Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2018) Volume 6, Issue 2
Wheat (Triticum aestivum L.) known as bread wheat (hexaploid), is an important cereal for millions around the world. Excessive salts in soil affect growth, development and productivity of crop plants and have been major constraints to agriculture. Plants, being sessile in nature, have developed mechanisms to cope with high salt concentrations in soil. To understand salt-tolerance in crop plant and its improvement, Total Phenolic Content (TPC), Lipid Peroxidation (LP), total Antioxidant activity (AO) and high affinity potassium transporters (HKTs) in shoot and root regions of contrasting wheat genotype WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) were studied on seed priming with two different biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). Primed seeds were sown in pots under controlled conditions in phytotron and were exposed to 150 mM NaCl stress after 14 days of germination. Expression pattern of HKT genes in the root and shoot of the primed and unprimed as well as stressed and unstressed seedlings were also studied using semi-quantitative and quantitative analysis. Higher levels of TPC, LP and AO activity were observed in the contrasting wheat genotypes under salt stressed condition. Both HKT1 and HKT3 genes were involved in regulating ion homeostasis. Up and down regulation of expression of HKT genes in shoot and root, respectively, provide resistance against salt uptake. Biotic elicitors (SA and MJ) were found to mitigate the effect of salt-stress by affecting expression of HKT genes as well their biochemical-processes. MJ showed better response than that of SA, and can be utilized to improve defense responses of crop plant against salt stress. The regulation of gene expressions and signaling cascades that regulate Na+ transporters remain to be elucidated and these studies will help in understanding the mechanism of ion homeostasis during salt stress to improve crop yield.
Keywords: Salt stress; High affinity potassium transporters; Wheat; Methyl jasmonate; Salicylic acid; Gene expression and RT-PCR
Wheat (Triticum aestivum L.), commonly known as bread wheat from the family Poacea, is the second most important staple food crop of India. Environmental factors such as light, carbon dioxide, temperature, salt, nutrients and water availability are important factors for the successful completion of a plant’s life cycle. Among these, salinity is one of the major problems in irrigated agriculture in most of the countries, particularly in wheat and rice growing areas. About onefifth of irrigated agricultural land is adversely affected by salinity. The main component to salinity stress in plants is an initial osmotic stress and a subsequent accumulation of toxic ions which negatively affects cellular metabolism. Salt stress inhibits plant growth through two major effects. First, osmotic or water-deficit effect, where, salt in soil water reduces the plant ability to take up nutrients through water, which eventually leads to reduced growth. Second, ion-excess effect, that is, accumulation of high level of toxic ions (e.g., Na+, Cl-) in the cytoplasm with a reduction in beneficial ions (K+, Ca2+) lead to oxidative stress, further reducing growth [1]. Sodium ions can enter the cell through several low and high-affinity K+ carriers. Some members of the HKT family function as sodium transporter and contribute to Na+ removal from the ascending xylem sap and recirculation from the leaves to the roots via the phloem vasculature. A correlation between organic osmolytes accumulation and plants have developed different physiological, biochemical and molecular strategies to adapt stress condition.
A common theme underlying responses to a range of biotic and abiotic stresses is the phenomenon of priming, whereby previous exposure makes a plant more resistant to future exposure. Primed plants display either faster and/or stronger activation of the various defense responses that are induced following attack by either pathogens or in response to abiotic stresses. The time interval that occurs between the priming event and the subsequent stress exposure, in which altered plant response is realized, is of considerable interest [2]. Pre-soaking of seeds with optimal concentration of plant regulators like Methyl Jasmonate (MJ) and Salicylic Acid (SA) has been shown to be beneficial to growth and yield of some crop species grown under saline conditions [3]. Salt stress induces a number of changes in plant metabolism like ion toxicity, osmotic stress and production of reactive oxygen species (ROS). It is the antioxidant potential of the plant that protects plant against the free radicals. It has been reported [4] that under stress conditions, malondialdehyde (MDA) accumulation takes place in plants due to membrane lipid peroxidation. Biochemical studies with reference to AntiOxidant (AO) activity, Total Phenolic Content (TPC) and Lipid Peroxidation (LP) of primed seeds would be useful in understanding the defense responses of plants under salt stress. Seed-priming has been found to increase tolerance against abiotic stresses, including, chilling, salinity, drought and heavy metals, in different plant varieties [5,6]. Seed-priming with Salicylic Acid (SA) enhanced moisture content, antioxidant activity and total chlorophyll content in wheat under drought stress [7]. Also, seedpriming with selenium (Se) and SA was found to be effective in protecting rice plants against chilling stress [8]. Recently, bio-elicitorinduced biochemical and molecular manifestations to improve drought tolerance in rice [9] and impact of seed priming on proline content and antioxidant enzymes to mitigate drought stress in rice have been already reported [10].
Adaptation of plants to salt stress requires cellular ion homeostasis involving net intracellular Na+ and Cl- uptake and subsequent vacuolar compartmentalization without toxic ion accumulation in the cytosol. Sodium ions can enter the cell through several low and high-affinity K+ carriers. Some members of the HKT family function as sodium transporter and contribute to Na+ removal from the ascending xylem sap and recirculation from the leaves to the roots via the phloem vasculature. Various mechanisms of salt tolerance in crops, and ion transporters, especially high affinity potassium transporters (HKT) have been found linked to a great extent with the salt tolerance mechanism of the plants. High-affinity Potassium Transporters (HKTs) belong to an important class of integral membrane proteins (IMPs) that facilitate cation transport across the plasma membranes of plant cells. HKT proteins have been characterized predominantly as monovalent cation transporters. HKT proteins-mediated transport of Na+ is known to be an important component of salinity tolerance in several species, including Arabidopsis , rice and wheat [11]. Heterologous expression of HKT gene in Arabidopsis thaliana enhancing salinity tolerance has been recently reported [12]. Researchers have tried to improve plant tolerance to salinity injury through either chemical treatments, or biofertilizer treatments to enhance a process used naturally by plants to minimize the movement of Na+ to the shoot, using genetic modification to amplify the process, in helping plants to do what they already do, but to do it in a much better way [13]. Plants live in complex environments in which they intimately interact with a broad range of abiotic and biotic factors with different strategies both constitutive and inducible. It has been reported that growth and yield in corn under drought stress generally increased with seed priming [14]. Recent advances in plant immunity research have provided exciting new insights into the underlying defense signaling network for their own benefits to antagonize the host immune response. Seed-priming with plant growth regulators and chemical compound is found to be effective in assuaging the damaging effects of abiotic stress in plants. Moreover, the biochemical, physiological and molecular basis of seed-priming to mitigate abiotic stresses in plants is becoming clear day-by-day. The results of the current research investigation will provide new insights that can lead to a better understanding of the biochemical and molecular basis of salt-sensitivity in wheat and its possible mitigation through biotic elicitors.
