Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Review Article - (2018) Volume 6, Issue 4
Keywords: Bacteria; Compatible solutes; Drought; Fungi; Horizontal gene transfer; Native plants; Salinity; Soluble sugars; Trehalose
It has been known for a long time that soluble sugars are among the main metabolites that sustain plant growth and development. Over the last five decades, these solutes have received much attention in many living organisms under various stress conditions [1-8]. When crop plants and microorganisms are exposed to various types of stress under natural environmental conditions and/or as a result of industrial activities during oil and gas production; such conditions make the physicochemical and biological properties of the soil unsuitable, not only for crops directly but also for many microbial activities necessary to support crop productions [9,10]. As a common response, these plants sacrifice from their energy and reserves to build many solutes in their cells and tissues; these solutes serve as osmo-protectants to stabilize biomolecules. Also, many other physiological and biochemical parameters are altered [11]. In an early work [1], it was concluded that soluble sugars contributed about 30-50% of the osmotic adjustment in the leaves of many glycophytes. These soluble sugars include fructose, glucose, sucrose and possibly trehalose, which play significant roles as compatible osmolytes for osmoregulation, osmotic adjustment, and maintaining the growth and structure of plant tissues [12,13]. These roles depend on the growth stage, plant species, composition of the growth medium, and environmental conditions. In recent years, other soluble sugars like trehalose in some plants, have been revealed to play various roles in plant metabolism, especially under extreme environmental conditions that cover many aspects of plant growth, physiology and biochemistry [8]. Looking into the literature, little has been reported about the roles of soluble sugars in plants at the Arabian Gulf region, under natural environmental conditions during various stages of plant life cycle. This review discusses the possible roles of soluble sugars during all stages of plant growth and development, and special attention has been paid to some case studies in the Middle East in general and the Arabian Peninsula in particular. Since little has been published about the metabolism of soluble sugars in crops and native plants in this region, special attention has been paid to trehalose as a compatible solute and will be discussed in a separate section.
In fact, the results of the huge number of articles and studies on sugar contents have been inconsistent, as these solutes regulate plant structure and physiology in a complex manner [14], and their roles in plant metabolism are conflicting in various plant species under different stress conditions [15,16], despite that number of studies that have discussed their roles in plant life at different stages of growth and development [8,17]. There are many reports of such controversial results and conclusions (Table 1); for example, increasing soluble sugars was reported in wheat, sesame, maize, fenugreek, and alfalfa nodules [18-24], while other reports have shown either stable or decreasing concentrations of these solutes in the cotyledons of soybean, watermelon (Citrullus lanatus), variegated coleus (Coleus blumei Benth), and in maize as well [25-29]. The results and conclusions of these studies were attributed to many reasons: (a) the type of plant under investigation, (b) the environmental conditions adopted in the experiments, (c) the methods used to determine these sugars, (d) the stage of growth and age of plants, and (e) the plant organ or tissue under investigation [2,44].
| Enzyme | Reaction | Effect of Osmotic Stress | References |
|---|---|---|---|
| α – Amylase | Hydrolysis of starch | Decreased (Germination) | [30-33] |
| β- Amylase | Hydrolysis of starch | Decreased (Germination) | [33] |
| α– Amylase | Hydrolysis of starch | Increased | [34,35] |
| β- Amylase | Hydrolysis of starch | Increased | [35] |
| Invertase (β- fructosidase), Acid and Alkaline | Hydrolysis of Sucrose | Acid: not consistent, Alkaline: increased, Increased in leaves*, Decreased in roots* |
[36,37*] |
| Glucokinase (GK) | phosphorylation of glucose | Low activity | [37] |
| Phosphoglucoisomerase (GPI) | interconverting glucose 6-phosphate and fructose 6-phosphate | Low activity | [37] |
| pyruvate kinase (PK) | Transferring of a phosphate group from (PEP) to (ADP), yielding one molecule of pyruvate | Low activity | [37] |
| Glucose-6-phosphate dehydrogenase (G6PDH) | Participating in the pentose phosphate pathway | Decreased | [37] |
| PEPCase | CO2 fixation in the mesophyll tissue of C4 plants | Increased | [38,39] |
| Rubisco | CO2 fixation | Decreased | [40] |
| PEPCase | CO2 fixation in the mesophyll tissue of C4 plants | Increased | [38,39] |
| Rubisco | CO2 fixation | Decreased | [40] |
| Sucrose phosphate synthase | UDP – Glu. + Fr.-6-P→Sucrose -6-P + UDP | Decreased | [36] |
| Sucrose synthase | Biosynthesis of sucrose | Increased | [36] |
| Trehalose-6-phosphate phosphatase | Biosynthesis of trehalose | Increased | [41,42] |
| Trehalose-6-phosphate synthase | Biosynthesis of trehalose | Increased | [41,42] |
| Trehalase | Trehalose degradation | Decreased | [43] |
Table 1: Effect of osmotic stress (water stress or/and salinity) on enzyme activity of carbon metabolism.
In general, the following roles have been reported in most studies: (a) play various activities during seed germination, flowering, senescence etc., and possibly others under abiotic stresses [8], (b) protect the metabolic machinery of the cell against the osmotic constraints imposed by extreme environmental conditions, (c) function as signal molecules to regulate the different genes involved in many physiological and biochemical activities [13]. In fact, during the plant life cycle, two main stages can be recognized during which soluble solutes exhibit different trends of metabolism and accumulation [11], namely: (1) germination stage: In this stage, plants rely on stored macromolecules (carbohydrates, proteins and lipids) to provide the basic units, organic solutes of low molecular weight, to build new cells and to provide support against extreme environmental stresses, (2) growth and yield stages: In this stage, plants are dependent mainly on photosynthesis to provide most of the basic skeletons for metabolism and offer support against various types of threats of natural environmental conditions. This review aims to discuss the alleviating functions of soluble sugars in the physiology and biochemistry of plants under extreme environmental conditions, and the possible roles of microorganisms in providing some of these solutes to plants and their environments.
Germination stage: All seeds normally store macromolecules like carbohydrates, proteins and lipids that are hydrolyzed and degraded during the first stages of germination to support the metabolic activities of new plants [45]. These activities are significantly affected when plants are exposed to extreme environmental conditions like drought, salinity and extreme temperatures (high temperature). These are the major abiotic factors in the Arabian Gulf region that cause direct changes in the plant environments, resulting to a negative impact on the physiological and biochemical activities, thereby exerting deleterious alterations in the germination, vegetative growth and yield of most plants including crops. Such alterations depend mainly on the plant species and their genetic background, plant age, growth stage and the level of stress exerted [45-47]. Considering the different plant crops, Fenugreek (Trigonella foenum-graecum L.), cereals like barley (Hordeum vulgare L. ), wheat (Triticum aestivum L. ) seeds, and Sorghum bicolor L.; exhibited different responses under different osmotic stresses in terms of soluble sugars; especially glucose, fructose and sucrose [5,21,48,49]. Fenugreek seeds (rich in proteins) showed substantial increase in total soluble sugars in the seedling tissues as water stress increased around the seeds. This was explained mainly by the substantial accumulation of sucrose, while monosaccharides diminished with decreasing osmotic potential of the solutions around the seeds (Table 2). The sucrose content seemed controversial during seed germination, the formation of sucrose in many germinating seeds helps cope with various extreme environmental conditions, as it contributes actively to the osmotic potential of plant tissues and the osmotic adjustment process [8,21]. The accumulation of sucrose in fenugreek seedlings under such conditions might be the result of converting the carbon skeletons of nitrogenous compounds to soluble sugars, as well as from the considerable degradation of polysaccharides [21,49,50]. These data revealed some facts about the complex changes in biochemical activities during carbohydrate metabolism. The reduction in monosaccharides that has been reported in many studies was attributed to their utilization as respiratory substrates, since these studies have found that osmotic stress increases respiration rate, thereby consuming substantial amounts of these carbohydrate components [45,51-56]. Moreover, proline accumulates significantly to play important roles of osmotic adjustment and maintaining the plant cell machinery [57]. The amino groups and carbon skeletons could come from the degradation of proteins and the metabolic changes of carbohydrate fractions [58].
