. However, the optimum temperature is likely to vary with geographical location and species. For instance P. pelagicus when reared at different temperatures within a range of 17 to 25°C showed highest survival at 25°C [5].
Adversities like mass mortality in some of the larval stages [5], low activity, low feeding, low growth and low survival [21] were reported when crabs were reared at low temperatures. However, no perceptible differences were observed in feeding, growth or activity among the larvae reared at various temperatures in the present study. Even at the lowest temperature tried (26°C), complete larval development to crab instars took place in all the replicates. This shows that 26°C is within the tolerable temperature range of the larvae of P. pelagicus.
The larval development period in crustaceans is chiefly dependent on the rearing water temperature [4]. In the present study, the lowering of temperature affected the larval development period adversely. The overall larval developmental period was found to decrease with increase in the temperature of the rearing medium. Similar inverse relationship between temperature and larval development period has been reported in B. latro [8] and the golden king crab Lithodes aequispinus [22].
In crustaceans, the enhancement of moulting at increased temperatures is due to the increased activity of the hormones [23]. So at higher temperatures, the rate of ecdysis increases resulting in shorter development periods. When the moulting is fast, the period under ‘moulting stress’ is very low. Lesser stress during moulting is likely to decrease the chances for the incidence of bacterial diseases. For example, vibriosis is caused by one or more Vibrio spp., which normally occurs in seawater and invades marine animals as opportunistic pathogens under stressful conditions [24]. The larvae do not feed when settled at the bottom for moulting, which would make them weak. Furthermore, when a larva is weak and settled, the risk of being cannibalised is also high. Cannibalism was evident in areas where the larvae aggregated, in the present study. The larvae aggregated mainly in places where the light intensity was higher and at the tank bottom. When settled, the cannibalism was comparatively lesser than when aggregated due to light. Mostly, the zoeae, which either moved with water current created by aeration or clung to the sides of the tanks survived better. All these point out to an increased survival rate at a higher temperature, within the limit, which is species specific. Extreme temperatures (high or low) are thought to affect both moult initiation and duration of moult cycle depending on the tolerance of the species [25]. Since studies on larval bacterial profile or cannibalism were not a part of the present work, the possibility of mortality due to these two reasons was assumed to be equal in all the replicates under the temperatures studied.
In crabs, salinity may be one of the key factors affecting embryogenesis, energetics, and larval quality [26]. The effects of salinity on the larval survival and development period have been investigated in many crabs like the Chinese mitten crab Eriocheir sinensis [27], S. serrata [3] and the estuarine crab Chasmagnathus granulata [28]. The peaks in juvenile recruitment of crabs under Portunidae in temperate climates is related to water temperature, while in tropical populations juvenile abundance is related to salinity [29]. The significant difference in the overall larval survival rate and larval developmental period when reared at 25 ppt shows that the lowest salinity tested influenced both moulting and survival of the larvae differently. Though the salinities 30 ppt and 35 ppt also influenced the overall survival rates and larval development period, the highest survival rate and lowest larval developmental period were shown at 35 ppt, in the present study. Several authors have reported adverse effects on larvae and/or young ones when reared under low saline conditions. For example, osmotic stress has been reported to elicit increased oxygen consumption in the juveniles of the pacific blue shrimp Litopenaeus stylirostris [30] and ammonia excretion in the post-larvae of the Pacific white shrimp Litopenaeus vannamei [31]. These adverse metabolic activities are likely to alter the culture environment in closed systems. The larvae under stress get infected quickly and succumb to death. Since the culture system is also likely to get infected by then, the chances for mass mortality are extremely high. Mortality due to cannibalism and low feeding, which increase under these conditions, aggravates the situation.
Apart from the stress caused due to environmental variations, mortality at lower salinities is also possible due to the imbalance in osmoregulatory mechanism [32]. In decapods, the ability for osmoregulation at very low salinities generally develops only by late larval or early juvenile stages [33]. Gills are the important sites of active ion transport in adult decapods; larvae develop gills in the penultimate or ultimate zoeal stages [32]. Osmoregulation also depends on eyestalk factors that become functional during the late zoeal or megalopa stages as in the heavy marsh crab Sesarma reticulatum [34]. In the grapsid crab Armases miersii, the hyper-osmoregulatory capacity increased gradually from hatching to adult and the hypo-osmoregulatory capacity from megalopa to adult [33]. The organism’s response to the osmotic and ionic concentration of the medium could also vary with the moult stages and at different periods after ecdysis [35]. In the present study also, P. pelagicus larvae seemed to have only a limited capacity to survive variations in salinity.
The adverse effects reported in many crabs at lower salinities were not observed in the present study. The larvae in all the replicates at 25 ppt underwent complete larval development. This indicates that the lowest salinity attempted here falls within the tolerable range of the larvae of the species. Though studies on the effect of salinity on the osmoregulatory mechanism of the larvae was not a subject of the present study, the absence of mass/excessive mortality at any of the stages in the salinity regimes attempted indicates that the lowest salinity treatment investigated had a comparatively lesser effect on the osmoregulatory mechanism of the larvae of P. pelagicus. Extreme salinities (low/high) have been found to affect the overall larval survival rate in crabs. It might be either due to the increased energy expenditure for osmoregulation than that for growth [36] or use of proteins as a source of energy for osmotic functions, which makes them unavailable for growth [6]. However, obvious or sudden variations in the growth pattern were not observed in any of the salinity treatments in the present study. The larval development period was significantly higher for the lowest salinity tried in the present study. The period of larval development decreased with the increase in salinity as observed in E. sinensis [27]. The survival rates were found to be the lowest at the lowest salinity tried (25 ppt). However, the highest survival at each stage varied between 30 ppt and 35 ppt. In general, it is seen that the survival rate bears an indirectly proportional relationship with the development period.
Since even the lowest salinity regime tried in the present study was able to produce crab juveniles from zoea 1 though with a lesser overall survival rate and a comparatively higher developmental period, the larvae of P. pelagicus seem to be tolerant to a salinity range from 25 ppt to 35 ppt. The inference agrees with the report in which it is mentioned that the natural distribution of P. pelagicus in the Peel-Harvey Estuary in south-western Australia is restricted to a salinity range of 25-35 ppt [37]. Furthermore, the reports recommending a salinity range of 20-35 ppt for better growth and survival of the early juveniles of P. pelagicus by Romano and Zeng [13] also suggests the euryhaline nature of the species.
The present study provided valuable information on the temperature and salinity requirements for the larval rearing of P. pelagicus, especially in the Indian subcontinent. Maintenance of the larvae at its most suitable temperature-salinity combination (28-30°C and 30-35 ppt) is recommended for the maximum overall survival.
The authors declare that they have no competing interests.
All authors have contributed equally to this work.
The authors wish to express their sincere thanks to Dr. G. Syda Rao, Director, Central Marine Fisheries Research Institute (CMFRI), Cochin for his guidance and encouragement. Acknowledgement is also due to Dr. G. Gopakumar, Scientist-in-Charge, Regional Centre of CMFRI, Mandapam for the facility provided.