Journal of Sleep Disorders & Therapy
Open Access
ISSN: 2167-0277
ISSN: 2167-0277
Research Article - (2016) Volume 5, Issue 2
It is known that light in the morning is able to induce phase advances of the endogenous clock, however most studies have tested the phase advances in highly controlled laboratory conditions. At home the environmental lighting is more variable. In theory, a high intensity short morning-light pulse in the short-wavelengths-range (blue light) should be capable of inducing phase advances. If this is also true in a home setting, this could be a firm basis supporting light treatment in late chronotypes who suffer from a late sleep phase. In a study carried out in summer, 11 normal to relatively late (habitual midsleep 4:15-6:09 hours) chronotypes (age range 23-27 years, 4f/7m) participated in two conditions: (1) 1 baseline day followed by 3 consecutive days of 30 min. blue morning-light pulses, (2) 1 baseline day followed by 3 consecutive days of 60 min. blue light pulses. Blue light was applied by use of the Philips GoLite BLU (HF3320, blue leds, intensity at the cornea 2306 melanopic-lux, 300 lux, 3.65 W/m2). During all four evenings, subjects were asked to protect themselves from light exposure (<10 lux). The response of the melatonin rhythm, calculated as a shift in dim light melatonin onset (DLMO), to a single 30 min. light pulse varied between subjects and resulted in a non-significant phase advance of 15 (±48) min. (t10 = 1.04; P = 0.33), and a significant advance of 30 (±41) min. to a single 60 min. light pulse. (t10 = 2.40; P < 0.05). After 3 days of light exposure both in the 30- and in the 60 min. light pulses condition significant phase advances of DLMO were observed; 49 (±58) min. (t10 = 2.80; P < 0.05) and 59 (±29) min. (t10 = 6.9; P < 0.001) respectively. No significant differences were found in the DLMO shifts between conditions. In addition there was a trend for a lower sleepiness score directly after waking up after using light for 3 days in both conditions (t10 = 3.38; P = 0.096, 60 min. and (t10 = 4.10; P = 0.070, 30 min.). A prelimin.ary analysis of actimetry data indicated some support for an effect on sleep timin.g. The data support the conclusion that light pulses of 30 min. in the morning on three consecutive days, in a home setting, in combination with dim light during the evenings, can be part of an efficient phase advancing chronotherapy protocol.
Keywords: Blue light; Home-setting; Melatonin; Phase advance; Light therapy; Late chronotypes; Sleep
In humans, the endogenous clock, located in the Suprachiasmatic Nuclei (SCN) in the hypothalamus, is responsible for generating oscillations with a period of approximately 24 hours in physiological, behavioural and psychological variables. In order to keep these circadian rhythms entrained to the natural alternation of light and darkness, this clock needs to be synchronized daily.
Light is the main synchronizer, also called ‘Zeitgeber’, for entrainment of our clock; the exposure of light induces phase shifts of our clock resulting in either a delaying (biological rhythms shift to a later clock time with light exposure in the evening) or advancing (biological rhythms shift to an earlier clock time with light exposure in the morning) direction [1,2]. Since light is a potent Zeitgeber, the use of light therapy to treat circadian rhythm sleep disorders is a potentially effective treatment [3-5]. However, most studies have tested the phase shifting effects of light in highly controlled laboratory conditions. At home, the situation is completely different.
Subjects are exposed to a highly variable environment with large fluctuations in daytime light exposure, natural light exposure in the sleeping room etc. The current study was set up to test whether short pulses of light in the morning, in such a highly varying environment, are able to induce significant phase advancing shifts of circadian rhythms within 1-3 days to make it useful to treat late timing of biological clocks and sleep phases.
The sleep-wake cycle in humans is one of the rhythms that show a clear 24h pattern; the generation of this 24-rhythm is partly under control of the biological clock [6-8]. If exposed to a regular light-dark cycle, the sleep-wake cycle will be entrained; in other words, sleep would occur at the same phase of the light-dark cycle each day.
In reality, sleeping out of phase with the external light-dark cycle, especially sleeping late, is very common in our civilized society and is related to health problems [9-12]. A large number of people may therefore benefit from a correction of their clock and sleep timing. Because light is so effective in phase shifting the clock, it is often proposed as the most effective method to advance a late sleep phase [3-5]. However there is no general accepted protocol for the optimization of light therapy.
