Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Short Communication - (2015) Volume 4, Issue 5
The neurotransmitter serotonin plays an essential role in a variety of physiological processes including learning and memory, mood, and neuromodulation. Consequently, dysfunction of serotonergic neurotransmission is associated with psychiatric diseases such as depression, anxiety disorders, and schizophrenia. Because the small number and vast arborization of serotonergic neurons in vivo impede their use in primary cell culture, stem cell-derived serotonergic neurons are applied as in vitro-models for serotonergic neurotransmission. Among others, 1C11 stem cells are used to generate serotonergic neurons in vitro, for example, to analyze the effect of antidepressant treatment on serotonin re-uptake or the function of microRNAs in serotonergic neurons. Since 1C11 stem cells differentiate uniformly into serotonergic neurons, there is no need to optimize terminal differentiation. Therefore the contribution of neurotrophic factors, such as the brain-derived neurotrophic factor (BDNF), to terminal differentiation of 1C11 stem cells is currently unknown. To bridge this gap, we differentiated 1C11 stem cells and determined the effect of endogenous BDNF release as well as of supplemented recombinant BDNF on neurite growth. With regard to the application of recombinant BDNF we also differentiated 1C11 stem cells in microfluidic isolation devices in order to determine whether the location of recombinant BDNF application may exert differential effects on neurite growth. Our data showed that endogenously synthesized and released BDNF is sufficient to form functional neuritic networks during terminal differentiation and that an exogenous BDNF source did not extensively promote neurite growth. However, the location of recombinant BDNF application did affect the microgrooves targeting behaviour of differentiating 1C11 stem cells in microfluidic isolation devices. In summary, our data shows that 1C11 stem cells rely on synthesis and release of endogenous BDNF for serotonergic network formation during terminal differentiation.
<In the central nervous system, the neurotransmitter serotonin contributes to different physiological process including neuronal cell migration and differentiation, regulation of other neurotransmitter systems or modulation of the stress response. Due to these varied tasks, a malfunction of serotonin neurotransmission is linked to pathomechanisms of several neurodevelopmental and psychiatric disorders [1-3]. The serotonergic neurotransmission is maintained by a small population of neurons located in the raphé nuclei from where they build and maintain an extensive axonal network with projections densely innervating several brain regions, including hippocampus and cortex among others [4-7]. Based on their small number and morphological features in vivo, serotonergic neurons are not suitable to establish a primary neuronal culture. This obstacle has been overcome by establishing several experimental systems that display serotonergic features, for example blood platelets or immortalized cell lines. However, all of the models only possess a partial instead of a complete serotonergic phenotype, which comprises the capacity for active serotonin uptake, release, storage, synthesis and metabolism of serotonin (discussed in [8]). In 1990 the research group of Odile Kellermann introduced 1C11 stem cells that can be differentiated either into fully functional catecholaminergic [9] or serotonergic neurons [8]. Until establishing murine embryonic stem cell-derived serotonergic neurons [10,11], 1C11 stem cell-derived serotonergic neurons and embryonic raphé cultures were the only cellular in vitro-models that provided neurons with a complete serotonergic phenotype. Compared to embryonic raphé cultures, which are reported to yield only 5% serotonergic neurons out of a total yield of 30% neurons [12], 1C11 stem cells differentiate into a homologous serotonergic population within 4 days of terminal differentiation [8,9]. So far, among other studies, 1C11 stem cell-derived serotonergic neurons were applied to investigate different aspects of serotonergic neurotransmission including the internalizing effect of antidepressant treatments on the serotonin transporter [13-15], the role of microRNAs in antidepressant response [16, 17], and the use of fluorescent transporter substrates to visualize the release and re-uptake of serotonin in live cell imaging studies [18].