| Osmotic potential (MPa) | DRV | Sucrose | TRV |
|---|---|---|---|
| Control | 8.7 | 15.3 | 24.9 |
| -0.1 | 10.3 | 17.4 | 28.6 |
| -0.3 | 3.7 | 29.3 | 34.6 |
| -0.5 | 5.6 | 39.2 | 46.9 |
| -0.8 | 1.3 | 26.4 | 29.1 |
Table 2: Carbohydrate fractions (mgg-1DW) of fenugreek germinating seeds after five days exposure to different osmotic potentials of mannitol [21]. DRV: Direct Reducing Value, TRV: Total Reducing Value.
Considering seeds rich in carbohydrate like cereals (barley and wheat) with limited protein content in the aleurone layer, some authors [48,49] found that the total reducing value (TRV) and soluble sugars, diminished in the growing seedlings when exposed to salt stress. The decrease in soluble sugar contents was explained mainly by lowering both monosaccharides and sucrose, as both carbohydrate fractions had a similar effect in their contribution to the value of TRV (Table 3). In fact, the degradation of the stored carbohydrate and the following transamination reactions of 2-Oxoglutaric acid (α-keto-glutaric acid) would produce glutamate (a precursor of proline). This reaction is accelerated during the darkness conditions of seed germination, then by diminishing soluble sugars, the carbon skeleton would produce proline to achieve osmotic adjustment as well as maintain the machinery of plant tissues [48,49,59]. This argument was confirmed by some authors [5] in Sorghum bicolor L., they suggested that water stress or salinity around the seeds would increase the conversion of monosaccharides and sucrose into other soluble solutes like proline and polyols in the germinating seeds. This study showed clearly that while the osmotic potential of -1.86 MPa of NaCl and PEG around sorghum seeds lowered the germination percentage substantially, it accelerated the accumulation of soluble sugars in the seedling axis (Table 4).
| NaCl (dSm-1) | DRV | Sucrose | TRV | |||
|---|---|---|---|---|---|---|
| Wheat | Barley | Wheat | Barley | Wheat | Barley | |
| Control | 21.7 | 14.6 | 18.7 | 23.7 | 42.5 | 40.3 |
| 10 | 12 | 10.3 | 9.7 | 23.5 | 22.8 | 35.6 |
| 20 | 9 | 6.6 | 10.9 | 15.5 | 21 | 23.3 |
| 30 | 5.7 | 3.2 | 3.1 | 12.9 | 9.3 | 17 |
| 40 | 4.5 | 1.3 | 3 | 12.6 | 7.9 | 14.6 |
Table 3: The effect of salt stress on DRV and sucrose formation (mgg-1DW) in the seedlings of the Qatari local wheat (Triticum aestivum L), Doha cultivar and Jordanian barley (Hordeum vulgare L.), Rum cultivar after five days of treatment [48,49].
| Treatment | Sugar content (µgg-1 DW) | ||
|---|---|---|---|
| Embryos | |||
| Glucose | Fructose | Sucrose | |
| Control | 15 | 75 | 7 |
| NaCl | 75 | 210 | 15 |
| PEG | 35 | 135 | 10 |
| Endosperms | |||
| Treatment | Glucose | Fructose | Sucrose |
| Control | 25 | 55 | 16 |
| NaCl | 36 | 123 | 25 |
| PEG | 35 | 117 | 20 |
Table 4: Effect of osmotic stresses of NaCl and PEG (-1.86 MPa) on the soluble sugar contents in the germinated seed of Sorghum bicolor [5]. N. B. Total sugar content at 0 h of germination (after 6 h of imbibition) was 193 µgg-1 DW tissues.
The glucose and sucrose contents were higher in the embryos and endosperms after osmotic stress treatments, and the fructose levels were always higher than those two soluble sugars. Moreover, other compatible solutes like trehalose, proline and glycinebetaine might accumulate to take part in all the metabolic activities under stress conditions [10,11]. The degradation processes of macromolecules like polysaccharides and proteins are decreased gradually by lowering the osmotic potential of the growth medium around the seeds, and at the end, such processes are substantially reduced leaving many metabolites at their lowest levels. In fact, when the solutions around the seeds are at a very low osmotic potential, the activities of most hydrolytic enzyme systems were suppressed [49,55,60]. This might substantially reduce the levels of many compatible solutes, including proline and soluble sugars in the seedling axes.
It is very interesting to consider the changes in the contents of polysaccharides, proteins and their fractions in the germinated seeds at high stress levels (for example at: -0.8 MPa mannitol, the germination % of fenugreek seeds were 6.7%; and at 40 dSm-1 NaCl, the germination % of barley seeds was 15%) [21,49]. Although, controversial results have been obtained regarding the sucrose content in these two plants, the degradation of macromolecules was very low because of the great inhibition of the activity of hydrolytic enzymes which affects all the metabolic activities, including the degradation processes during the germination stage [45]. The accumulation of proline under that level of stress condition showed the same trend i.e., by increasing osmotic stress at a very high level, the level of proline decreased substantially when the germination % reduced to its lowest rate. By analyzing the possible contribution of the degradation processes of macromolecules to the accumulation of soluble sugars and proline, both polysaccharides and proteins seemed to contribute equally to support the seedlings under stress conditions. As soluble sugars showed a gradual and consistent reduction with increasing salt stress around the seeds, so as to provide the carbon skeleton to build other active soluble solutes in the seedling tissues. One of these solutes was proline; and it accumulates to maintain the machinery of plant tissues, in addition to its contribution to the osmotic adjustment (Table 5) [48,61-63].
| NaCl (dSm-1) | Polysaccharides (mg g-1DW) | % of the degradation out of the original content | Protein (mg g-1 FW) | % of the degradation out of the original content | Proline (µgg-1 FW) |
|---|---|---|---|---|---|
| Control | 30.8 | 58 | 4.9 | 63 | 80 |
| 20 | 52.0 | 29 | 7.0 | 47 | 256 |
| 40 | 69.9 | 5 | 12.6 | 5 | 348 |
Table 5: The effect of salt stress on some biochemical parameters in the germinated seeds of the Jordanian barley (Hordeum vulgare L.) cultivar, Rum [49]. The content of the untreated seeds: Polysaccharides=73.7 mgg-1DW, protein=13.3 mgg-1FW, and proline=11.0 µgg-1 FW.