There are several factors that are important in optimizing light therapy. One is, like the correct timing of light. Phase response curves (PRC’s) have been measured to quantify the induced phase shift of the biological clock as a function of the time at which the subject is exposed to light. The most complete phase response curve (PRC) in humans has been published by Khalsa and colleagues [13]. The authors found the maximum phase advance when applying the center of a long 6.7-hour light pulse 9 hours after the start of the rise of melatonin, also known as the ‘dim light melatonin onset’ (DLMO). In a laboratory study of St. Hilaire and coworkers [14] a PRC with a hour white light pulse was tested. The results revealed that also for pulses of 1 hour the largest phase advances are found when timing the center of the light pulse around 9 hours after DLMO.
The intensity and duration of light exposure are other important factors in optimizing light therapy; both these factors contribute in a non-linear, dose dependent manner to the magnitude of a phase shift [15]. Constructed a dose response curve for light intensity with long (5 hours) light pulses. They found that high intensity full spectrum white light doses (9500 lux) generated the largest phase shifts.
Under these conditions, saturation of the logistic regression curve occurred already at one tenth (1000 lux) of the intensity of a maximum dose. This dose still generated a phase shift of 90% of the maximum. Under natural circumstances people are not dark adapted and they may need higher light intensities to show similar phase shifts.
The dose response curve for the duration of a light pulse is also not linear. The 1h light pulses in the study of St. Hilaire et al. [16] in humans elicited a maximal phase advance of 1.2h, which is 44% of the response of 2.7h generated by the 6.7h light exposure in the study of Khalsa et al. [13]. Such dimin.ishing responses to extended stimulation are considered the result of adaptation processes; therefore the beginning of a light pulse is most important for the phase shifting effects [17].
In addition to timing, intensity and duration, a fourth important factor that can be used to optimize light therapy are the spectral characteristics of the light source. There is accumulating evidence that both light-induced phase shifts-as well as the suppression of melatonin-are most sensitive to light with short wavelengths: 440-490 nm, or blue light [18-23]. This sensitivity to blue light is due to the protein melanopsin, located in the “intrinsically photosensitive retinal ganglion cells” (ipRGC’s) [24,25]. This photopigment is most reactive to ~480 nm light [25-27].
In a recent study, a comparison was made between the PRC obtained by Khalsa and colleagues [13] to 6.7h full spectrum white light with the PRC to blue light pulses (monochromatic 480 nm light) of similar duration (6.5) [28]. Although the blue light pulse contained only 0.4% of the energy of the full spectrum white light pulse, no significant differences could be found in the magnitude of the phase shifts.
Since blue light is very effective in shifting our circadian system, it may potentially decrease the intensity and duration of light therapy needed for an optimal effect and therefore improve therapy compliance, and final treatment success.
Therefore the hypothesis of this study is, whether optimally timed, short light pulses of 30 min. per day with a high blue photon density, applied by the subjects themselves at home, are able to shift the circadian rhythm of melatonin. If so, it can be an effective tool in the treatment of a delayed biological clock and sleep phase.
Subjects
Subjects were recruited from the student population of the University of Groningen, the Netherlands, via newspaper- and internet advertisements. Thirteen healthy subjects (seven men and six women, mean age (±SD) 23.9 ± 1.9 years) were selected and completed the protocol. During the analysis, two subjects were excluded due to noncompliance and possible improper timing of the light pulses as a consequence; analysis is performed on 11 subjects (4f/7m, mean age (±SD) 24.1y ± 1.6y, range age 23-27y).
Selection of subjects was based on the following criteria: Subjects had to be healthy without suffering from depressed mood (Beck Depression Inventory-II, Dutch version, (BDI-II_NL) < 8; [29,30], did not report sleep disturbances, were non-smokers, did not consume too many drinks with caffeine (no more than 8 units per day) or alcohol (no more than 10 units per week), did not work in night shifts, did not cross more than two time zones in the preceding three months and were not colour blind (tested with an Ishihara colour blindness test). Female subjects were only included if they were using hormonal contraceptives. Chronotype was determined using the habitual mean midsleep (hMS, this is the clock time at the midpoint between sleep onset and sleep end), based on 7 days of data weighted for day of the week (Munich Chronotype Questionnaire (MCTQ)) [31].
Subjects were selected if their habitual midsleep was between 4:15 and 6:09 hours, indicating normal to moderately late chronotype for the age category of 20-30 years (based on the MCTQ 2008 NL database, Gordijn et al. unpublished) [17]. This selection was on the premise that inducing possible phase advances of endogenous rhythms and sleep would be more advantageous for relatively late people. The average hMS in the selected subjects in this study was (±SD) 5:00 ±35 min. All subjects were Dutch citizens from birth and fluent in Dutch. All subjects gave written informed consent and were compensated for their participation. The experimental protocol was approved by the Medical Ethics Committee of the University Medical Center Groningen.