In contrast to 1C11 stem cells, primary raphé cultures were used in several studies as an in vitro-model to address molecular mechanisms in serotonergic differentiation. These studies showed that the simultaneous activation of brain-derived neurotrophic factor- (BDNF) and cAMP-dependent signaling pathways are essential for the induction of the serotonergic neuronal phenotype [12,19-21]. In addition to enhance the expression of serotonergic phenotype genes such as serotonin-synthesizing protein tryptophan hydroxylase-2 and of the serotonin transporter, the stimulation of these two signaling cascades also increases the number of serotonergic neurons derived from primary raphé cultures by at least 80% and up to120% [12,21,22].In accordance with BDNFs effect on neurite outgrowth, serotonergic neurons in primary raphé cultures also exhibit a significant 2-4 fold increase in neurite length by application of recombinant BDNF to the culture medium [12,22]. Since the induction of terminal differentiation of 1C11 stem cells results in a complete conversion of stem cells into serotonergic neurons, the beneficial effects of stimulating BDNF-/ cAMP-dependent signaling pathways to promote the serotonergic phenotype may not be required for this cell line. However, the question whether application of recombinant BDNF influences the terminal differentiation of 1C11 stem cells with regard to enhanced neurite outgrowth and formation of more extensive neuronal networks, has not been investigated yet. Furthermore, the question whether and to which extend BDNF synthesized and released by 1C11 stem cells contributes to the process of terminal differentiation also has not been addressed yet.
We have differentiated 1C11 stem cells into serotonergic neurons and analyzed the contribution of both, endogenously synthesized BDNF and recombinant BDNF on neurite growth during terminal differentiation. In order to find out, whether the location of recombinant BDNF application exerts differential effects during terminal differentiation, we have differentiated 1C11 stem cells using microfluidic isolation devices. Our results clearly revealed that application of recombinant BDNF during the terminal differentiation of 1C11 stem cells only slightly affected neurite outgrowth. Consequently 1C11 stem cells appear to be truly self-sufficient and provide all molecular requirements for terminal differentiation with regard to neuronal network formation and functionality.
Generation of stem cell-derived serotonergic neurons
1C11 stem cells were cultured and differentiated into serotonergic neurons as described previously [8]. Briefly, differentiation of 1C11 stem cells into serotonergic neurons takes 4 days and was induced by addition of 500 μg/mL dibutyryl cyclic cAMP and 500 μg/mL cyclohexanecarboxylic acid (both Sigma Aldrich). For immunofluorescence and morphological analysis, 20,000 progenitor cells/μm2 were plated in μ-slide 8 well dishes (Ibidi). Each day during terminal differentiation, 20 ng/mL recombinant BDNF and 0.4 μg/mL recombinant human TrkB Fc Chimera (TrkB-IgG; both by R&D Systems) were added individually or in combination to the differentiation medium of 1C11 progenitors. Differentiation medium of 1C11 progenitors cultured under control conditions was supplemented with the according amount of sterile PBS.
Immunofluorescence and morphological analysis
1C11 stem cells were immunostained using primary antibodies against BDNF (1:400; Novus Biological NBP1-03458), serotonin transporter (1:500; ATS AB-N09), and TrkB (1:100; Santa Cruz H-181 or N-20). Secondary antibodies applied were donkey-anti-sheep conjugated to AlexaFluor555 and donkey-anti-rabbit conjugated to AlexaFluor647 (1:2000; Life Technologies). For measurements of neurite length, cells were immunostained with primary antibodies against TuJ1 (Figure 1; 1:200; Synaptic Systems, #302302) and secondary antibodies were donkey-anti-mouse conjugated to AlexaFluor488 (1:2000; Life Technologies). Immunostaining of 1C11 stem cells was performed as described previously [14]. Confocal images were acquired on a Leica TCS SP5 equipped with an acusto-optical beamsplitter and a 63x objective. Confocal z-stacks were converted into sum projections using NIH ImageJ 1.45s software. Quantification of neurite length was performed using the NeuronJ plug-in [23]. Variance analysis with subsequent post-hoc analysis (Tukey test) was performed with the raw data generated by NeuronJ using the Graph Pad Prism software. In the figures, p-values <0.05 are indicated by *, p<0.01 by **, and p<0.001 by ***.