Seed material and seed priming
Seed material of two wheat genotypes-WH 542 (Salt-susceptible) and KRL210 (salt-tolerant) were procured from Directorate of Wheat Research, Karnal. The root and shoot region of these varieties were analyzed for morphological, Total Phenolic Content (TPC), Lipid Peroxidation (LP), total antioxidant activity (AO) and HKT gene specific expressions. Seeds of the contrasting wheat genotypes were sterilized with 0.1% HgCl2 solution prior to priming with biotic elicitors. Primed and Control seeds were grown in petri-plates under controlled conditions. Seed priming with biotic elicitor solutions of appropriate concentrations will be used according to the method given [3] for Methyl Jasmonate (MJ) and [5] Salicylic Acid (SA). After screening on the basis of germination rate and physiological growth, 0.1 mM concentration of each of the priming reagent was optimized for further analysis. Two sets of plants (sample name with their respective abbreviations) in triplicate for each SA and MJ (Table 1) were prepared and 10 seeds at equal distance were sown in the pots under controlled National Phytotron Facility. Salt-stress (150 mM NaCl) in half strength Hoagland solution was imposed on two weeksold plants. Three biological samples of shoot and root in triplicate were harvested on 14 days after treatment (DAT) for biochemical and molecular observations.
| S.No. | Abbreviation | Sample |
|---|---|---|
| 1 | UWC | Unprimed WH 542 control |
| 2 | PWSAC | Primed WH 542 with salicylic acid control |
| 3 | PWMJC | Primed WH 542 with methyl jasmonate control |
| 4 | UKC | Unprimed KRL 210 control |
| 5 | PKSAC | Primed KRL 210 with salicylic acid control |
| 6 | PKMJC | Primed KRL 210 with methyl jasmonate control |
| 7 | UWT | Unprimed WH 542 treated |
| 8 | PWSAT | Primed WH 542 with salicylic acid treated |
| 9 | PWMJT | Primed WH 542 with methyl jasmonate treated |
| 10 | UKT | Unprimed KRL 210 treated |
| 11 | PKSAT | Primed KRL 210 with salicylic acid treated |
| 12 | PKMJT | Primed KRL 210 with methyl jasmonate treated |
Table 1: Abbreviations used in sets of plants for biotic elicitors/ priming reagents.
Total Phenolic Content
Total Phenolic Content (TPC) was determined in root and shoot tissues of contrasting wheat genotypes method [15]. According to this method adding 0.5 ml of the aqueous extract to 2.5 ml of 10% Folin- Ciocalteu reagent (v/v) and 2 ml of 7.5% sodium carbonate. The reaction mixture was incubated at 45°C for 40 min and the absorbance was measured at 765 nm in the spectrophotometer. Gallic acid was used as a standard phenol [15]. The mean of three readings was used and the TPC was expressed as milligrams of gallic acid equivalents/g extract.
Reagents: i. 7.5% Sodium carbonate-Dissolved 15 g Na2CO3 in 200 ml distilled water.
ii. 0.1% TCA (Trichloroacetic acid)-Dissolved 0.1 g TCA in 100 ml distilled water.
iii. Gallic acid (stock 250 micro g/ml)-Dissolved 2.5 mg gallic acid in 10 ml distilled water.
Lipid peroxidation
Lipid peroxidation was measured as MDA in the shoot and root, according to Cakmak and Horst method. This method is based on the reaction with thiobarbituric acid. Fresh tissues of root and shoot (1.0 g each) were ground properly in 20 ml of 0.1% TCA solution and centrifuged for 10 min at 12000 g. One ml of the supernatant was reacted with 4 ml of 20% TCA solution comprising 0.6% thiobarbituric acid and then it was heated for 30 min at 95°C in a water-bath and then immediately cooled on ice. After centrifugation for 10 min at 12000 g, the absorbance of the supernatant was read at 532 and 600 nm respectively on a UV-VIS Spectrophotometer. The contents of MDA were worked out using the extinction coefficient of 155 mM-1 cm-1 using the formula [16]:
MDA level (nmol)= D A (532- 600) nm/ 1.56 × 105
The percentage increase in lipid peroxidation was calculated to find out the resistant and salt sensitive cultivars.
Reagents: i. 0.1% TCA-Dissolved 0.1 g TCA in 100 ml distilled water.
ii. 20% TCA+0.6% TBA (Thiobarbituric acid)-Dissolved 20 g TCA in 100 ml distilled water with 0.6 g TBA.
Total antioxidant activity
The antioxidant activities of the wheat extracts were measured using the stable DPPH (1,1-diphenyl-2-picrylhydrazyl) radical according to the method [17]. The alcoholic solution of DPPH radical (0.5 ml, 0.2 mM) was added to the 100 μl of sample solution and the mixture was shaken vigorously to left stand for 30 min in the dark. The absorbance was measured at 517 nm and the capacity to scavenge the DPPH radical was calculated using the following equation:
(%)=[(A0-A1)/A0] ×100
Where, Ao is the absorbance of the control reaction and A1 is the absorbance of the sample itself. The inhibitory concentrations at 50% (IC50) values (extract concentration that cause 50% scavenging of DPPH radical) were determined. All determinations were carried out in triplicate.
Reagents: 0.2 mM DPPH-Dissolved 2.24 mg DPPH in 25 ml ethanol.
RNA isolation and cDNA synthesis
Root and shoot sample (200 mg) was collected from plants of both the contrasting wheat genotypes treated as control (0 mM NaCl) and exposed to salt stress using 150 mM NaCl against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ) for 14 days. The samples were wrapped in aluminum foil, frozen immediately in liquid nitrogen and stored at -80°C until used for RNA extraction in batches. Total RNA was isolated in triplicate from the collected samples using RNA isolation Nucleospin RNA plant kit (Machery-Nagel; Cat.#740949.50) following the manufacturer's instructions. To remove contaminating DNA, isolated RNA (10 μg) samples were treated with RNase-free DNase I (Fermentas; Cat.#EN0525) according to the manufacturer's protocol. Integrity of the total RNAof shoot and root regions (Figure 1) was checked by denaturing agarose (1.0%) gel electrophoresis, and RNA concentration (Table 2) was estimated with the help of Nano-drop. Only those RNA samples showing 260/280 ratio in the range of 1.9-2.2 and 260/230 ratio>2.0 were used for cDNA synthesis. First-strand cDNA was synthesized from 0.5 μg of total RNA using Revert Aid Premium first strand cDNA synthesis kit (Fermentas; as per the manufacturer’s instructions.
| Sno. | Sample | shoot | root | ||
|---|---|---|---|---|---|
| Concentration (ng/µl) | Purity (260/280) nm | Concentration (ng/µl) | Purity (260/280) nm | ||
| 1 | UWC | 293.1 | 21.7 | 244.4 | 2.17 |
| 2 | PWSAC | 479.8 | 2.16 | 191.7 | 2.15 |
| 3 | PWMJC | 221.2 | 2.19 | 169.4 | 2.15 |
| 4 | UKC | 214.2 | 2.14 | 153.8 | 2.15 |
| 5 | PKSAC | 361.4 | 2.17 | 79.7 | 2.15 |
| 6 | PKMJC | 293.1 | 2.15 | 131 | 2.13 |
| 7 | UWT | 411.2 | 2.16 | 214 | 2.15 |
| 8 | PWSAT | 244.2 | 2.13 | 238 | 2.16 |
| 9 | PWMJT | 349.8 | 2.2 | 243 | 2.17 |
| 10 | UKT | 478.4 | 2.15 | 131 | 2.18 |
| 11 | PKSAT | 245.7 | 2.15 | 201 | 2.15 |
| 12 | PKMJT | 486 | 2.17 | 241 | 2.15 |
Table 2: Total RNA quantification of shoot and root region of contrasting wheat genotype-WH 542 and KRL 210 on 14 days after treatment of NaCl by using Nano Drop.