Melatonin profiling day
In order to estimate the optimal timing of the blue light pulses, the experiment started with a ‘melatonin profiling day’ in our human time isolation facility centre, Groningen (NL). Subjects arrived 10 hours before their habitual midsleep and left 5 hours after their habitual midsleep (used as an individual reference time). They collected a total number of twelve saliva samples in dim light (<10 lux) during the evening or in darkness during short moments of awakening in the night and early morning. Saliva was collected by chewing on a cotton swab (Salivette®, Sarstedt BV, Etten-Leur, NL).
At least 1 ml. of saliva per sample was collected every hour during the time subjects were awake and every two hours during scheduled sleep time. The saliva samples were centrifuged and frozen at -80?C for further analysis. Melatonin concentration was assessed by radioimmunoassay (RK-DSM; Bühlmann Laboratories, Alere Health, Tilburg, The Netherlands). The inter assay variance was 11.3% for the low concentrations and 12.2% for the high concentrations. The intra assay variance was 10.7% for the low concentrations and 10.0% for the high concentrations.
A bimodal skewed baseline cosine function was fitted through the individual data points [32]. The maximum value of the fitted curve of each individual was set at 100% and used to normalize each curve. Dim Light Melatonin Onset (DLMO) was calculated as the moment at which the melatonin rhythm crosses the 25% value of this fitted maximum by linearly interpolating the raw values preceding and following the 25% value. The average DLMO time on the melatonin profiling day was 22:20 (±0:15 SEM).
Experimental design
Subjects participated in two experimental conditions in a random order: a 30 min.ute and a 60 min.ute blue light pulse condition. The first experimental condition started at least one and not more than 2 weeks after the ‘melatonin profiling day’. This allowed for full recovery from the sleep interruptions during that day. Each condition consisted of 4 consecutive days, starting with 1 baseline evening/night followed by 3 mornings with light pulses (Figure 1).
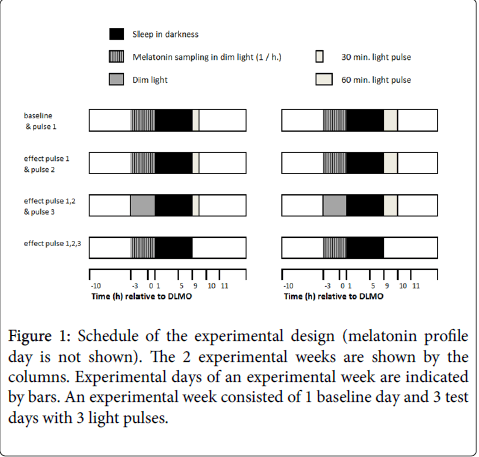
Figure 1: Schedule of the experimental design (melatonin profile day is not shown). The 2 experimental weeks are shown by the columns. Experimental days of an experimental week are indicated by bars. An experimental week consisted of 1 baseline day and 3 test days with 3 light pulses.
There was at least 1 week and not more than 2 weeks of no interventions in between conditions. All light pulses and measurements were conducted at home. During the week prior to each condition, subjects were asked to schedule their sleep according to their habitual sleep timin.g. This was checked with actigraphy and data from 8 subjects was available. During the week before starting the experiment hMS was on average (±SD) 2 min. (±13) earlier than the hMS calculated from the MCTQ questionnaire.
From these data it seems fair to conclude that the participants had consistent sleep habits prior to the start of the experiment. During each experimental condition, subjects received instructions when to go to sleep, when to stay in dim light and how and when they had to expose themselves to the blue light pulses. Sleep timing during the experimental weeks was adjusted to the timing of the light pulses in order to allow for 8 hours of sleep (Figure 1). Effect of the light pulses on the melatonin rhythm was measured by analyzing saliva samples collected after 1 pulse i.e., on the 2nd evening and after 3 pulses i.e., on the 4th evening and compared to melatonin pattern at the baseline evening i.e., 1st evening. During these evenings, subjects were required to collect the saliva samples themselves (Figure 1, starting at -3 and ending +1 hours relative to DLMO calculated from the melatonin profiling day) in dim light (<10 lux).