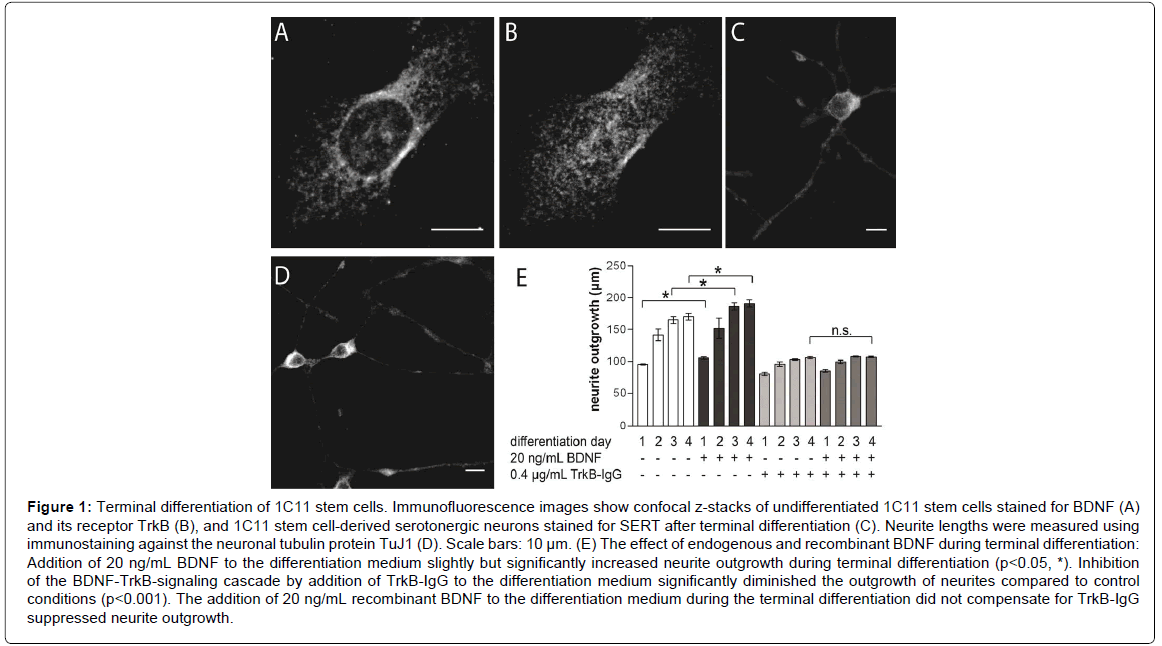
Figure 1: Terminal differentiation of 1C11 stem cells. Immunofluorescence images show confocal z-stacks of undifferentiated 1C11 stem cells stained for BDNF (A) and its receptor TrkB (B), and 1C11 stem cell-derived serotonergic neurons stained for SERT after terminal differentiation (C). Neurite lengths were measured using immunostaining against the neuronal tubulin protein TuJ1 (D). Scale bars: 10 μm. (E) The effect of endogenous and recombinant BDNF during terminal differentiation: Addition of 20 ng/mL BDNF to the differentiation medium slightly but significantly increased neurite outgrowth during terminal differentiation (p<0.05, *). Inhibition of the BDNF-TrkB-signaling cascade by addition of TrkB-IgG to the differentiation medium significantly diminished the outgrowth of neurites compared to control conditions (p<0.001). The addition of 20 ng/mL recombinant BDNF to the differentiation medium during the terminal differentiation did not compensate for TrkB-IgG suppressed neurite outgrowth.