Figure 1: Total RNA from (a) shoot and (b) root tissue of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant), 14 days after salt (150 mM NaCl) treatment.
Semi-quantitative amplification of candidate genes (RTPCR)
The first strand cDNA (2.0 μl) was used as template for amplification of candidate HKT genes using gene specific primer pairs designed based on the conserved region of HKT genes in barley, oat, maize and rice having the sequence information available in the NCBI database. Two sets of primer pairs (Table 3) for different HKT genes were tested for their transferability, and two genes namely HKT1 and HKT3 (forward: 5'-TGGGATGCCACCAAAGAC-3'; reverse: 5'- TGATACGCAAATGTTGAGC-3' and 5'- TGGGATGCCACCAAAGAC-3'; reverse: 5'- TGATACGCAAATGTTGAGC-3', respectively) were amplified. PCR conditions were set as initial denaturation at 94°C for 3 min, followed by 39 cycles each at 94°C for 1 min, 58°C for 1 min, 72°C for 1 min, and a final extension at 72°C for 7 min. An aliquot of the PCR product was analyzed on 1% agarose gel for its expected size and DNA concentration. The constitutively expressed actin gene (UniGene cluster Ta54825; forward primer: 5'-TGGGATGCCACCAAAGAC-3'; reverse primer: 5'-TGATACGCAAATGTTGAGC-3') was used as reference gene in the semi-quantitative RT-PCR analyses. The RT-PCR experiment was repeated with three biological trials and three technical replicates.
| HKT genes | Old name | Probes / Primer |
|---|---|---|
| HKT2;1 | HKT1 | F1-TATGTGATGAGTCGCAGCTTGAA |
| R1-GCAACAAGAGGCCTGAATTCTTT | ||
| HKT2;3 | HKT3 | F2-TCTTAGTTCGGCAAGGCATATCA |
| R2-TGCACGGTAACCGATGTAACTCT | ||
| WheatActin | Ta 54825 | F-GTGCCCATTTACGAAGGATA |
| R-GAAGACTCCATGCCGATCAT |
Table 3: Probes/ Primersused for HKT gene expressions in contrasting wheat genotypes.
Real-Time/ quantitative amplification (RT-qPCR)
The quantitative gene expression analysis of the HKT genes-HKT1 and HKT3 was carried out according to guidelines [18]. Three biological replications, resulting from three different RNA extractions, RT and RT-qPCR reactions, and three technical replicates for each biological replication were used for quantitative analysis. First-strand cDNA was synthesized from 0.5 μg of total RNA, diluted 5-fold before using in RT-qPCR of 20 μl reaction volume containing 1.0 μl of cDNA and 10 picomoles forward and reverse primers. The RT-qPCR was performed using an Mx3000PTM real time PCR system (CFX-96, BioRad Platform) with SYBR Green qPCR Master Mix kit (BioRad; Cat.#170-8880AP). PCR amplification was carried out with an initial denaturation at 95°C for 3 min, followed by 40 cycles each of 30s denaturation at 94°C, 30s annealing at 58°C and 45s extension at 72°C. Amplification data collection was set at the end of each extension step. Preliminary qPCR assays was carried out with 10 picomole of primers using three different concentrations of cDNAs to generate lowest Ct value. Actin gene was used as reference gene. The Pfaffl formula for all the samples was used to calculate the relative expression of HKT1 and HKT3 genes under 150 mM NaCl salt stress on 14 days of treatment.
The biotic elicitors/ priming reagents-Salicylic Acid (SA) and Methyl Jasmonate (MJ) have been identified as vital cellular regulators to mediate diverse developmental processes and defense responses against biotic and abiotic stresses. The effect of these two biotic elicitor treatment/priming was investigated on the basis of morphological variations, Total Phenolic Content (TPC), antioxidant activity (AO), Lipid Peroxidation (LP) and high affinity potassium ion transporter (HKT) gene specific expressions in the shoot and root regions of two contrasting wheat genotypes (WH 542 and KRL 210) on 14 Days of salt stress imposition (150 mM NaCl) under controlled conditions.
Morphological expressions
The morphological expressions of salt-susceptible (WH 542) and salt-tolerant (KRL 210) wheat genotypes were influenced by priming effect of biotic elicitors-MJ and SA respectively (Figures 2 and 3). Primed seeds of both the contrasting wheat genotypes were showing healthier growth parameters as compared to the unprimed seeds. The comparison of unprimed control seeds i.e., without salt-stress and with salt-stress imposition indicated deterioration in the growth pattern and it was more pronounced in the salt-susceptible wheat genotype than that in the salt-tolerant one. The effect of priming with biotic elicitors provided initial rescue against the salt-stress in both the contrasting wheat genotypes (Figure 4). The effect of priming with respect to improved morphological growth of salt-susceptible wheat genotype than the salt-tolerant one indicated that this technology can be used in combating salt-stress and improved salt tolerance capacity of crop plants for better adaptation.
Figure 2: Differential morphological expression in WH 542 (saltsusceptible) wheat genotype with (a) methyl jasmonate and (b) Salicylic Acid (SA) priming after 14 days of 150 mM salt stress.

Figure 3: Differential morphological expressions in KRL 210 (salttolerant) wheat genotype with (a) methyl jasmonate and (b) Salicylic Acid (SA) priming after 14 days of 150 mM salt stress.
Figure 4: Comparative morphological expressions in WH 542 (saltsusceptible) and KRL 210 (salt-tolerant) wheat genotype with (a) Methyl Jasmonate (MJ) and (b) salicylic acid priming after 14 days of 150 mM salt stress.
Total Phenolic Content
The evaluation of Total Phenolic Content in contrasting wheat genotypes-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt-stress (150 mM) condition against priming of seeds with two biotic elicitors-salicylic acids (SA) and Methyl Jasmonate (MJ) was carried out. In both shoot (Figure 5) and root (Figure 6) regions of contrasting wheat genotypes under salt-stress conditions, Total Phenolic Content was found more in treated samples against priming with biotic elicitors. Tolerant wheat genotype (KRL 210) primed with methyl jasmonate (PKMJ) showed significantly higher phenolic content due to salt-stress imposition both in root and shoot regions. Root region showed higher total phenolic contents as compared to shoot region either primed with SA or MJ (Figure 7). Comparative analysis showed that the Total Phenolic Content was more accumulated in shoot and root regions of primed samples than that in unprimed ones.