This resulted in 5 samples per evening. Subjects were asked to write down the exact timing of each saliva sample collection; no single timing point deviated more than 5 min. from the time point in the instructions. Subjects already knew how to collect saliva, because they had prior experience during the melatonin profiling day in the lab. At home, they were instructed to stay in dim light starting one hour prior to the first sample, and not change posture 5 min. prior to and during collection of the sample. They were not allowed to drink anything else than water, nor to eat during the sampling period, starting one hour prior to the first sample. They were instructed to rinse their mouth 15 min. prior to each sample.
Light exposure
Subjects were asked to expose themselves to either the 30 min. or 60 min. high intensity blue light pulses by sitting in front of the GoLite Blu lamp with an angle of 45° (Philips GoLite BLU HF3330, peak transmission at 470 nm, intensity at the cornea 2306 (m (elanopic)-lux (See Lucas et al. [33] for an explanation of the unit ‘m-lux’), 300 lux, 3.65 W/m2)) during 3 consecutive mornings. Light exposure was scheduled 9h after DLMO (Figure 1), as determin.ed from the melatonin profiling day. Because the average DLMO time on the melatonin profiling day was 22:20 (±0:15 SEM), light exposure started at 07:20 (±0:15 SEM) on average.
During all four evenings of each condition, including the one evening without saliva collection, subjects were asked to stay in dim light (<10 lux).
Rest-activity and light measurements
In order to quantify the timing and quality of the 24 hours profile of rest and activity, subjects wore an activity monitor, called Actiwatch® (AW2, Philips/Respironics, Fypro, Geffen, NL), on the non-domin.ant wrist during the experimental weeks. The AW2 is an activity-recording device that measures wrist movements to quantify the rest-activity cycle and it records environmental light exposure. The device has the appearance and the weight of a watch (without a clock). Subjects were asked to wear the AW2 all day and night.
While analyzing the data of all 13 subjects, we carefully inspected the sleep offset times of the AW2 data. We excluded 2 individuals who did not wake up within one hour of the instructed wake-up time. We concluded that these subjects did not comply with the instructions, and could not have had their light exposure at the proper time.
In addition, to measure properly light exposure and to be able to check for compliance of the light treatment the subjects wore either a Lightwatcher (Object Tracker, Vienna, Austria) or a Daysimeter (Lumen Tech Innovations, New York, USA). Unfortunately, because of failure of devices, the differences between devices and the way of wearing the devices, a complete data analysis of environmental light exposure in a quantitative way was not succesful.
In those cases where we could analyze whether subjects received a light pulse at the proper time, compliance was 100% in the 60-min. light pulse condition (10 subjects) and on average 87% in the 30-min. light pulse condition (1 pulse could not be recognized in 4 out of 7 subjects). Since this was a within study and compliance in the observed data was 100% in the 60-min. condition, it was concluded that the missing pulses in the 30 min. condition were most likely due to not wearing the device in the proper direction, more than that subjects did not obey the rules.
Questionnaires
Each morning of the experimental days, subjects were asked to complete a sleep diary. The diary consisted of a list of questions about sleep timin.g, sleepiness and sleep quality. Subjects rated their sleep quality with marks (from 0 = Very bad to 10 = Excellent). Subjective ratings of sleepiness were scored with the Karolinska sleepiness scale (KSS) (Akerstedt and Gillberg, 1990). Ratings on the KSS range from 1 to 9, with 1 meaning very alert and 9 meaning very sleepy. Subjects had to rate their sleepiness immediately after waking up and after the experimental light exposure.
Statistical analysis
For the differences in timing of DLMO, sleep onset, sleep offset, sleepiness and sleep quality between conditions, a repeated-measures ANOVA was used with one factor for condition (2 levels; 30 min. and 60 min.) and one factor for the number of pulses (4 levels; 0-3). If the factor ‘condition’ or the interaction of ‘condition *pulses’ was significant, a ‘simple contrasts’ test (equal to a paired sample T-test) was included to the ANOVA to see how many pulses were needed to see significant differences between conditions.
The conditions were also tested separately for the effects of the number of pulses. This was done with a repeated-measures ANOVA with one factor for the number of pulses and ‘simple contrast tests’ were used to compare the number of pulses to zero pulses (baseline day). The significance level was defined as p = 0.05. All parameters were tested two-tailed.
DLMO shifts
The salivary melatonin level rose in the evenings of all three nights in the two conditions (Figure 2). The 25% level of the maximum fitted value (definition of DLMO, see methods) is indicated by the black horizontal lines. The median DLMO times in the condition with 30 min. light pulses were 23:02h (baseline day i.e., 0 pulses), 22:47h (2nd day i.e., after 1 light pulse) and 22:24h (4th day i.e., after 3 light pulses). The median DLMO times in the condition with the 60 min. light pulses were 23:23h (0 pulses), 22:50h (after 1 light pulse) and 22:08h (after 3 light pulses). See also Figures 2A and 2B.