Terminal differentiation in the presence of a BDNF-gradient in microfluidic isolation devices
To obtain and maintain BDNF gradient during the terminal differentiation of 1C11 stem cells, we applied 150 μm microgroove containing microfluidic isolation devices (MID, Xona Microfluidics), which maintain a gradient between the two compartments for up to 48 h. The devices were mounted in 3.5 cm culture dishes (Ibidi) as specified by the manufacturer. 5 μL cell suspension (5x106 cells/mL) of 1C11 stem cells were plated into the channel adjacent to the microgrooves in one compartment, which is defined as the cell compartment. Culture medium supplemented with 20 ng/mL recombinant BDNF was then added to the reservoirs of either the cell compartment (somatic BDNF application) or the opposing compartment, which is defined as the neurite compartment (remote BDNF application). 30 min after plating, the first images were acquired on a Leica DM IRB microscope equipped with a CCD colour camera (Olympus) as well as 10x and 20x objectives. Subsequent images were acquired every hour for 24h. Outgrowth of neurites was measured using the NIH ImageJ 1.45s software. Cells were considered targeting the neurite compartment as soon as contact between a cell’s soma or neurites with the opening of a microgroove was established (indicated by the dotted red line in Figure 2A). The speed of neurite growth was determined for neurites growing in microgrooves upon neurite contact to the opening of the microgroove. The raw data obtained was used for the statistical analysis. Variance analysis with post-hoc comparison was performed for the neurite growth speed; Fisher’s test for analysis of targeting microgrooves (Graph Pad Prism software). As described above, asterisks are assigned for the respective p-values in the figures.
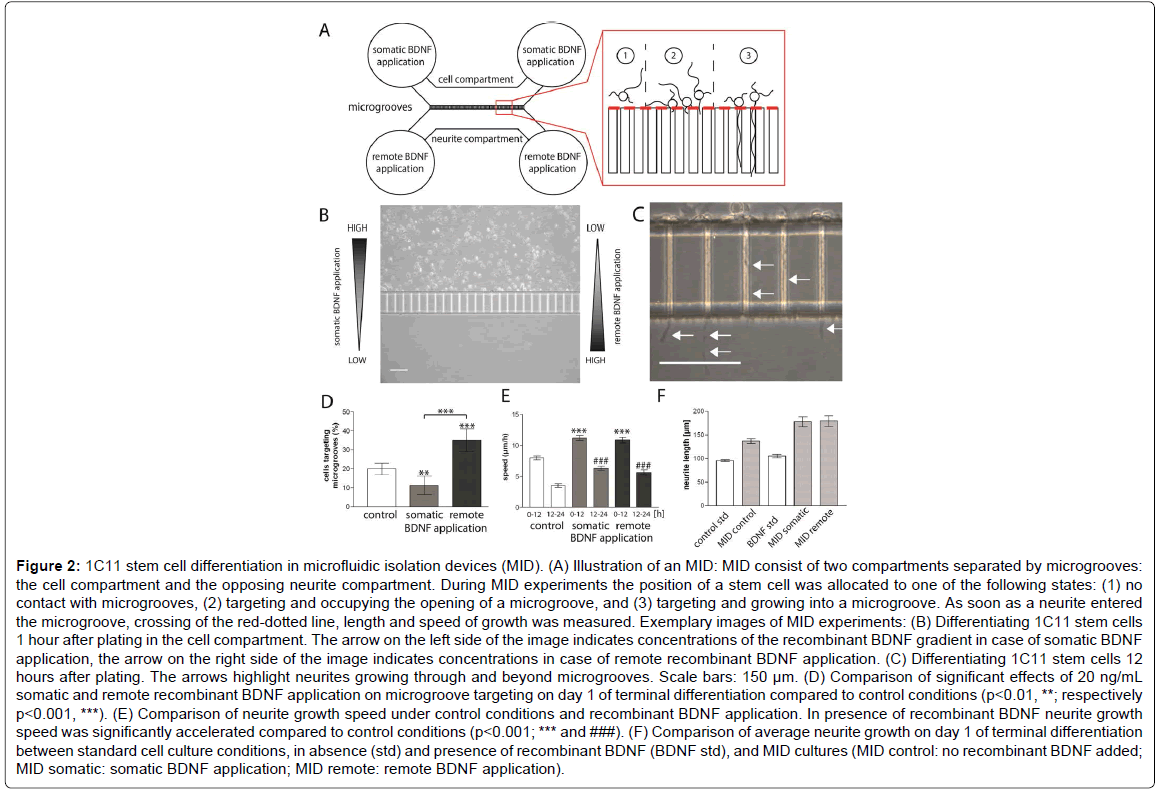
Figure 2: 1C11 stem cell differentiation in microfluidic isolation devices (MID). (A) Illustration of an MID: MID consist of two compartments separated by microgrooves: the cell compartment and the opposing neurite compartment. During MID experiments the position of a stem cell was allocated to one of the following states: (1) no contact with microgrooves, (2) targeting and occupying the opening of a microgroove, and (3) targeting and growing into a microgroove. As soon as a neurite entered the microgroove, crossing of the red-dotted line, length and