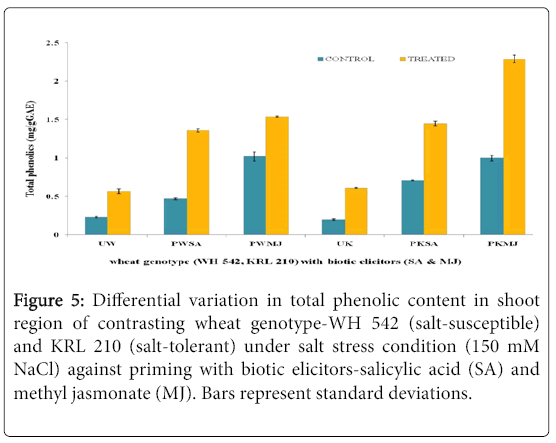
Figure 5: Differential variation in Total Phenolic Content in shoot region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). Bars represent standard deviations.
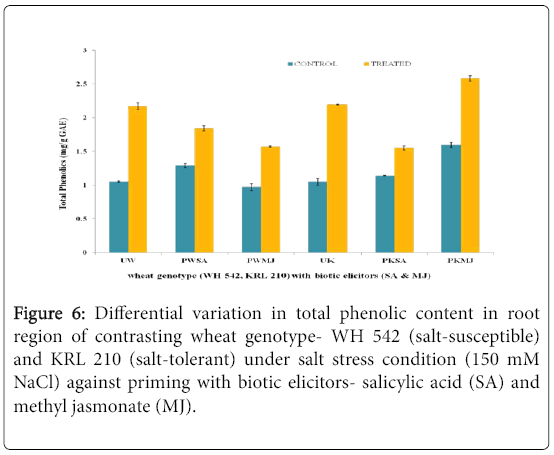
Figure 6: Differential variation in Total Phenolic Content in root region of contrasting wheat genotype- WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors- Salicylic Acid (SA) and Methyl Jasmonate (MJ).
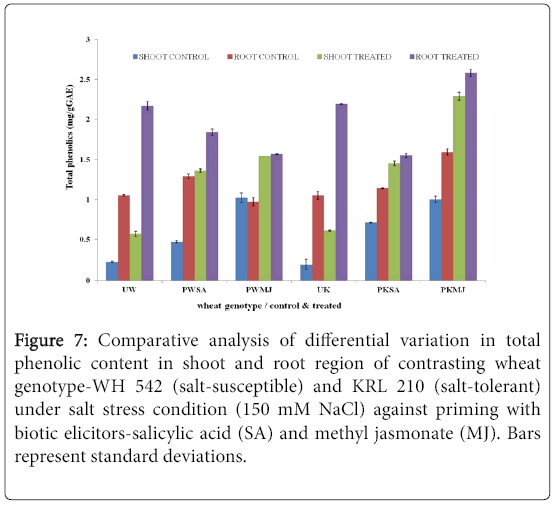
Figure 7: Comparative analysis of differential variation in Total Phenolic Content in shoot and root region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). Bars represent standard deviations.
Lipid peroxidation
The impact of salt stress (150 mM) on shoot (Figure 8) and root (Figure 9) region of contrasting wheat genotypes-WH 542 (saltsusceptible) and KRL 210 (salt-tolerant), primed with biotic elicitors (SA and MJ) revealed that lipid peroxidation was differentially expressed. Unprimed salt-susceptible wheat genotype exhibited higher level of lipid peroxidation under salt-stressed condition than the control one while in the unprimed salt-tolerant wheat genotype, reverse pattern was observed. Priming with both the elicitors lowered the lipid peroxidation level in salt-susceptible wheat genotype. In the case of SA-primed salt-tolerant wheat genotype, 50% reduced level of lipid peroxidation was found, while MJ-primed seeds of salt-tolerant genotype showed around 10 fold increase in the shoot region (Figure 8). It was indicated that SA and MJ were antagonizing the effect of each other.
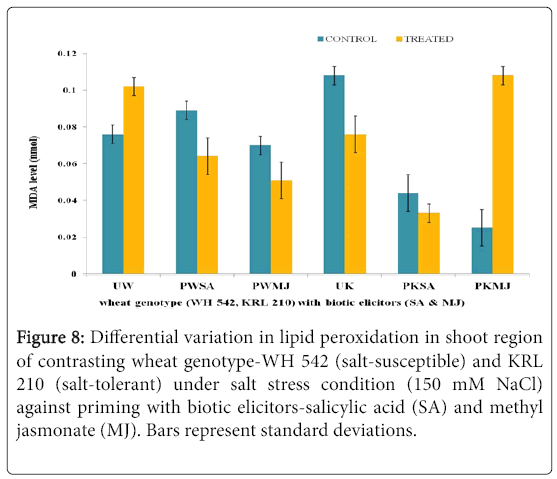
Figure 8: Differential variation in lipid peroxidation in shoot region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). Bars represent standard deviations.
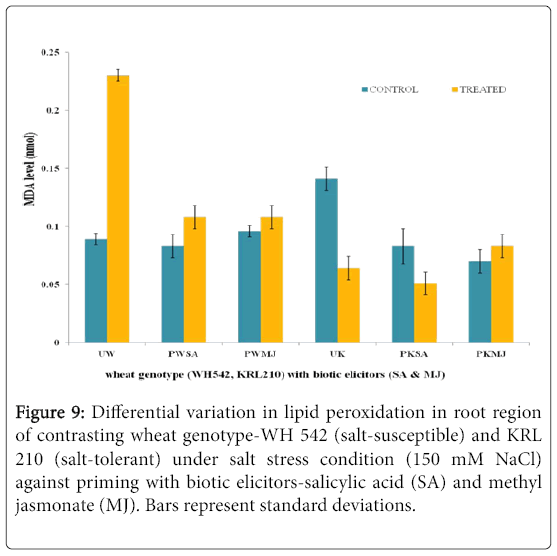
Figure 9: Differential variation in lipid peroxidation in root region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). Bars represent standard deviations.
In both shoot and root regions of contrasting wheat genotypes under salt stress conditions, lipid peroxidation was found more in primed treated samples. The salt-tolerant wheat genotype (KRL 210) primed with methyl jasmonate (PKMJ) showed significantly higher lipid peroxidation due to salt-stress imposition both in root and shoot regions. Root region showed higher LP as compared to shoot region either primed with SA or MJ. Comparative analysis showed that the LP was more in primed samples than the unprimed ones in both shoot and root regions (Figure 10).
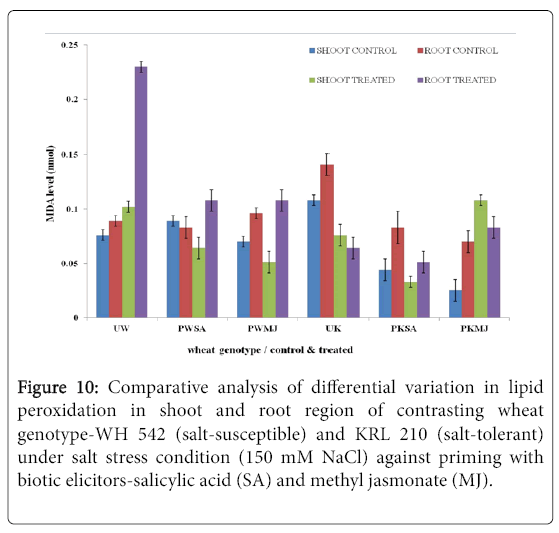
Figure 10: Comparative analysis of differential variation in lipid peroxidation in shoot and root region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ).