These data clearly show that proline accumulates substantially in the seedlings, despite the low degradation rate of macromolecules in the seedling tissues, as osmotic stress increased around the seeds. From these results, the following explanations can be drawn: (1) At low osmotic stress, most of the reserves in the seeds are used to build new cells which are expressed in terms of high germination percentages and vigorous growth, (2) by increasing the osmotic stress around the seeds, the degradation of macromolecules declines depending on the plant species and environmental conditions of the experiments. However, the soluble solutes that resulted from the degradation of stored macromolecules are directed to the building of new cells and to maintaining the machinery of cells under stress. At this stage, there is a decrease in the growth of seedlings while compatible organic solutes accumulate significantly, and (3) at high osmotic stress, the degradation processes of macromolecules are inhibited substantially and are expressed in terms of low germination percentages and little growth of radicles. However, most of the soluble solutes that are formed as a result of the degradation of macromolecules are directed to maintaining the osmotic balance between plant tissues and the growth medium, as well as maintaining the active sites around the cells [48,49,58,64] (Figure 1; Table 6). On the other hand, when the proline concentration in the germinated seeds is compared with the nongerminated seeds under high salt levels (30 and 40 dSm-1) as shown in Table 7, it is clear that once the seeds are suppressed by high osmotic stress, the enzyme systems of proline biosynthesis are inhibited. Also, all metabolic activities [49], including the reduction in hydrolysis of macromolecules (carbohydrates, proteins and lipids) in the embryonic axis result to diminution of germination and seedling growth [65,66].

Figure 1: During germination: metabolites can be used mainly either for growth (A) and / or for osmotic adjustment (B) under dierent osmotic stress conditions.
| NaCl (dSm-1) | Germination (%) | Radicle length (mm/radicle) | Polysaccharide (mg glucose g-1DW) | Protein (mg protein g-1FW) | Proline (µgg-1 FW) |
|---|---|---|---|---|---|
| Control | 100 | 88 | 18 | 4.9 | 72 |
| 10 | 98 | 59 | 32 | 6.79 | 121 |
| 20 | 95 | 25 | 43 | 6.95 | 248 |
| 30 | 58 | 4 | 68 | 7.12 | 365 (70)* |
| 40 | 15 | ≈ 2 | 72 | 12.58 | 348 (63)* |
Table 6: Changes in proline and macromolecules in the germinated seeds of the Jordanian barley (Hordeum vulgare L.) cultivar, Rum, in response to salt stress [49]. *Proline content of the non-germinated seeds. The content of barley seeds before treatments: Polysaccharides=73.7 mg glucose g-1DW, protein=13.3 mgg-1FW, and proline=11.0 µgg-1 FW.
| NaCl (dSm-1) | Germinated seeds | Non-germinated seeds | ||||
|---|---|---|---|---|---|---|
| Temperature° C | ||||||
| Control | 15 | 20 | 25 | 15 | 20 | 25 |
| 30 | 393 | 414 | 365 | 83 | 72 | 70 |
| 40 | 358 | 307 | 348 | 65 | 60 | 63 |
Table 7: Proline concentration (µgg-1 FW) in the germinated and non-germinated seeds of the Jordanian barley (Hordeum vulgare L.) cultivar, Rum, at three temperatures and high salt concentrations [49]. The concentration of proline in the un-treated seeds was 11.0 µgg-1 FW.
Growth and development stages: Subsequent plant growth, after the germination stage, relies on photosynthesis and other metabolic activities to provide all the metabolites necessary for maintaining the machinery of plant tissues. Osmotic stresses (salinity and drought) are known to adversely affect many physiological processes and biochemical reactions in crops and wild plant life as well, such effects involve morphogenesis and many metabolic pathways [67]. The reduction of total leaf area is a common response to such abiotic stresses and are attributed to the reduction in the area of individual leaves [47,68]. Both growth processes, cell division and cell expansion, show considerable inhibition, and have a great influence on the number and volume of cells [69], thereby reducing the size and area of leaves. In fact, some reviews [70] have discussed the mechanism of the impact of these factors on growth variables (growth rate and growth duration), growth processes (cell division and cell expansion) and summarized the physiological and biochemical activities involved in these variables and processes: (1) reduction in nucleic acid synthesis, (2) inhibition of the activity of both auxin and cytokinin, and (3) changes in the metabolic activities of the cell wall. Recent studies have concluded that all these activities involve changes in the formation of soluble sugars by one way or another [8]. For example, monosaccharides such as glucose was found to accumulate in the leaves of fenugreek plants grown in a soil with low matric potential, such accumulation might have contributed to the process of osmotic adjustment. Table 8 shows the content of some important parameters responsible for maintaining the osmoregulation and the machinery in the shoot system of fenugreek plants under water stress conditions. A significant accumulation of monosaccharides and proline was found in the shoot system which was almost linear with the reduction in relative water content (RWC), and that accumulation was gradual and consistent as the matric potential of soil decreased, reaching its highest contents at –0.4 MPa of soil matric potential. Such high solute contents, at very low soil water potential, could be caused by physiological drought stress [71], which plants attempt to overcome by the accumulation of organic compounds, thereby causing a decrease in the solute potential and keeps a positive balance of water absorption and translocation [69,72]. Some evidences have been given that the accumulation of these compatible solutes is correlated with the xerophytic features and dehydration tolerance of native plants [21,73].
| Soil matric potential (MPa) | DRV | Sucrose | Proline | RWC |
|---|---|---|---|---|
| Control | 7.95 | 21.4 | 1.21 | 91 |
| -0.1 | 8.75 | 21.6 | 1.24 | 90 |
| -0.2 | 8.77 | 22.3 | 1.4 | 92 |
| -0.3 | 12.36 | 22.3 | 1.83 | 89 |
| -0.4 | 12.49 | 17.6 | 12.73 | 83 |
Table 8: Soluble sugars (mgg-1DW), proline (µmol g-1 FW), and RWC (%) in the shoot system of fenugreek plants exposed to different soil matric potentials [21].
On the other hand, sucrose showed almost stable content under different levels of stress conditions which could explain its role in turgor maintenance. These results were confirmed in a study on the callus of two wheat genotypes under osmotic stress conditions, total soluble carbohydrates and proline accumulate to accelerate water uptake and maintain growth [74]. Results have been obtained from other plants [75] like rice (Oryza sativa L.), which showed varietal differences by comparing salt resistant cultivars with sensitive ones. The starch content decreased more in the roots of sensitive cultivars, but no changes were found in the shoot system, as the starch phosphorylase activity decreased in all cultivars. On the other hand, the content of soluble sugars (reducing and non-reducing) along with sucrose phosphate synthase increased more in the salt sensitive cultivars, while the activity of acid invertase decreased in the shoots of salt tolerant cultivars and increased in the salt sensitive ones. Almost similar results were obtained later from another study [76] on other rice cultivars which differ in their salinity response. It was found that soluble sugars accumulate in salt sensitive cultivars whereas tolerant ones had almost the same content of soluble sugars under all salinity levels. Others [31], found that there was a substantial decrease in the activity of α-amylase and protease under salt stress in some cotton cultivars. However, these parameters clearly distinguished the cultivars studied; for example, the cultivar K-115 showed better performance than other cotton cultivars like NIAB-Karishma and NIAB-86. Also, the cultivar K-115 had the capacity to mobilize a significant content of sugars, total free amino acids and the reserved protein during germination and early seedling growth stages. Moreover, other studies [32] have reported that more soluble sugars and less starch content in the cotyledons of okra (Abelmoschus esculentus L.) along with lower amylase activity as NaCl increased around the germinating seeds. This study has concluded that such disturbances of germination were accompanied with the increase in Na+ of the embryonic axis. In contrast, the early findings of [34] have concluded that the α-amylase activity in barley leaves under water stress increased as a result of the enhancement of its synthesis and the induction of α-amylase gene and an increase in the level of the corresponding mRNA. Such findings have been confirmed by other authors [35], in some crops like Glycine max , salt stress reduced invertase activity and increased amylase activity. This study concluded that the salt stress that caused a restriction of sucrose utilization in the leaves could partly be a result of the ionic effect on degradation, while no role was seen from the starch.