speed of growth was measured. Exemplary images of MID experiments: (B) Differentiating 1C11 stem cells 1 hour after plating in the cell compartment. The arrow on the left side of the image indicates concentrations of the recombinant BDNF gradient in case of somatic BDNF application, the arrow on the right side of the image indicates concentrations in case of remote recombinant BDNF application. (C) Differentiating 1C11 stem cells 12 hours after plating. The arrows highlight neurites growing through and beyond microgrooves. Scale bars: 150 μm. (D) Comparison of significant effects of 20 ng/mL somatic and remote recombinant BDNF application on microgroove targeting on day 1 of terminal differentiation compared to control conditions (p<0.01, **; respectively p<0.001, ***). (E) Comparison of neurite growth speed under control conditions and recombinant BDNF application. In presence of recombinant BDNF neurite growth speed was significantly accelerated compared to control conditions (p<0.001; *** and ###). (F) Comparison of average neurite growth on day 1 of terminal differentiation between standard cell culture conditions, in absence (std) and presence of recombinant BDNF (BDNF std), and MID cultures (MID control: no recombinant BDNF added; MID somatic: somatic BDNF application; MID remote: remote BDNF application).
1C11 stem cells were immunostained with antibodies targeted against BDNF or its receptor TrkB. The BDNF immunostaining displayed a predominantly globular staining pattern in perinuclear as well as peripheral structures, most likely showing BDNF positive staining for ER and Golgi structures (Figure 1A). A similar distribution of immunosignals was observed for the TrkB immunostaining (Figure 1B). However the confocal projections revealed a less pronounced recess for the nucleus indicating that more TrkB molecules localize to perinuclear structures as observed for BDNF. A colocalization study of TrkB immunosignals and a nuclear staining dye clearly showed that nuclear localization of TrkB could be excluded (data not shown). These observations indicate that undifferentiated 1C11 stem cells already express the molecules required for activation of auto- or paracrine BDNF-dependent mechanisms, which are important for serotonergic differentiation in vitro [19,21], and that upon induction of the terminal differentiation, no immediate de novo-synthesis of BDNF and TrkB is required.