Total antioxidant activity
The antioxidant potential of shoot (Figure 11) and root (Figure 12) regions of contrasting wheat genotypes-WH 542 and KRL 210 revealed that scavenging percentage was higher in salt treated than control samples. The salt-susceptible wheat genotype-WH 542 showed more antioxidant activity as compared to the salt-tolerant wheat genotype. The seed primed plant samples were found to show more antioxidant activity under salt-stressed condition than that to salt-unstressed one. The biotic elicitor-MJ imparted more influence than the SA by increasing antioxidant activity to combat salt-stress. Roots exhibited more antioxidant activity compared to shoot in both the contrasting wheat genotypes (Figure 13) indicated it’s primary and direct involvement in the defense mechanism of plants.
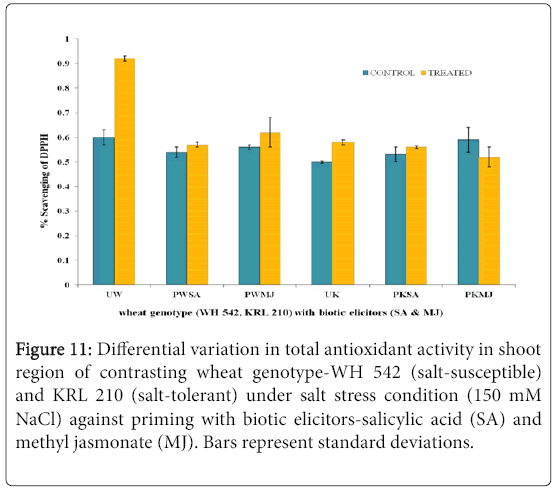
Figure 11: Differential variation in total antioxidant activity in shoot region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). Bars represent standard deviations.
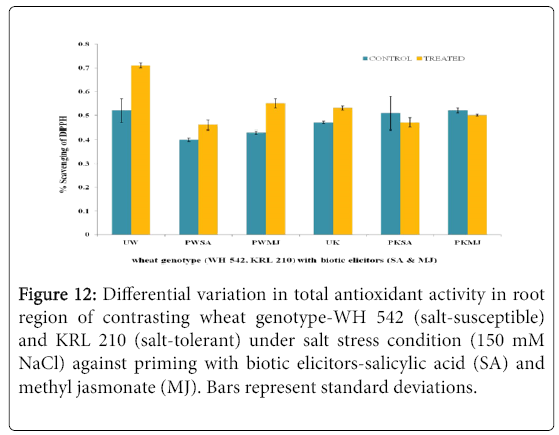
Figure 12: Differential variation in total antioxidant activity in root region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). Bars represent standard deviations.
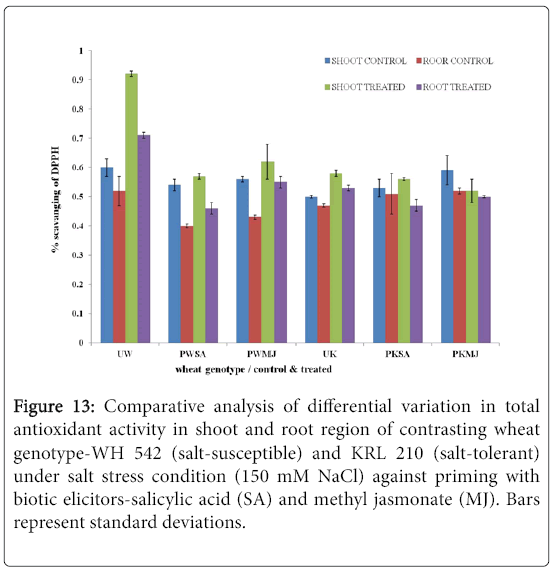
Figure 13: Comparative analysis of differential variation in total antioxidant activity in shoot and root region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) under salt stress condition (150 mM NaCl) against priming with biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). Bars represent standard deviations.
Expression analysis of high affinity potassium transporters semi-quantitative expressions (RT-PCR)
Based on HKT gene-specific primer pairs (Table 2), two HKT genes were amplified from shoot and root regions (Figures 14 and 15) of the two contrasting bread wheat genotypes-WH 542 and KRL 210 in all the samples as given in Table 1 respectively. One of the PCR products amplified with HKT1 gene-specific primers was observed to be of ~0.3 Kb, while another one with HKT3 gene specific one was of ~0.4 Kb molecular size. HKT genes-specific semi-quantitative RT-PCR analysis showed that the two HKT genes were differentially expressed in these contrasting wheat genotypes. HKT1 and HKT3 genes showed contrasting expression pattern in the bread wheat genotypes at 150 mM NaCl salt-stress imposition. The HKT genes were up-regulated in salt-susceptible wheat genotype while down-regulated in salt-tolerant wheat genotype. There was a differential variation in their expressions in root as well as shoot regions with reference to the induced effects of biotic elicitors (SA and MJ). The difference in expression level was found to be more prominent in root than in shoot after 14 days of salt treatment. As expected, expression level of the reference (actin) gene was observed to be constant throughout the samples of both the contrasting wheat genotypes.

Figure 14: Expression analysis of HKT1 and HKT3 genes by semiquantitative (RT-PCR) in shoot region of contrasting wheat genotype-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ) with respect to the actin (reference) gene expression.

Figure 15: Expression analysis of HKT1 and HKT 3 genes by semiquantitative (RT-PCR) in root region of contrasting wheat genotype- WH 542 (salt-susceptible) and KRL 210 (salt-tolerant) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ) with respect to the actin (reference) gene expression.
Real-time/ Quantitative expressions (RTqPCR)
Quantitative difference in expression level of HKT genes was further validated by Real-time/Quantitative PCR (RT-qPCR). Expression of both HKT1 and HKT3 genes was found to be up-regulated in the shoot samples of salt-susceptibe (Figure 16) and salt-tolerant (Figure 17) wheat genotypes. However, HKT3 expression in SA-primed saltsusceptible control-PWSAC (Figure 16) and HKT1 expression in unprimed salt-tolerant control-UKC sample (Figure 17) was found to be down-regulated. Figure 18 showed a comparative account of both the HKT genes (HKT1 and HKT3) in shoot region of both contrasting wheat genotypes at a glance. Quantitative expression analysis of HKT genes in the root region of all the samples of salt-susceptible wheat genotype (Figure 19) revealed that seed priming with SA was found to induce the up-regulation of both the HKT genes while MJ seed priming was down regulating the expressions. HKT3 gene was about 1.5 folds more down-regulated as compared to HKT1 in MJ-primed salt-susceptible root samples, while not much variation was found in the up-regulated expressions of both HKT1 and HKT3 genes in SAprimed ones. Unprimed salt-susceptible control and treated ones showed contrasting pattern with reference to HKT1 and HKT3 gene expressions (Figure 19). Expression of both HKT genes in the root region of unprimed control of salt-tolerant wheat genotype (KRL 210) was found to be down-regulated, while unprimed salt-tolerant control (UKC) and SA-primed salt tolerant control (PKSAC) samples showed progressive down regulation i.e., maximum in UKC and minimum in PKSAC and expression of HKT3 gene was more prominent than the HKT1 one. In the case of SA-primed salt-tolerant stressed-(PKSAT) sample, HKT1 was down-regulated, while HKT3 was up-regulated. The MJ-primed control samples of root region of salt-tolerant wheat genotype-(PKMJC) showed up-regulation of both the HKT genes but not as significant as that of MJ-primed salt-treated PKMJT (Figure 20), and the pattern of HKT1 and HKT3 gene expression was contrasting to each other. The expression of HKT1 was 2/3 fold higher than the HKT3 gene expression. Figure 21 showed a comparative account of both the HKT genes in root region of both contrasting wheat genotypes at a glance. Both the HKT genes were found to be downregulated in the root samples of PWSAC, PWMJC, UKC, UKT, PKSAC, PKSAT; and HKT3 showed more prominent down regulation than that of HKT1 gene. Figure 22 showed comparative account of both HKTs (HKT1 and HKT3) in both root and shoot regions of the contrasting wheat genotypes (WH 542 and KRL 210) at a glance. Thus, it was inferred that down regulation of HKT genes in root region of salt-tolerant wheat genotype provided tolerance to salt-stress imposition and MJ-induced seed priming showed more influence in the down regulation of HKT gene expressions than that of SA-induced ones in the roots of both the contrasting wheat genotypes.