Soluble Sugars in Native Plants: Native plants at the Arabian Gulf region in general and in the State of Qatar in particular, include mainly xerophytes and halophytes, and the checklist of these plants has been reviewed several times, and the last report [77] about the wild plants in Qatar listed about 400 plant species [78]. These plants belong to various families with different morphological and physiological characteristics [79,80]. In fact, these plants have adopted different metabolic mechanisms which resulted from different abilities to accumulate soluble sugars under their natural harsh environments (Table 9).
| Plant Species | Main characteristics | Soluble sugars (mgg-1 DW)* | References** |
|---|---|---|---|
| Aeluropus lagopoides | Gramineae, Grass, Xerophyte, Not succulent, | 1.5-1.7 | [81] |
| Anabasis setifera | Chenopodiaceae, Low undershrub, Xerophyte, Succulent leaves, | 1.6-2.0 | [79,82] |
| Arthrocnemum macrostachyum | Chenopodiaceae, Low circular undershrub, Halophyte, Succulent shoots, | 4.0-14.1 | [83] |
| Avicennia marina | Avicenniaceae, Mangrove tree, Halophyte, Not succulent, | 2.2-6.0 | [84] |
| Halocnemum strobilaceum | Chenopodiaceae, Low circular undershrub, Halophyte, Succulent shoots, | 1.8-2.5 | [85] |
| Halopeplis perfoliata | Chenopodiaceae, Undershrub, Halophyte, Succulent shoots, | 1.9-4.6 | [86] |
| Heliotropium bacciferum | Boraginaceae, Undershrub, Xerophyte, Not succulent, | 10.6-16.6 | [87] |
| Limonium axillare | Plumbaginaceae, Undershrub, Halophyte, Succulent leaves, | 4.7-8.0 | [88] |
| Ochradenus baccatus | Resedaceae, Shrub, Xerophyte, not succulent, | 2.9-3.5 | [89] |
| Pulicaria crispa | Compositae, Suffrutescent, Xerophyte, Not succulent, | 8.8-9.9 | [79,86] |
| Pulicaria gnaphalodes | Compositae, Suffrutescent, Xerophyte, Not succulent, | 6.3-7.2 | [79,86] |
| Salsola imbricate | Chenopodiaceae, Undershrub, Halophyte, Succulent leaves, | 1.6-3.1 | [79,86] |
| Seidlitzia rosmarinus | Chenopodiaceae, Undershrub, Halophyte, Succulent shoots, | 2.8-4.7 | [85] |
| Sporopolus spicatus | Chenopodiaceae, Grass, Xerophyte, Not succulent, | 3.3-3.4 | [90,91] |
| Suaeda aegyptiaca | Chenopodiaceae, Undershrub, Halophyte/Xerophyte, Succulent shoots, | 3.9-4.3 | [92] |
| Suaeda vermiculata | Chenopodiaceae, Undershrub, Halophyte, Succulent shoots, | 6.7-19.8 | [92] |
| Tetraena qatarensis | Zygophyllaceaae, Undershrub, Halophyte/Xerophyte, Succulent shoots, | 2.9-4.3 | [93] |
Table 9: List of some native plants in Qatar and the range of concentrations of soluble sugars in the shoot system [79]. *Range of 10 readings, trehalose not measured, **These references did not show that trehalose accumulates in these plants.
However, the range of these concentrations showed clearly that native plants in this region accumulate less soluble sugars as compared to the crop plants mentioned above; like wheat, barley and fenugreek, which might give the impression that native plants accumulate other compatible solutes like proline and glycinebetaine, etc., to cope with these habitats. The main soluble sugars found in these plants are monosaccharides and sucrose [79]. However, from the best of our knowledge, there is no report about the accumulation of other soluble sugars like trehalose in the tissues of these plants. The literature reviews conducted all over the world did not show any confirmation that trehalose accumulates in the native plants of Qatar. However, during the last two decades, studies have shown that the soluble sugars content were higher in Suaeda vermiculata and Heliotropium bacciferum , followed by Arthrocnemum macrostachyum, Pulicaria crispa , Pulicaria gnaphalodes , Limonium axillare , and the least concentrations were found in Sporopolus spicatus, Tetraena qatarensis , Ochradenus baccatus , Anabasis setifera and Aeluropus lagopoides [79].
The above studies have concluded that there is no consistency in the response of the carbohydrate metabolism, and the reasons behind such a conclusion have been summarized recently [11] as follows: disturbances in the balance between photosynthesis which produces sugars with the processes of their utilization like respiration and other metabolic activities, variation in the stomatal resistance between plants under osmotic stress, which affects the accumulation through CO2 fixation in photosynthesis, disturbance in the metabolism of carbohydrates which includes increasing and/or decreasing enzyme activities like amylase, invertase (β- fructosidase), sucrose phosphate synthase and sucrose synthase [21].
Trehalose is a disaccharide composed of two molecules of glucose units joined by a 1-1 alpha glycosidic bond, giving it the name of α-Dglucopyranosyl-( 1→1)-α-D-glucopyranoside, a molecule of it is a nonreducing sugar (Figure 2).

Figure 2: The molecule of trehalose.
Some reviews [96] reported that this compound was discovered in 1832 by Wiggers in the ergot of rye, and after many years, it was isolated from mushrooms by Mitscherlich in 1858, who called it mycose. Then, after a long period of time, this sugar was recognized to have many roles, activities, and functions in various taxa of living organisms [97]. Its presence in these groups, from bacteria, fungi, protista, animals, and plants, is to play various functions in the metabolic homeostasis, as compatible solute and stress metabolite to compete with osmotic stress, as osmo-protectant and/or cryoprotectant agent offering tolerance mechanism and preservation of the membrane systems against various abiotic stress conditions [12,98].
It also functions as a protection agent of cell organelles and proteins, and it is useful for other roles including the structural components of insect and shrimp shells, and cell wall glycolipids of some bacteria, as a component in some food stuffs, and as an external carbon source [98-101].