In vivo studies in mice showed that application of recombinant BDNF promotes regenerative outgrowth of neurotoxin-damaged serotonergic neurons [24,25]. Similar observations were made during the serotonergic in vitro-differentiation of neurons in embryonic raphé cultures, in which adding recombinant BDNF to the culture medium significantly enhances neurite growth [12,22]. To find out to which extent the addition of recombinant BDNF contributes to neurite outgrowth the terminal differentiation of 1C11 stem cells, we supplemented the differentiation medium with 20 ng/mL BDNF. Differentiating 1C11 stem cells were fixed and immunostained for TuJ1 in order to label and measure neurites (Figure 1D). Under control conditions we observed an outgrowth of serotonergic neurites of 95.4 ± 1.8 μm during the initial 24 hours (Figure 1E). During the next days of terminal differentiation the neurite outgrowth of our control cells reach an average of 141.5 ± 18.6 μm on day 2, and an average outgrowth of 164.9 ± 9.3 μm on day 3. Upon completion of the terminal differentiation on day 4, on which 1C11 stem cell-derived serotonergic neurons express the serotonin transporter (Figure 1C), the average neurite length under control conditions was 170.3 ± 9.6 μm (Figure 1E). The addition of recombinant BDNF to the differentiation medium slightly elevated the neurite length from 95 μm to an average neurite length of 105.3 ± 3.8 μm (Figure 1E; p=0.0321). The slightly enhanced neurite growth in the presence of recombinant BDNF was maintained during the following days of terminal differentiation. It peaked on day 4 with an average neurite outgrowth of 190.0 ± 9.6 μm, which is significantly longer compared to the average neurite length of 170 μm under control conditions (Figure 1E; p = 0.0396). Compared to the observations made during the differentiation of embryonic raphé cultures, where the application of recombinant BDNF resulted in a 2 – 4 fold increase of neurite growth [12,22], addition of recombinant BDNF during the terminal differentiation of 1C11 stem cells increased neurite growth by only 11%. Given this observation, we wanted to find out to which extend the release of endogenous BDNF contributes to neurite growth during the terminal differentiation of 1C11 stem cells. Therefore we have added TrkB-IgG to the culture medium, which inhibits BDNF binding to TrkB and thereby suppresses ligand dependent activation of the BDNF-TrkB-signaling cascade [26-28]. In presence of TrkB-IgG during the terminal differentiation, neurite outgrowth was significantly, but not completely diminished (p<0.001). Already on day 1, the addition of TrkB-IgG to the culture medium decreased neurite growth compared to control conditions by 15μm to an average neurite length of 80.6 ± 4.6 μm. The reduction of neurite length in the presence of TrkB-IgG was consistent during terminal differentiation and resulted on an average neurite length of 105.9 ± 3.1 μm on day 4 (Figure 1E), which is only 62% of the outgrowth obtained under control conditions. The daily addition of recombinant BDNF did not compensate for the diminished neurite outgrowth caused by the application of TrkB-IgG. In the presence of recombinant BDNF and TrkB-IgG, neurite outgrowth was similar to the single application of TrkB-IgG. In the end, the average neurite length on day 4 was 106.9 ± 1.9 μm and similar to the length observed in presence of TrkB-IgG on day 4 (Figure 1E).
Generally, 1C11 stem cells showed the strongest outgrowth during the first two days of terminal differentiation. Here, neurites already grew up to 80% of the total neurite length achieved at the end of terminal differentiation. In presence of TrkB-IgG neurite outgrowth on day 2 even amounted for at least 90% of total outgrowth. Here we would like to point out that we did not expect TrkB-IgG treatment to inhibit terminal differentiation of 1C11 stem cells. Indeed, in BDNF-deficient mice the number of serotonergic neurons is slightly increased by 25% compared to wild type and the expression of the serotonin transporter is elevated [29]. Taken together, our findings indicate that 1C11 stem cells contain all prerequisites to form a functional neurite network during terminal differentiation and exogenous neurotrophin sources may not essentially contribute to this network formation (Figure 1).
We also took into consideration, that the general application of recombinant BDNF in differentiation medium may obscure differential effects depending on the location of recombinant BDNF application. In vivo experiments in rodents showed that the regenerative outgrowth of serotonergic neurons from the raphé nuclei is induced by a prefrontal cortex perfusion with recombinant BDNF [24], and that a BDNF overexpression in the hippocampus resulted in increased serotonergic innervation [30]. Therefore we differentiated 1C11 stem cells in microfluidic isolation devices (MID) that feature two neighbouring compartments that are divided by a 150μm microgroove barrier (Figure 2). Plating 1C11 stem cells in one compartment of the MID (cell compartment) enables us to perform terminal differentiation while either exposing 1C11 stem cells to a chemical BDNF gradient, by supplementing the media of the neighbouring neurite compartment with recombinant BDNF (remote BDNF application), or to directly expose 1C11 stem cells to BDNF, by application of recombinant BDNF to the cell compartment (somatic BDNF application) (Figure 2).