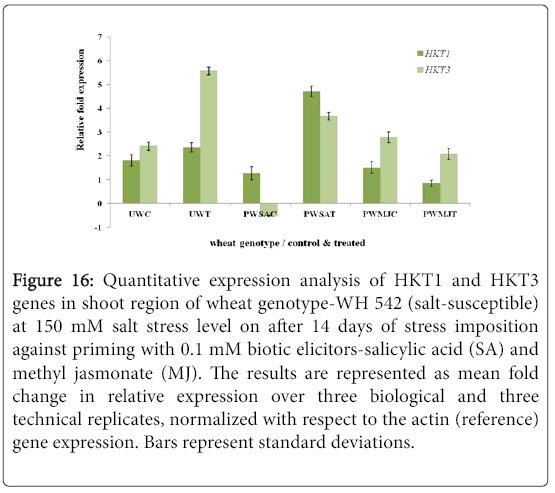
Figure 16: Quantitative expression analysis of HKT1 and HKT3 genes in shoot region of wheat genotype-WH 542 (salt-susceptible) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). The results are represented as mean fold change in relative expression over three biological and three technical replicates, normalized with respect to the actin (reference) gene expression. Bars represent standard deviations.
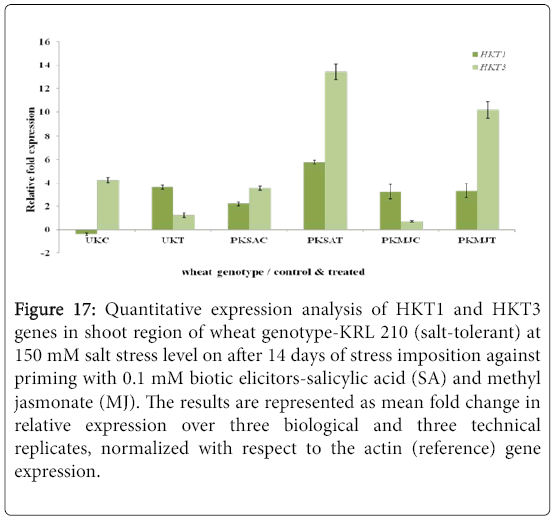
Figure 17: Quantitative expression analysis of HKT1 and HKT3 genes in shoot region of wheat genotype-KRL 210 (salt-tolerant) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). The results are represented as mean fold change in relative expression over three biological and three technical replicates, normalized with respect to the actin (reference) gene expression.
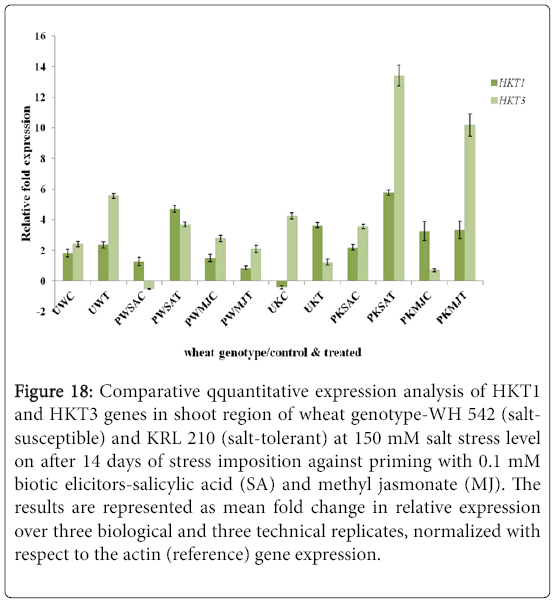
Figure 18: Comparative qquantitative expression analysis of HKT1 and HKT3 genes in shoot region of wheat genotype-WH 542 (saltsusceptible) and KRL 210 (salt-tolerant) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). The results are represented as mean fold change in relative expression over three biological and three technical replicates, normalized with respect to the actin (reference) gene expression.
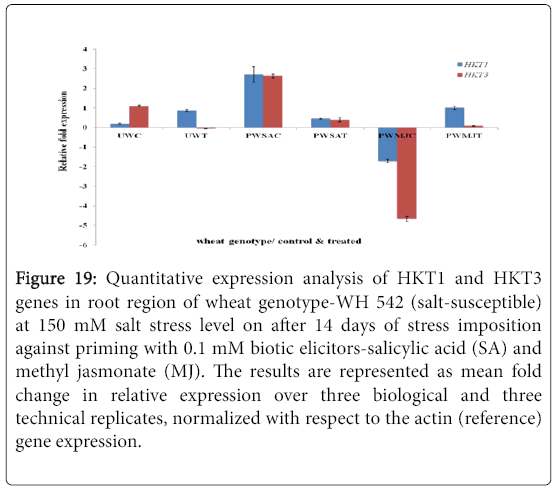
Figure 19: Quantitative expression analysis of HKT1 and HKT3 genes in root region of wheat genotype-WH 542 (salt-susceptible) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). The results are represented as mean fold change in relative expression over three biological and three technical replicates, normalized with respect to the actin (reference) gene expression.
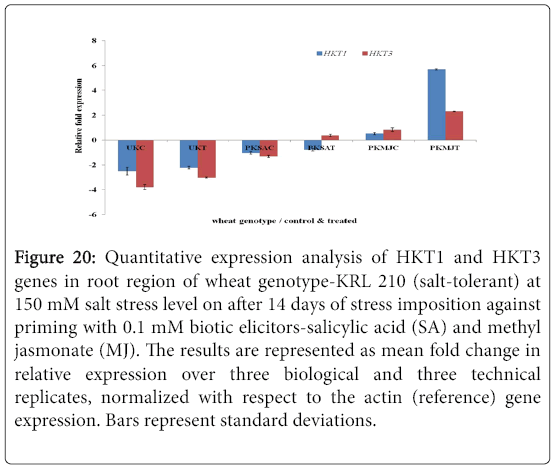
Figure 20: Quantitative expression analysis of HKT1 and HKT3 genes in root region of wheat genotype-KRL 210 (salt-tolerant) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). The results are represented as mean fold change in relative expression over three biological and three technical replicates, normalized with respect to the actin (reference) gene expression. Bars represent standard deviations.