Several reports have indicated that the presence of trehalose in these living organisms, is to serve as a signaling molecule to control some metabolic activities at various growth stages [102], as well as other roles like carbon storage and the scavenging of reactive oxygen species [67]. Moreover, trehalose can be considered as an important part of the defense systems of living organisms that involves intracellular accumulation of protective compounds that shield macromolecules and plasma-membranes from a damage against these stresses [11]. As a compatible solute, trehalose accumulates substantially in microorganisms like bacteria and fungi. However, in most vascular plant species, this is not the case. They do not accumulate significant amounts of this sugar with the rare exception of some tolerant plant species to desiccation stress [4,103,104], for example, some ferns and the angiosperm Myrothamnus flabellifolia [90]. Some articles [105] have indicated that genes from Arabidopsis are able to complement yeast strains deficient in trehalose metabolism, and such findings could be utilized in the development of strategies and applications for plant biotechnology under osmotic stress. Also, some salt tolerant glycophytes might accumulate trehalose which have been considered as a good model for studying the molecular approach to improving the resistance of crops under extreme environmental conditions [102]. Regarding the presence of trehalose in the native plants of Qatar, so far, no serious attempts have been made to measure the amount of trehalose in these plants. Moreover, a huge number of articles around the world have shown that these plants accumulate soluble sugars at some degree or another, but there is no clear indication that trehalose is among the soluble sugars found in these plants.
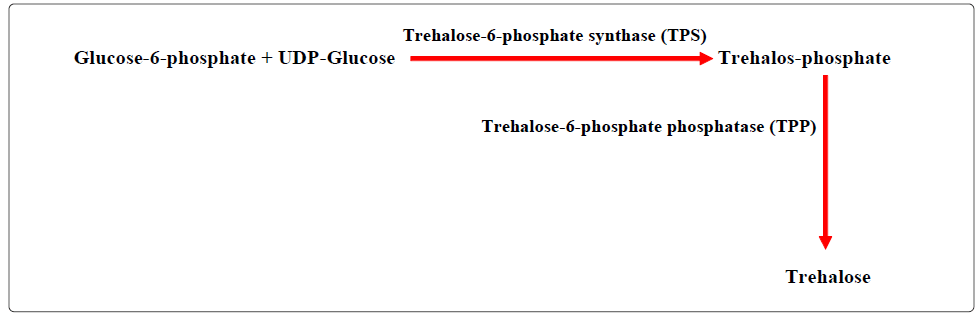
Five metabolic pathways have been reported for the biosynthesis of trehalose in various living groups, one of these pathways is common and widely found in bacteria, fungi, invertebrates and plants [106,107]:
From the best of our knowledge, there are no reports about the presence of trehalose in the native plants of the Arabian Gulf region. However, at least one report [108] has revealed that a compound similar to α-α-trehalose (α-D=glc-1-1-α-D-glc) is found in the Polysiphonia * species.
*Polysiphonia is a genus of filamentous red algae with about 200 species worldwide.
Trehalose in Biological Soil Crusts (BSC)
BSC are comprised of various living groups and communities of bacteria, fungi, lichens, cyanobacteria and mosses, which are essential parts of the soil biota playing vital roles and activities in improving nutritional status and stabilizing the soil surface by preventing erosion, especially in the dryland and Sabkhas. Many microorganisms adjacent to and/or associated with these plants might build trehalose, in order to conduct many important roles to support the growth of these plants.
Recent reports [11,109] have confirmed that these microbes could adopt many mechanisms to alleviate the harsh abiotic stresses facing plants in general and crops in particular. These methods and mechanisms include the following: (a) biofilm formation, (b) polymers production, (c) chemotaxis, (d) phytohormone production, (e) nitrogen fixation, (f) phosphate solubilization, (g) production of phytohormone-degrading enzymes and (h) osmolytes biosynthesis. However, the last mechanism has been very interesting in providing a great deal of support to native plants and crops, and some important articles and reviews, based on serious experimental works showed clearly that microorganisms adjacent to and/or associated with many plants might produce solutes to confer some resistance traits under extreme environmental conditions [12,110,111]. Therefore, from the above records, serious and comprehensive research plans should focus on the ability of native plants to synthesize trehalose. These reports have come to a conclusion that a new approach has emerged during the last decade that microorganisms might provide some compatible solutes to native plants, enabling them to resist extreme environmental conditions. Moreover, the halotolerant and halo-thermophilic bacteria found in Qatari soils could be used in many biotechnological applications, in addition to other natural methods of Horizontal Gene Transfer (HGT), to provide trehalose sugar to crop plants, thereby conferring some important resistance traits against extreme environmental conditions.
Bacteria: Bacteria at the coastlines and Sabkhas of the Arabian Gulf region have a remarkable capacity for adaptation to various harsh stresses such as drought, salinity and high temperatures [109,112]. Considering the bacteria species in Qatar (Table 10), the international records indicated that many of them are efficient in accumulating trehalose sugar, and these species include: Escherichia coli (E. coli ), Halorhabdus sp., Natronobacterium sp., Pseudomonas spp., Staphylococcus spp., and Streptomyces spp. Other species might be able to accumulate trehalose, especially those that proved positive in trehalose fermentation and biochemical tests; Achromobacter xylosoxidans, Aeromonas hydrophila, Bacillus amyloliquifaciens, Bacillus cereus, Enterobacter cloacae, Klebsiella pneumoniae, are examples, bearing in mind that many other bacteria species might not have been tested yet.
| Bacteria species | Trehalose | References |
|---|---|---|
| Achromobacter xylosoxidans | Trehalose Fermentation Test*:+ | [116,117], Unpublished observations |
| Aeromonas hydrophila | Trehalose Biochemical Test*:+ | [118,119], Unpublished observations |
| Bacillus amyloliquifaciens | Trehalose biochemical test:+ | [120] |
| Bacillus cereus | Trehalose Biochemical Test: + N. B. https://biochemicaltest.com/biochemical-test-of-bacillus-cereus/; | [11,109,112,119,121] |
| Bacillus megaterium | Trehalose presence:? | [119,121], Unpublished observation |
| Bacillus thuringiensis | Trehalose presence:? | [11,109,112,119,121] |
| Burkholderia spp. | Trehalose presence:? | [122,123], Unpublished observations |
| Chromobacterium violaceum | Trehalose presence:? https://microbewiki.kenyon.edu/index.php/Chromobacterium_violaceum; N. B. Metabolise trehalose |
[119], Unpublished observations |
| Enterobacter cloacae | Trehalose biochemical test:+ | https://microbiologyinfo.com/wp-content/uploads/2017/05/Biochemical-Test-and-Identification-of-Enterobacter-cloacae.pdf |
| Escherichia coli | Trehalose presence :+ | [124] |
| Haloarcula spp. | Trehalose presence :? | [125] |
| Halobacterium spp. | Trehalose presence :? | [126] |
| Haloquadratum sp. | Trehalose presence:? | [12] |
| Halorhabdus sp. | Trehalose presence: + | [127] |
| Klebsiella pneumoniae | Trehalose Biochemical Test:+ | https://microbiologyinfo.com/biochemical-test-and-identification-of-klebsiella-pneumoniae/ |
| Kocuria kristinae | Trehalose presence:? | [128] |
| Natronobacterium sp. | Trehalose presence:+ | [129,130] |
| Sphingomonas paucimobilis | Trehalose presence:? | [131] |
| Pseudomonas spp. | Trehalose presence:+ | [100] |
| Staphylococcus spp. | Trehalose presence:+ | [113], http://delrio.dcccd.edu/jreynolds/microbiology/2421/lab_manual/StaphID.pdf |
| Salinibacter ruber | Trehalose presence:? | [132] |
| Streptomyces spp. | Trehalose presence:+ | [114] |
Table 10: The bacteria species found in Qatar are adjacent and/or associated with native plants and might accumulate Trehalose. *The purpose of these tests is to see if the microbe can ferment trehalose as a carbon source.