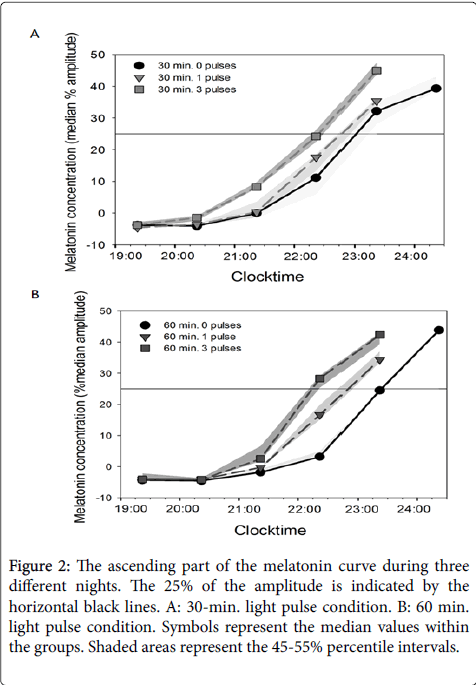
Figure 2: The ascending part of the melatonin curve during three different nights. The 25% of the amplitude is indicated by the horizontal black lines. A: 30-min. light pulse condition. B: 60 min. light pulse condition. Symbols represent the median values within the groups. Shaded areas represent the 45-55% percentile intervals.
Exposure to one 30 min. light pulse, actually timed 8.4 (SD 1.2) hours after baseline DLMO the night before, did result in a nonsignificant phase advance of the melatonin rhythm of 15 (±15 min. SEM) (t10 = 1.04; P =0.33). Exposure to one 60 min. light pulse, timed 8.2 (SD 0.8) hours after baseline DLMO the night before, resulted in a significant phase advance of 30 (±12 min. SEM) (t10 = 2.40; P < 0.05).
The actual timing of the first light pulses in the 30 min. blue light pulse condition and the 60 min. blue light pulse condition, although both a bit earlier than the scheduled 9h after DLMO, did not significantly differ from each other (t = 0.90; df = 10; P = 0.39). After 3 days of light exposure both 60- and 30 min. light pulses were able to elicit significant phase advances of the melatonin rhythm; a phase advance of 49 (±17 min. SEM) (t10 = 2.80; P < 0.05) resulted from exposure to three 30 min.
light pulses and a phase advance of 59 (±8.7 min. SEM) (t10 = 6.9; P < 0.001) from exposure to three 60 min. light pulses (Figure 3). Interestingly, no significant differences were found between the DLMO shifts after 30 min. light pulses and 60 min. light pulses: neither the shifts after one pulse (F1,10 = 0.96; P = 0.35) nor after three consecutive days with light pulses (F1,10 = 0.291; P = 0.65) were found to be significantly different from each other (Figure 3).
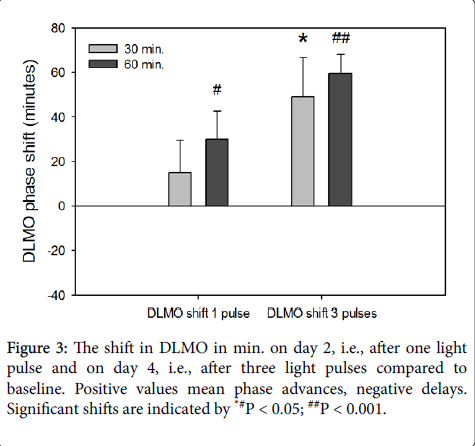
Figure 3: The shift in DLMO in min. on day 2, i.e., after one light pulse and on day 4, i.e., after three light pulses compared to baseline. Positive values mean phase advances, negative delays. Significant shifts are indicated by *#P < 0.05; ##P < 0.001.
Sleepiness patterns over the week
In order to see possible differences in the decline in sleep inertia, participants were asked to provide their sleepiness scores with the Karolinska Sleepiness Scale (KSS score) 5 min. after waking up, as well as 30 min. after waking up or after using the GoliteBLU lamp for 30 or 60 min. The decline in sleep inertia was found to be significant on all days, however no differences in sleep inertia decline were found for the interaction of the number of pulses and conditions (Pulses X Condition F3,8 = 0.81; P = N.S).