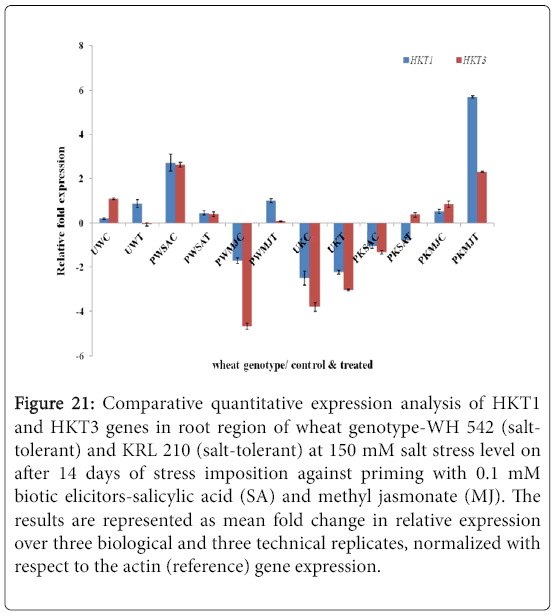
Figure 21: Comparative quantitative expression analysis of HKT1 and HKT3 genes in root region of wheat genotype-WH 542 (salttolerant) and KRL 210 (salt-tolerant) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). The results are represented as mean fold change in relative expression over three biological and three technical replicates, normalized with respect to the actin (reference) gene expression.
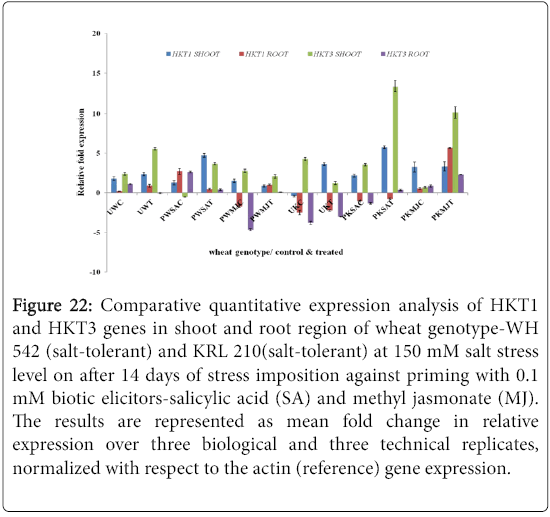
Figure 22: Comparative quantitative expression analysis of HKT1 and HKT3 genes in shoot and root region of wheat genotype-WH 542 (salt-tolerant) and KRL 210(salt-tolerant) at 150 mM salt stress level on after 14 days of stress imposition against priming with 0.1 mM biotic elicitors-Salicylic Acid (SA) and Methyl Jasmonate (MJ). The results are represented as mean fold change in relative expression over three biological and three technical replicates, normalized with respect to the actin (reference) gene expression.
Salt-stress produces a wide range of free radicals in whole plant and disturbs the most biochemical and metabolic reaction, and ultimately resulting in low grain yield or death of whole plant in severe conditions [19]. The results obtained in the present work clearly demonstrated that the two contrasting wheat genotypes (WH 542 and KRL 210) displayed distinct variation in salinity tolerance during vegetative growth. This reduction in growth might be due to toxicity of the ions or low osmotic potential as well as a decrease in wall extensibility [20,21]. Malondialdehyde (MDA) is a product of peroxidation of unsaturated fatty acids in phospholoipids, and the level of lipid peroxidation has been used as an indicator of free radical damage to cell membrane under stress conditions. MDA has been widely used to mark salt injury as membrane damage criterion in various plants [22]. Lipid peroxidation was found to occur in both chloroplast and mitochondrial [23] organelles of plant cell. Tolerance to NaCl-stress in higher plants correlates to the level of antioxidant system and substrate [24,25]. To overcome the effects of salinity induced oxidative stress, plants make use of a complex AntiOxidant (AO) system in strong correlation with accumulation of Total Phenolic Content (TPC) as osmolyte and Lipid Peroxidation (LP) as well. A positive correlation between TPCs and AntiOxidant (AO) activity (r=0.98) indicated [26] that AO activity is highly contributed by phenols [27]. Relatively higher antioxidant activities have been reported in tolerant genotypes when compared to susceptible ones, suggesting that the antioxidant system plays an important role in plant tolerance against abiotic stresses. Thus, genotypes responded differently as a result of variation in their antioxidant system [28,29]. The level of lipid peroxidation, measured as malondialdehyde (MDA) content, has been considered an indicator of salt-induced oxidation in cell membranes and a tool for determining salt tolerance in plants [30]. Lipid peroxidation rate was found to increase with increase of salt stress especially in sensitive cultivars [31,32]. Our results also agree thatsalt-tolerant (KRL 210) wheat genotype contain low level of MDA accumulation compare to salt-susceptible (WH 542) wheat genotype.
Concerted attempts have been made to mitigate the harmful effects of salinity by application of plant growth regulators [33]. Thus the detrimental effects of high salts on the early growth of wheat seedlings may be reduced to some extent by treating seeds with the proper concentration of a suitable hormone [34]. This injury is probablycaused by overloading the vacuolar capacity to compartmentalize toxic salt species. Alternatively, thesetoxic salt species might build up in cell wall and causedehydration [35]. Our results indicated differential variation in TPC, LP and AO activity in these contrasting wheat genotypes even at the unstressed i.e., control level, substantiating that these are genetically different in their constitution. The increase in Lipid Peroxidation (LP) level in saltstressed samples may be due to oxidative damage affecting both organelles (chloroplast and mitochondria). The lower LP level in saltstressed tolerant genotype than the un-stressed i.e., control is important in terms of salt tolerance in different studies [22,24,36], and confirming their tolerance to salinity stress compared to saltsusceptible genotypes. The possibility of LP/MDA the involvement in combating salt tolerance [37,38] has been reported and our findings are in accordance to these reports. Primmed crops grew more vigorously, flowered earlier, yielded higher [5,39] and showed beneficial effects on germination percentage [40]. In present study, saltstress caused significant retardation in physiological growth of contrasting wheat genotypes and in agreement with those obtained decrease in wheat germination due to salinity stress [3]. It may be due to reduced water potential, increased reactive oxidant potential and membrane damage i.e., lipid peroxidation and high accumulation of phenols, resulting slower rate of imbibition.