Early studies of the eighties of the last century had concluded some important facts about the trehalose accumulation in some bacteria species like Staphylococcus spp. and Streptomyces spp. Stevens, Jones [113] succeeded in differentiating two Staphylococcus spp. from other Staphylococci, using a plate medium containing trehalose. Other studies [114] found that Streptomyces antibioticus accumulates trehalose during various stages of its growth, it was detected during all phases of colony development and in most of its aerial hyphae and spores. They suggested some important roles and activities that this group can play like: carbohydrate storage, energy source and protecting the structural and functional integrity under desiccation stress. More attempts [115] have succeeded to understand the mechanism of trehalose metabolism in dormant spores and during spore germination in Streptomyces griseus. Trehalase enzyme was inactive in dormant spores; trehalose accumulates, on the other hand, trehalose was hydrolyzed upon initiation of spore germination to conduct various roles. However, the actual mechanism by which trehalose metabolism is regulated in S. griseus spores was not resolved at that time.
In their studies on a range of archaebacteria, some authors [129] concluded that trehalose was identified in a number of halophilic archaebacteria including Natronobacterium spp., thermophilic and sulphur-dependent archaebacteria and methanogenic archaebacteria, while others [130] have confirmed the presence of some trehalose derivatives like sulfotrehalose in Natronobacterium sp. and Natronococcus occultus.
During the last two decades, some other important roles have been recognized in other bacteria species. For example, E. coli utilizes a variety of organic compounds, and trehalose is among these solutes. Trehalose serves as a primary protective osmolyte [12]. It has been found that genes encoding trehalose biosynthesis are widely distributed in nature and have been extensively studied in E. coli (Bacteria) and Saccharomyces spp. (Yeast). In E. coli , the trehalose biosynthetic operon (otsBA) is induced by severe osmotic stress, extreme heat, cold conditions as well as desiccation stress [124].
These authors have concluded that the overproduction of trehalose could be a useful trait for future modern genetic approach, to improve the resistance of living organisms including crop plants against those environmental conditions [96,133]. Halorhabdus sp. is found in some salterns and Sabkhas around Qatar, and some species belonging to this genus possess trehalose synthase (TreS); this enzyme catalyzes the reaction:
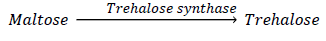
Trehalose can be utilized in some important functions as a compatible solute as well as a storage compound [127]. Other studies [100] reported that Pseudomonas syringae accumulates trehalose and other organic compatible solutes under water limiting conditions, they evaluated the contribution of this sugar in epiphytic fitness with tomato plants. Moreover, some researchers [134] have proposed that the trehalose produced by some microbes like the plant growthpromoting rhizobacteria (PGPR); e.g., Pseudomonas putida , might offer some kind of protection to plants including crops like tomato and pepper against drought and possibly other abiotic stresses; and surely such achievements have been very useful and interesting. In a recent report [109], E. coli was predominantly found in untreated ponds around Doha city; also, other bacteria species were found and should be investigated [119]. Thus, E. coli could be a good choice as an experimental material to conduct comprehensive research activities to consider the possibility of trehalose production by this microorganism, thereby supporting the growth of plants and possibly crops, under the natural environmental conditions. Therefore, any future research plan should cover the testing of trehalose in native plants and the associated microorganisms to gain more understanding of the mechanisms against environmental stresses in the wild life of this region.
Regarding the presence of soluble sugars in cyanobacteria, some studies [135] have found some soluble sugars like sucrose, glucosylglycerol and trehalose in many strains of cyanobacteria under osmotic stress conditions. They could not find any clue of differences between those strains living in freshwater and saline habitats, in terms of these compatible solutes. Such findings were confirmed by previous studies [136] on some drought-resistant cyanobacteria which showed different abilities to accumulate sucrose and trehalose, under drought stress. Several studies have discussed the accumulation of trehalose in cyanobacteria [137], Nostoc punctiforme accumulates trehalose when exposed to salt and water stresses, and no trehalose was found in fully hydrated cells. The genes responsible for trehalose biosynthesis were induced under stress conditions, and, on the other hand, trehalase activity was strongly inhibited in the presence of 10 mM NaCl, meanwhile trehalose synthesis remained active in the presence of salt. Therefore, this study concluded that under these conditions, the accumulation of trehalose is regulated by two factors: (a) the rate of trehalose synthesis exceeds its degradation, and (b) the suppression and control of trehalase activity. Other authors [138] have reported that under osmotic stress; sucrose, trehalose, glucosylglycerol, proline and glycinebetaine are the main compatible solutes found in cyanobacteria playing various roles. The genetic and molecular basics of the biosynthesis of these compatible solutes can be utilized for various useful roles as follows: 1) to increase the resistance against various stresses for various biotechnological purposes and 2) to characterize the adaptation of these living organisms to certain habitats and ecological niches.
Although few studies have been conducted about the cyanobacteria in Qatar, some articles have reported some genera and species like: Anabaena and Nostoc found adjacent to Sabkhas [112], Chroococcidiopsi s, Aphanothece, Pleurocapsa, Oscillatoria, Lyngbya, Leptolyngbya, Phormidium and Scytonema found among hypolithic microbial communities [139], and Microcoleus and possibly others found underneath the rocks in many locations around the country [109]. These species and genera could play significant biological roles to support wildlife as well as provide experimental material for modern biotechnological studies, to improve the resistance of plants against extreme environmental stresses [140].
Fungi: They play significant roles and activities in the energy equilibrium of the ecosystem, and in the soil, they decompose and break many dead organisms into molecules, and at the end these molecules become available and usable by other living organisms including native plants and crops. Few studies have been published about the role of fungi in the Qatari ecosystem, and Qatari soil fungi were first reported by Moubasher and co-workers [141,142], 53 genera and 142 species and 8 varieties were found in most habitats and locations around Qatar. The most recognized genera in the Qatari soil were: Aspergillus (23 species and 5 varieties), Penicillium (16 species), Fusarium (8 species and one variety), Cladosporium (5 species), Stachybotrys (one species with one variety), Acremonium (5 species), Botryotrichum (2 species), Alternaria (5 species), and Microascus (4 species), with rare occurrence of other species. Moreover, some macrofungi were listed by [143] and included: Tulostoma album, Agaricus sp., Leococoprinus sp., Tirmania nivea (Zbaidi desert truffle), and Trefezia claveryi (Khlasi desert truffle). All these groups, genera and species are surely well adapted to the environmental condition at the Arabian Gulf region. Some of the above-mentioned fungal species were isolated from the Qatari soil at a temperature as high as 45°C. These species included: Aspergillus fumigatus, Malbranchea pulchella, Absidia corymbifera, Myriococcum albomyces, Thermomyces lanuginosus, Humicola insolens, Thermoascus aurantiacus and possibly others. Moreover, 14 genera; isolated from different habitats around Qatar, were added to the above list belonging to Ascomycota and Basidiomycota [144]. These included: Agaricus sp., Tulostoma album , Volvariella volvacea , Terfezia claveryi, Tirmania nivea, Ganoderma lucidum, Phaeangium lefebvrei, Montagnea arenaria, and others. Regarding the research activities, no serious works have been done about the eco-physiological aspects of these fungi and any biotechnological applications in Qatar or in any other Arabian Gulf State. Thus, any future plan should concentrate on these fungi and tackling all the above issues and the roles that these species might play in the ecosystem.