There were small trends for different KSS scores immediately after waking up after 2 and 3 pulses. In the 60 min. condition there was a trend for a lower KSS score immediately after waking up after 3 pulses 5.9 (±0.5 SEM) (t10 = 3.38; P = 0.096) compared to baseline KSS score 6.4 (±0.5 SEM). A similar trend for a decline in sleepiness immediately after waking up was observed in the 30 min. light condition; after 2 pulses the KSS score was on average 6.6 (±0.5 SEM) (t10 = 4.23; P = 0.067) and after 3 pulses 6.1 (0.6 SEM) (t10 = 4.10; P = 0.070), compared to the average baseline KSS score 7.6 (0.5 SEM).
The sleepiness scores in the 30 min. and 60 min. light pulse conditions were not significantly different from each other, not immediately after waking up (F1,10 = 2.89; P = 0.12), nor after using the GoliteBLUE lamp (F1,10 = 0.98; P = 0.35).
Sleep timing
To optimize the timing of the light pulses and to allow for 8 hours of sleep per night, subjects received individual instructions when to go to bed and when to wake up. Therefore, sleep timing was influenced by scheduled sleep and wakeup times. However, subjects were allowed to sleep and wake up earlier (not later) if they wanted.
After the experiment, the actual sleep timing was determined from data from the Actiwatch (AW2) that subjects wore on all experimental days, or from the sleep diary, if data from the AW2 was not available, which was the case for 4 out of 11 subjects. Sleep timing data, i.e., average (±SE) sleep onset and sleep offset times for the baseline night and for the nights after one, two or three light pulses are shown in w.
Sleep times in the 30 min. and 60 min. light conditions were not significantly different from each other, not for sleep onset (F1,10 = 0.17; P = 0.69), nor for sleep offset (F1,10 = 1.90; P = 0.20). Both experimental conditions resulted in a significantly earlier sleep onset compared to baseline (Table 1), though this was obtained later i.e., after 3 pulses in the 30 min. light pulse condition and already after 1 pulse in the 60 min. condition.
| Sleepparameter | condition | N | baseline | 1 lightpulse | 2 lightpulses | 3 lightpulses | |
| Sleep onset (h:min.) | 60 min.* | 11 | 0:42 (0:16) | 0:00 (0:19)* | 0:12 (0:18)* | 0:00 (0:13)* | |
| Sleep offset (h:min.) | 60 min. | 11 | 7:24 (0:15) | 7:24 (0:16) | 7:24 (0:16) | 7:24 (0:15) | |
| SOL (min.) | 60 min. | 7 | 4.3 (2.6) | 4.3 (2.4) | 8.7 (3.2) | 17.3 (5.4) | |
| Duration (h.) | 60 min.* | 7 | 5.8 (0.2) | 6.5 (0.1)* | 6.3 (0.2) | 6.2 (0.2) | |
| Awakenings/h. | 60 min. | 7 | 4.3 (0.4) | 4.2 (0.4) | 4.5 (0.4) | 3.8 (0.4) | |
| WASO (min.) | 60 min. | 7 | 44.4 (4.3) | 50.1 (2.6) | 55.0 (5.4) | 41.3 (7.7) | |
| SQR(Dutch marks) | 60 min. | 11 | 6.4 (0.3) | 6.4 (0.5) | 6.5 (0.3) | 6.4 (0.4) | |
| Sleep onset (h:min.) | 30 min.* | 11 | 0:36 (0:14) | 0:24 (0:23) | 0:06 (0:22) | 0:00 (0:12)* | |
| Sleep offset (h:min.) | 30 min. | 11 | 7:36 (0:19) | 7:30 (0:17) | 7:30 (0:15) | 7:30 (0:16) | |
| SOL (min.) | 30 min.* | 7 | 20.6 (3.5) | 6.7 (3.0)* | 9.1 (3.2) | 10.7 (3.3) | |
| Duration (h.) | 30 min.* | 7 | 5.9 (0.2) | 6.2 (0.2) | 6.1 (0.3) | 6.4 (0.1)* | |
| Awakenings/h. | 30 min. | 7 | 4.6 (0.4) | 4.3 (0.4) | 4.2 (0.4) | 4.3 (0.5) | |
| WASO (min.) | 30 min. | 7 | 50.0 (3.7) | 48.9 (3.9) | 55.3 (9.3) | 53.7 (7.9) | |
| SQR (Dutch marks) | 30 min. | 11 | 5.8 (0.5) | 6.3 (0.3) | 6.4 (0.5) | 6.3 (0.3) | |
| Italic parameters= Conditions are significantly different P<0.05 | Italic numbers = Significantly different from baseline P<0.05 | ||||||
| WASO= min. awake after sleep onset | SOL= Sleep onset latency | SQR=Sleep quality rating | |||||
Table 1: Sleep parameters with either 30 or 60 min. light pulses.