Seed priming is found to be one of the useful physiological approaches to mitigate the effect of salt-stress. In the present investigation, two biotic elicitor-Salicylic Acid (SA) and Methyl Jasmonate (MJ) were used for seed priming of contrasting wheat genotypes-WH 542 (salt-susceptible) and KRL 210 (salt-tolerant). It was quite clear that seeds primed with SA and methyl jasmonate MJ proved to be effective in inducing salt tolerance in wheat genotypes and are in accordance with reports related to SA-primed seeds [41]. Several studies supported a major role of SA and MJ in mediating the plant responses to abiotic stresses; however, the molecular events in their signaling pathway are not yet completely understood. Characterization of gene encoding jasmonic acid carboxymethyl transferase has provided information on the role (s) of phytohormone in gene activation control and systemic long-distance signaling [42]. Transcriptional results of cross talk between the SA and MJ pathways indicated their antagonism and synergism type of gene regulation in plant signaling responsive cascade [43]. Lipids found to be connected to the plant defense response through potential action as signaling molecules [44] through the fatty acid-linolenic acid, acting as the precursor for 12-OxyPhytoDienoic Acid (OPDA) and JA synthesis via octadecanoid pathway [45]. Concerted attempts have been made to mitigate the harmful effects of salinity by application of plant growth regulators [33]. Thus, the detrimental effects of high salts on the early growth of wheat seedlings may be reduced to some extent by treating seeds with the proper concentration of a suitable hormone [34]. The injury is probably caused by overloading the vacuolar capacity to compartmentalize toxic salt species. Alternatively, these toxic salt species might build up in cell wall and cause dehydration [35]. MJ primed seed plants exhibited more antioxidant activity than SA primed ones and salt-stressed showed [46] higher AntiOxidant (AO) activity than unstressed (control) ones.
Plant plasma membrane transporters are thought to be involved in Na+ homeostasis by extruding the ion from root epidermal cells at the root-soil interface and by regulating the root-shoot distribution of Na+ in coordination with HKT proteins [47]. Adverse effects of salinity on plant growth may be due to ion cytotoxicity and osmotic stress [48]. In many species, a large component of tolerance to long-term exposure to Na+ can be attributed to the ability of plants to exclude Na+ from the shoot. In wheat, one of the major mechanisms conferring salt tolerance is sodium exclusion from the leaves, where it is transported from root xylem via root specific transportes. Sodium exclusion is achieved through low net uptake by cells in the root cortex and tight control of net unloading of the xylem by parenchyma cells in the stele. Hexaploid bread wheat (Triticum aestivum , AABBDD) cultivars exhibited slow rates of Na+ transport to the shoot and maintain a high K+/Na+ ratio in leaves. This enhanced K+/Na+ discrimination trait confers some degree of salt tolerance. Wheat contains a large number of bioactive compounds also which may possess the scavenging capacity against free radicals and Na+ and Cl- ions. High affinity potassium transporters (HKTs) being integral membrane protein (IMPs) worked in a network mode to regulate ion homeostasis in the plant system. A new source of Na+ exclusion was found in durum wheat, Line 149, which maintained low Na+ concentrations and high K+/Na+ ratios in the leaf blade, quite similar to that of bread wheat [5]. In our observations, the expression level of transcripts of HKTs in the shoot region was found up-regulated while down-regulated in the roots of contrasting wheat genotypes. Up-regulation of these transporter genes in the salt-tolerant wheat genotype might be a response to more Na+ in the vacuoles and high K+/Na+ ratios in the leaf blade. Therefore, the capacity to maintain a low Na+ concentration or a high K+/Na+ ratio in the leaves may be considered as an indicator of potential salinity tolerance in the cereals.
The HKT family plays a major role in K+ uptake in plants by mediating Na+/K+ co-transport. Plants have an absolute requirement for potassium (K+), while sodium (Na+) is toxic for many biological reactions in the cytoplasm. On the contrary, in the vacuolar processes, replacement of K+ by Na+ does not produce toxicity and is regulated with the help of HKTs only. Most of the plants show a high degree of K+/Na+ discrimination for uptake and plant genome contain a high number of genes encoding potassium transporter and channels. Evidences using a root uptake system, supported that TaHKT2.1 functions as a Na+ uniport [49]. In durum wheat, the gene homologous to TmHKT7 which is the best candidate for Nax1, could control Na+ unloading from xylem in root and sheath of line 149 (salt tolerant), but not of salt sensitive (Tamaroi) variety.
With reference to the above research findings, we analyzed the expressions of two HKT genes in the shoot and root region of the contrasting bread wheat genotypes. Both the HKT1 and HKT3 belonging to HKT2; ‘y’ subfamily showed similar expression pattern in both the genotypes. Findings suggested that HKT genes are involved in Na+/ K+ transport and tolerant genotype may combat the salt stress by up regulated expression of its HKT genes in shoot and down regulated in the roots. This indicated that HKT transporters in the roots will be reducing the uptake of salt from the soil, while in the shoots they will be increasing the efflux/removal of salt ions through vacuoles. Seed priming has induced the salt tolerance capacity of salt-susceptible genotype at much higher level than the salt-tolerant genotype. These biotic elicitors (SA and MJ) were found to mitigate the effect of saltstress by affecting expression of HKT genes as well their biochemical mechanisms and detoxification pathways. MJ as abiotic elicitor showed more prominent response than that of SA and can be utilized to improve defense responses of crop plants against salt stress. However, there is a need to further analyze plant salt-stress response at molecular level due to complexity of events associated with the sensing of salt stress and the activation of specific pathways.
In order to achieve salt tolerance, firstly, damage must be prevented or lessened; secondly, homeostasis must be re-established in the stressed conditions; and thirdly, even if at a lowered rate, plant growth must be resumed following the modifications at the cellular and whole-plant levels. The toxic effects caused by sodium ion and osmotic homeostasis need to be quickly neutralized to evolve an efficient detoxification mechanisms. Seed priming-a double technology to enhance rapid and uniform emergence, and to achieve high vigour and better yields in saline areas can be utilized to overcome the deleterious effects due to salt-stress in crop plants. Finding shere indicated that Total Phenolic Content (TPC), Lipid Peroxidation (LP) and AntiOxidant (AO) activity are crucial biochemical markers for resistance against salt stress and it seems that the defense responses against salt-stress are often dependent on these. There was a progressive increase in TPC, LP and AO activity during salt stress imposition and increase was found to be of higher magnitude in the salt-susceptible genotype than that of the salt-tolerant one. Both HKT1 and HKT3 genes from HKT2 subfamily were involved in regulating ion homeostasis. Up-regulated HKT gene expressions in shoot and down regulation in root under salt-stress indicated that HKT transporters in the roots will be reducing the uptake of salt from the soil, while in the shoots they will be increasing the efflux/removal of salt ions through vacuoles. Seed priming has induced the salt tolerance capacity of salt-susceptible genotype at much higher level than the salt tolerant genotype. These biotic elicitors (SA and MJ) were found to mitigate the effect of salt-stress by affecting expression of HKT genes as well their biochemical mechanisms anddetoxification pathways. MJ as abiotic elicitor showed more prominent response than that of SA and can be utilized to improve defense responses of crop plants against salt stress. The great leaps and bounds towards understanding plant salt-stress responses and tolerance mechanisms have been achieved in the past years. However, many challenges still lie ahead and plant breeders confront serious challenge to develop superior cultivars with broad spectrum of abiotic resistance. The regulation of gene expressions and signaling cascades that regulate Na+ transporters remain to be elucidated. Thus, present findings will facilitate insight in unraveling the mechanism of ion homeostasis during salt-stress to improve crop yield.
All the research work was done at Division of Biochemistry, ICARIndian Agricultural Research Institute, New Delhi, India.