Starting with the main mechanisms adopted by BSC to deal with the natural environmental conditions in Qatar, compatible solutes including soluble sugars have been recognized as the main solutes accumulated in these fungi, to conduct various roles in soil biota [109,112], bearing in mind that the trehalose sugar could be among these solutes [145,146]. Table 11 shows the most common species of fungi in the State of Qatar that proved active in the trehalose metabolism. The reports on trehalose accumulation in fungi started a very long time ago, as it was first documented in 1832 in the fungus Claviceps purpurea , this fungus grows on rye and other related plants and produces alkaloid compounds causing ergotism in humans and cattle [97]. Going back to the sixties of the last century, trehalose was first reported by some authors [147,161,162], who published works about the presence of this sugar in the same species of fungi found in Qatar like mushrooms (Agaricus bisporus ) and Myrothecium verrucaria . Gas chromatography technique was first used to determine trehalose in the basidiospores of Agaricus bisporus at different developmental stages which showed that trehalose ranged between 0.03–0.3% of the fresh weight. However, Agaricus sp. identified in Qatar is commonly found in garden soils; and there is need to investigate trehalose metabolism and its role under the current environmental conditions. Most of the metabolic pathways of trehalose biosynthesis and degradation have been well determined and documented, in fact, a huge number of articles have discussed the changes in trehalose in many species of fungi across the world, especially those found in Qatar. Other authors [157] have studied the main characteristics of trehalases in the thermophilic fungus Humicola grisea var. thermoidea ; both the cytosolic and the conidial enzymes were compared in these reports [158].
| Fungi: Groups, Genera and Species | Presence of Trehalose | References |
|---|---|---|
| Agaricus sp. | ? | [147] |
| Aspergillus nidulans | + | [148,149] |
| Aspergillus niger | + | [150] |
| Aspergillus fumigatus | + | [151] |
| Aspergillus fischeri | + | [152] |
| Aspergillus flavus | ? | [149] |
| Alternaria alternata | + | [153] |
| Arthrobotrys oligospora | + | [154] |
| Beauveria alba | ? | [155] |
| Fusarium oxysporum | + | [156] |
| Humicola grisea | + | [156-158] |
| Malbranchea pulchella | + | [159] |
| Metarhizium anisopliae | ? | [160] |
| Mucor spp. | ? | [156] |
| Myrothecium verrucaria | + | [161,162] |
| Paecilomyces spp. | ? | [163] |
| Rhizoctonia solani | + | [164] |
| Rhizopus spp. | ? | [165] |
| Thermoascus spp. | ? | https://www.sciencedirect.com/topics/immunology-and-microbiology/ascospore |
| Trichoderma sp. | ? | [166] |
| Mushrooms | + | https://www.sciencedirect.com/topics/medicine-and-dentistry/trehalose |
| Yeast | + | https://www.sciencedirect.com/topics/medicine-and-dentistry/trehalose |
Table 11: The common species of fungi found in most habitats and locations around Qatar that might accumulate Trehalose.
Others [167] have suggested that the presence of glucose in yeast is required for many regulatory phenomena, one of which is the glucosesensing mechanism for the trehalose-6-phosphate synthase catalytic subunit of the trehalose synthase complex, which has been widely found in yeast and possibly in other living organisms. Trehalose sugar is common in various parts and structures of fungi like the vegetative cells, spores and fruiting bodies.
Also, its biosynthesis is induced under extreme environmental conditions to play various roles and activities [102]. Trichoderma harzianum (Ascomycota) is a good example of a fungus that accumulates trehalose substantially in the ascospores, under these conditions [166]. Many other reports [152] have shown that the ascospores of many Ascomycota genera like Neosartorya, Byssochlamys and Talaromyces contain substantial amounts of trehalose. Such high sugar content could help these ascospores to survive high temperatures and possibly desiccation stress. More evidences have been introduced that two classes of trehalase in some fungi like yeasts and filamentous fungi such as Fusarium oxysporum and Humicola grisea play different roles [156]. These roles are independent and specific, depending on the extracellular trehalose or cytosolic origin for conducting some activities inside fungal cells. Pereira et al. [159] found that Malbranchea pulchella var. sulfurea produced a significant amount of extracellular trehalase activity when grown for long periods on starch, maltose or glucose as the main carbon source, this enzyme utilizes trehalose as a carbon source. Also, trehalose acts as a stress protectant in Rhizopus oryzae , in response to various environmental stresses like heat, osmotic stress, nitrogen starvation and pH changes [165]. Molecular approach started in the nineties of the last century with Wolschek and Kubicek [150], who found two genes encoding trehalose-6-phosphate synthase in Aspergillus niger (tysA and tysB) which have 64-70% identity at the amino acid level with the GGS1/TPS1 genes of some yeast species like Saccharomyces cerevisiae and Kluyveromyces lactis and the TPS1 gene of Schizosaccharomyces pombe. Some researchers [148] have found a regulator gene (VosA) in Aspergillus nidulans that couples the completion of sporogenesis, i.e., negative-feedback regulation of sporogenesis, and accumulation of trehalose in spores. Trehalose, on the other hand, as a storage compound may serve as a substitute for mannitol in the spore germination of Alternaria alternata while growing on the host tobacco plant [153]. Several studies have been conducted to investigate the relationship between Aspergillus fumigatus virulence and the trehalose biosynthesis pathway. Others [151] have confirmed that this pathway is a promising antifungal drug target and plays a critical role in regulating key aspects of the biology of this fungus including glycolytic flux, cell wall integrity and virulence. Also, more innovative and contemporary studies have been conducted to improve the virulence of the locust fungus Metarhizium acridum . This fungus was modified genetically to overexpress an endogenous hydrolase of trehalose, which is the main carbon source in insect hemolymph [160]. During the last decade, more investigations were conducted about the genes responsible for conidiation, sclerotia formation and other activities in some Aspergilli . These studies concluded that these species have differential gene expressions in the promotion and reduction of many reproductive activities [149]. Moreover, recent works [155] examined the functions of two trehalose-6-phosphate synthase (TPS) genes (tpsA and tpsB) in Beauveria bassiana, these genes play complementary roles in sustaining trehalose synthesis, conidiation capacity, conidial quality, multiple stress tolerance, and virulence. This study highlighted the fungal adaptation to the environment and host. Recent works on the fungus Rhizoctonia solani [164]; the causal agent of rice sheath blight which causes severe economic losses in the rice-growing regions across the world, have found that sclerotia represents an important part of the life cycle of this fungus. This fungus has six genes of differential expressions affecting sclerotia development. This study showed that ROS produced under some extreme environmental stresses (example: salt stress), might have significant accelerating influence on the development of sclerotia. Trehalose, on the other hand, has a counter effect on the formation of sclerotia by this fungus. This has been proven by the exogenous application of trehalose which has a significant inhibitory effect on the activities of ROS enzymes and it resulted to a reduction in sclerotial dry weight. Finally, trehalose has been recognized as the main compatible solute for many fungal activities: (1) found in reproductive bodies; sexual and asexual, (2) essential for survival and infection of fungal spores.