Sleep quality
Objective sleep quality was analysed from the data of the AW2’s (Table 1). Due to failures of the AW2’s in data collection, data on sleep quality of only 7 subjects (2f/5m) could be analysed. Objective sleep quality measures were sleep onset latency (SOL), sleep duration (Duration), the number of awakenings per hour (Awakenings/h) and min. awake after sleep onset (WASO). The subjective sleep quality rating (SQR) was asked for in the sleep diary on experimental days (Table 1).
The 30 min. and 60 min. conditions were only found to be significantly different for sleep onset latency (SOL) (F1,6 = 10.9; P < 0.05). This difference may have been caused by the longer sleep onset latency on the baseline day in the 30 min. condition. For the other sleep quality parameters there were no significant differences between conditions: sleep duration (F1,6 = 0.50; P = 0.51), number of awakenings/h (F1,6 = 0.29; P = 0.61), WASO (F1,6 = 1.15; P = 0.33), SQR (F1,10 = 0.45; P = 0.52).
Compared to baseline, sleep duration was longer for the 60- and 30 min. conditions on the 2nd and 4th day respectively. Sleep onset latency was shorter on the second day (after one light pulse) in the 30 min. condition compared to the baseline day. Again, this may have been caused by the longer sleep onset latency of the baseline day in the 30 min. condition.
This study demonstrates that three optimally timed, high blue light intensity, short pulses (30 min.), applied in a home situation, accompanied by dim light prior to sleep, are able to significantly phase advance the melatonin rhythm. The phase shifting effects of these three short blue light pulses are not significantly different from the effects of the positive control condition with three longer (60 min.) blue light pulses. The effects on the sleep-wake rhythm and morning sleepiness were also not significantly different between the 30-min. and the 60- min. blue light pulse conditions.
The idea that short light pulses are able to influence the circadian system is not new. However, up till now the positive findings were based on the effects of short light pulses mostly in highly controlled lab conditions. A dose response study of light pulse duration by Chang and colleagues (2012) revealed that the melatonin rhythm can be phase-delayed by a single light pulse in the evening of only 12 min. A more recent study of Gabel and colleagues (2013) showed that the melatonin rhythm can also be phase-advanced by a single blue light pulse in the morning of only 20 min.
Both these studies were carried out with subjects living in a controlled situation with a dimly lit environment outside the light exposure period. In a recent other home setting study [34] the effects of 30-45 min. of bright white light were tested in 10 subjects suffering from Delayed Sleep Phase Disorder. In that study, light was administered in one out of four different conditions, including a control condition, at waking up in a protocol wherein subjects received instructions to wake up one hour earlier each day until the preferred rising time was reached during a 2-week period. Unfortunately possible effects of the treatment on DLMO could be assessed in only 4 subjects and no statistical effect of light was observed in this small group. Analysis of actigraphy data on sleep parameters during the second week of treatment did not show a significant difference between the four conditions.
In our study we asked the subjects to stay in dim light during the evenings, both to be able to accurately estimate the onset of melatonin, and to prevent a possible counteracting effect of evening light exposure on the phase shifting effect of the morning light pulse. It is possible that protection to evening light by itself has played a role in the observed phase advance, but this has not been tested. Nevertheless, it seems unlikely that an almost one hour phase advance is the result of four consecutive dim light evenings alone.
The current study confirms the results of the controlled lab studies [16,35] and supports the conclusion that a combination of short light pulses of 30 min. in the morning and dim light in the evening is also capable of advancing the melatonin rhythm when applied in a home situation with no real control over environmental light. This is the situation in which light treatment for circadian rhythm sleep/wake disorders predominantly will take place.
The main focus of the present study was to test the effects of short blue light pulses on the melatonin rhythm. Therefore the setup of the study was not meant to test the effects on sleep, and actimetry data could be obtained in only 7 out of 11 subjects. For this reason, the results on the effects of the experimental conditions on sleep should be interpreted with caution.