Our first observation during the initial 24 hours of terminal differentiation using MID was that the location of recombinant BDNF application affected the targeting of microgrooves by 1C11 stem cells (Figure 2D). Under control conditions, 20% of the differentiating 1C11 stem cells targeted the microgrooves by intrinsic movement. Compared to this, the somatic BDNF application kept differentiating 1C11 stem cells from targeting the microgrooves in a similar degree. In this case, the partition of cells targeting microgrooves significantly dropped to 11.3% (p<0.01). Compared to the other experimental conditions, the remote BDNF application on the other hand significantly increased microgroove targeting by differentiating 1C11 stem cells up to 35% (p<0.001 in both cases). These findings showed that during the first day of terminal differentiation 1C11 stem cells were responsive to the chemotactical BDNF gradient inside the MID.
We also observed a general growth enhancing effect of recombinant BDNF on the neurite growth inside the microgrooves of MIDs (Figure 2E). Under control conditions neurites grew up to 7.9 μm/h inside the microgrooves during the initial 12 hours of terminal differentiation. Growth speed than decelerated to 3.5 μm/h till the end of day 1 of terminal differentiation. Both kinds of recombinant BDNF application significantly elevated the growth speed of neurites inside the microgrooves during the first day of terminal differentiation (Figure 2E, p<0.001). In case of somatic BDNF application, the growth speed of neurites inside the microgrooves averaged 11.2 μm/h for the initial 12 hours before dropping to 6.3 μm/h for the following 12 hours. The remote BDNF application elevated growth speed to a nearly similar speed of 10.8 μm/h during the first half of day 1. Here too, during the second half of the day the growth speed was reduced to 6.6 μm/h. In summary, the speed of neurite growth inside the microgrooves is significantly accelerated in the presence of recombinant BDNF independent of the location of the BDNF source. However, neither intrinsic nor BDNF-accelerated neurite outgrowth of differentiating 1C11 stem cells reached the speed of neurites sprouting from serotonergic neurons in vivo as determined by histological studies in rodent brains [31].
Interestingly, under control conditions, inside the microgrooves neurites grew up to an average length 136.8 μm ± 4.9 μm during the first day of terminal differentiation compared to an average of 95.4 ± 1.8 μm in normal culture dishes. The enhanced neurite growth may be caused by the three-dimensional microgroove surrounding of the neurite, since micropattern environments have an effect on cellular function and growth (for example [32,33]). For 1C11 stem cells differentiating in the presence of recombinant BDNF a similar observation was made. The average length of neurites growing inside the microgrooves reached 181.4 ± 8.5 μm for somatic BDNF application, and 184.9 ± 10.5 μm for remote BDNF application, while in standard culture dishes BDNF only slightly increased neurite growth up to 105.3 ± 3.8 μm. Our experimental data covered only the first day of terminal differentiation of 1C11 stem cells, since during the second day of differentiation microgrooves were evenly occupied under all conditions tested, and neurites, once grown into the neurite compartment, did not grow beyond the size determined on day 1 of terminal differentiation. Therefore further differences during terminal differentiation inside a MID could not be determined.
In conclusion, our experiments show that 1C11 stem cells provided all prerequisites for terminal differentiation into biochemically functional serotonergic neurons. The addition of recombinant BDNF only slightly enhanced neurite outgrowth in standard culture conditions but accelerated neurite outgrowth inside the microgrooves of microfluidic isolation devices. Given the circumstance that, compared to serotonergic neurons in vivo, 1C11 stem cell-derived serotonergic neurons only build a less extensive neuronal network, they are a most suitable in vitro-model to investigate the biochemical characteristics of serotonergic neurotransmission.
We sincerely thank Josef Frank, Dept. Genetic Epidemiology in Psychiatry (CIMH), for his help on statistic evaluation, Janina Ziegler, Biochemical Laboratory (CIMH), for her excellent technical assistance, and Friederike Matthäus, Biochemical Laboratory (CIMH), for her valuable comments on the manuscript. This work was funded by the DFG SFB636 to P.S.