The most common lichens that have been recorded in Qatar include: Acarospora sulphurata, Buellia subalbula, Caloplaca aurantia, Caloplaca brouardii, Caloplaca irrubescens, Collema tenax, Diploschistes albescens, Gloeoheppia turgida, Lecidea sp., Lecidella euphorea, Ramalina farinacea, Ramalina maciformis, Verrucaria sp. and Xanthoria parietina [168]. None of these lichens species have been proven to accumulate or metabolize trehalose.
The genetic variability among plants has been considered as experimental material for modern biotechnological studies to determine the structural, physiological and biochemical features that are consistently associated with the resistance against various biotic and abiotic factors. During the last two decades, significant achievements have been made to improve the resistance of crop plants to abiotic stresses, these efforts concentrated on increasing the concentration of compatible solutes, especially soluble sugars like trehalose, in plant tissues [169]. Genetic manipulations have been introduced and adopted [170,171] using conventional breeding methods (selection and hybridization), and modern biotechnological techniques (these include: tissue culture techniques, somatic cell genetics, and recombinant DNA techniques) to develop transgenic plants [78,172], and recently using resistant genes from some microbes [173]. Transgenic plants are capable of synthesizing solutes like proline, glycinebetaine and possibly other compatible solutes like trehalose (Figure 3), to provide a protection mechanism for alleviating various types of stresses [11,109,174,175].

Figure 3: Possible methods and mechanisms can be adopted by microorganisms (bacteria and yeast) to produce trehalose and increase abiotic stress resistance in plants. This figure was modified from figure 20 of [109].
In fact, some serious attempts have been made to transform some crops like rice with a trehalose-6-phosphate synthase/phosphatase (TPSP) fusion gene that includes the coding regions of the E. coli otsA and otsB genes (encoding TPS and TPP, respectively) [176]. These efforts have resulted in the regulated overexpression of trehalose biosynthetic genes in rice which has considerable potential for improving abiotic stress tolerance [4]. These outcomes could be exploited to improve the resistance of other major crops like barley, wheat, and maize against severe environmental conditions. More researches should be conducted on native plants across the Arabian Gulf region including Iraq, Iran and Yemen to evaluate the possibility of the presence of this compound in native plants, and to adopt modern biological techniques to improve crops capable of accumulating trehalose. The cost of these methods is still very high and requires efforts and time to improve the resistance of crop plants under extreme environmental conditions.
A third environment-friendly biological approach has emerged in the last decade, microorganisms that colonize various plant species or those even adjacent to plants can provide some desirable traits using one or more methods. These microorganisms are capable of synthesizing compatible solutes which are then released to the environment in support of the host plants. Some microorganisms may introduce genes to the environment; by the natural method of HGT to introduce genetic materials to other living organisms including plants. The HGT method has been considered as a successful approach compared to the traditional genetic modification techniques [177]. This approach has been supported by many incidents which have been widely observed across the domains of life [178]; moreover, many genes were found in eukaryotes to have microbial origin [179]. This approach appears to be a good alternative to both conventional methods and biotechnological techniques and has been considered as a natural and cost-effective approach.
Inevitable conclusions
Scientists study tiny creatures to improve other living organisms by introducing unique traits that can produce desirable products to solve problems mankind is facing in agriculture, health, economy, and industry. Increasing plant production, qualitatively and quantitatively, has been the main objective of scientists and decision makers. The essence of this is to counteract the current challenges which will be faced by future generations such as: increasing the world population, increasing the soil, water and air pollution, increasing salinity and desertification worldwide, and decreasing the global crop yields etc. One important trait is the production of soluble sugars like trehalose in microorganisms and crop plants [4,180] under extreme environmental conditions. Trehalose, as an important compatible solute, offers resistance trait and other desirable features to these living organisms. Meanwhile, some other trials have succeeded in developing transgenic plants with trehalose biosynthetic genes, to offer novel strategies for improving abiotic stress tolerance in crop plants [181]. There have been more efforts as envisages and trials in some models from crops using bacteria and yeast, as a source of genes, to develop plants with significant ability to resist abiotic stress by accumulating trehalose sugar [182,183].
On the other hand, the accumulation of compatible solutes, might cause undesirable consequences on various aspects of plant growth and development, physiology and biochemistry. The following are some concerns that have been raised over the last three decades to support this argument: (1) the biosynthetic processes consume a significant amount of energy, in terms of ATP, and reducing power in terms of NADPH, (2) considerable amounts of metabolites (organic and amino acids) are used to build these compatible solutes to achieve osmotic adjustment and osmoregulation to maintain water balance and plant life at its lower levels rather than being used to build new cells, (3) the reduction in plant growth under these conditions is a consequence of the requirements of osmotic adjustment and osmoregulation, as plants sacrifice from their energy and growth to meet these requirements [49,184-186], (4) some authors have argued that the accumulation of compatible solutes is a response to the adverse effect of extreme environmental factors (salinity, drought and high and low temperatures) more than the advantages gained by plants in resisting these conditions. For example, while many studies have indicated the existence of a positive relationship between the accumulation of compatible solutes with stress tolerance, some studies have suggested that the increase in their concentrations under stress is a product of stress rather than being an adaptive response [187,188], and (5) at certain levels, some compatible solutes might have negative impacts; toxic effect on growth, development and metabolism [186]. Trehalose was given as an example, in spite of its role in maintaining growth and protecting the tissue machinery under osmotic stress, Wingler [103] has suggested that genes for trehalose biosynthesis in Arabidopsis and in a number of crop plant species might give a clear indication that the ability to synthesize trehalose is widely distributed in the plant kingdom. Others [90] have suggested that the reason behind the lack of trehalose accumulation in plants might be as a result of the presence of trehalase enzyme; as it was detected in some tissues of higher plants like pollen grains and the root nodules of legumes [105], and by inhibition of this enzyme, trehalose was detected in some plants. These results are controversial issues as trehalose might have adverse effects at certain levels of accumulation [57,174,189]. Living organisms seem to have safe guards, by one way or another; which can be used when exposed to threats affecting their life, and certainly a sacrifice should be given from their growth to sustain their machinery.
Therefore, there is always a balance between loss and profit, even in the world of biology when exposed to extreme environmental conditions.
Authors would like to thank the Department of Biological and Environmental Sciences, Department of Biology and Department of Biological Sciences at Universities of Qatar, Sanaa, and Al-al-Bayt, in Qatar, Yemen and Jordan respectively for the great supports to publish this work. The authors acknowledged the editing process was done by: QUEEN'S ENGLISH.