Nevertheless, in addition to the shift in the melatonin rhythm there was also a small, but significant shift (30-40 min.) in sleep onset time. This shift was not significantly different for the 30- and 60 min. light pulse conditions. The shift in sleep onset time might reflect a shift in the circadian pacemaker, or be the effect of the shift of the melatonin rhythm, or the fact that the participants were instructed to wake up earlier than that they normally did (higher sleep pressure in the evening), or a combination.
Another finding was a trend for a reduction of sleepiness in the morning. Not only after light exposure, but even directly at waking up, probably resulting from a shift of the circadian pacemaker influencing sleep propensity and daytime alertness [8].
In contrast to the previous lab study of Gabel et al. [35], who showed a significant phase advance after 120-min. light pulse, the current home study shows more individual variation in phase advances in response to shorter light pulses, resulting in a non-significant phase advance after 130-min. light pulse? The larger variation may result from the different environmental circumstances, as the lighting conditions are variable at home while more constant in the lab.s
In addition, due to less controlled conditions, both in light exposure and in accurate timing of saliva collection, the estimation of DLMO may be more variable by itself, resulting in the non-significant shift after one day with a 30-min. light pulse in this study.
Indeed, light in the morning is not the only factor responsible for the phase-advancing effects of light. The studies of Hébert et al., Chang et al. and Giménez et al. [36-38] revealed that the ‘light history’ is also important for the responsiveness to light. Participants in lab conditions with dim light environments are more dark adapted and are therefore more sensitive to light (Hebert et al., Chang et al.) [37].
Even in a real-life setting, but with a reduction in blue light exposure through blue reducing contact lenses, it is possible to influence the non-image forming responses to light (Giménez et al.) [38].
Another important factor for an optimal substantial phase advance is the complete pattern of light-dark exposure during the rest of the day (Appleman et al., Burgess et al.) [39,40]. Especially low light levels in the evening are essential for an optimal phase advance. It is even possible to phase advance the melatonin rhythm without extra light in the morning.
When subjects are forced to sleep 2-3 hours earlier than they normally do, in other words, if they do not receive light in the evening their melatonin rhythm can also be phase advanced (Gordijn et al., Sharkey et al.) [41,42].
Although the subjects were told to dim their lights during the experimental evenings of the current study, there may have been variation in these evening light settings. Besides, there were no light restrictions during the remaining part of the day, and this study was carried out in summer.
Therefore, the light exposure patterns between individuals and between days must have been very different and these differences in prior light history may have influenced the phase shifts.
It is against this background of varying daytime and evening time light exposure that we still found significant phase advances with three days of only 30 min. extra blue light exposure in the morning. This emphasizes the conclusion that blue light therapy in the morning is potentially highly effective as a treatment for circadian rhythm sleep disorders with a late sleep phase.
A limitation of this home study is the lack of objective compliance data for all subjects both to the light therapy/dark instructions and to the strict timing of the collection of saliva for melatonin analysis. The data that were obtained supported the fact that subjects indeed followed instructions to a large extent, but a 100% certainty is not possible.
For a follow-up study, it is interesting to specifically monitor the extra light that is received during the light therapy session in relation to environmental light conditions over the whole day, to investigate whether this indeed plays a role in the efficacy of morning light treatment. The attempt to do this in the current study failed due to technical problems. Also the lack of a control condition without experimental light exposure is a limitation of the study.
Expectations of subjects have been demonstrated to influence light treatment effects in depression studies (Putilov & Danilenko, Flory et al., Knapen et al.) [43-45] and it is possible that the instructions of sleep scheduling or being admitted to the study protocol by itself influenced the observed effects.
Although an optimal ‘placebo light’ condition to control for expectations and for the effects of the experimental conditions is not possible in studies on the effects of light therapy, including a control condition with a ‘placebo lamp’ with the same visual brightness in a next study is definitely needed.
From the current study it is clear that optimally timed, short duration, high intensity blue light pulses of 30 min. have high potential to be effective when used in a light therapy design for phase advancing the circadian system. This includes both a proper instruction of when and how long to use the light, as well as when to protect from light, e.g. in the evening. A light therapy protocol for clinicians would benefit from a more easy instruction for the timing of light than in the current study where timing of light was relative to DLMO.
Since the phase angle of DLMO and sleep seems to be highly variable, especially in late chronotypes, this issue should be solved in additional experiments. Also to be able to generalize the effects of these short blue light pulses to the population, larger groups- and more variation within participants is needed.
Finally, to be clinically relevant, it still needs to be demonstrated whether also the sleep-wake rhythm and sleep architecture will improve in response to blue light therapy and if morning sleepiness at waking up can indeed be reduced with a longer treatment protocol.