Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2015) Volume 3, Issue 3
Cassava (Manihot esculenta Crantz) is an important perennial tropical crop for millions of people globally, particularly in subSaharan Africa. It is preferred due to its agronomical attributes such as ability to grow in poor soils and drought resistance. Cassava storage roots are a good source of starch. Further, cassava leaves and shoots are eaten as vegetables and are an excellent source of vitamins, minerals and protein. Cassava is also used in the production of ethanol for fuel, animal feed, and as a raw material for the starch industry. Cassava has high photosynthetic rates and its roots can persist in the soil for 8-24 months without decaying, thereby making it an ideal food security crop. In Kenya, cassava is a major source of subsistence and cash income to farmers in agroclimatically-disadvantaged regions and high potential areas of coast, Central and Western regions of Kenya. Sadly, cassava leaves and roots contain potentially toxic levels of cyanogenic glycosides. Cassava is largely propagated clonally making it an ideal plant for improvement through genetic engineering. This study was designed to produce transgenic acyanogenic cassava plants in which the expression of the cytochrome P450 genes (CYP79D1/D2) is downregulated through RNAi. Three Kenyan cassava genotypes; Adhiambo lera, Kibanda meno and Serere along with an exotic model cultivar TMS 60444 were used. The transformable lines were then taken through Agrobacterium-mediated transformation with an RNAi cassette harbouring cytochrome P450 genes (CYP92D1) to down regulate production of cyanoglycosides. Molecular analysis by PCR and RT-PCR confirmed transformation of the putative transformants.
Keywords: Manihot esculenta; RNAi; PCR; RT-PCR; Southernblotting; Agrobacterium tumefasciens
Cassava (Manihot esculenta Crantz) is the fourth most important crop in the developing countries surpassed only by maize, rice and sugarcane as a source of calories [1]. In sub-Saharan Africa cassava provides up to 60% of the daily calorie uptake [2,3] and more than 80% of the harvest is used as food. In certain regions the leaves, which contain appreciable quantities of protein and vitamins, are used as a major component of the diet to provide supplementary protein, vitamins and minerals to complement the carbohydrate rich staple [4]. Cassava is a vital food security crop that helps the fight against hunger and poverty in developing countries and especially in Africa, where the food security problems are the most severe in the world [5]. Traditional breeding of cassava is difficult due to irregular flowering and low fertility as well as low seed set and germination rates of the plants, and attempts to improve the quality of cassava roots have so far been unsuccessful. Advances in plant genetic engineering now provide an alternative to traditional breeding in improving cassava. The recent development of transformation methods [6] will allow the addition of agriculturally valuable traits to cassava for improved root quality. Methods used to introduce DNA into plant species include transformation using Agrobacterium [7-9] and direct gene transfer methods such as protoplast transformation [10], electroporation [11,12], microinjection[13], and microprojectile bombardment [7,14]. Cassava was considered to be a recalcitrant species for genetic engineering [15-19]. However, recent studies have optimized successful transformation protocols for a wide variety of cassava genotypes [6]. This paper reports successful Agrobacterium-mediated transformation of regenerable Kenyan cassava varieties using immature leaf lobes as explants.
Plant material
Sterile in vitro cassava plants were initiated from immature stem cuttings obtained from 4 to 6 months-old growth room-grown plants. Stems were excised, and divided into pieces with two to three nodes that were then surface sterilized to arrest contamination. The stem pieces were cultured in MS shoot multiplication medium [20] supplemented with 20 g/L sucrose, MS vitamins (Duchefa, Germany) and 8 g/L of noble agar. Cultures were grown at 28°C under a 12-hour light/12- hour dark regime until young actively growing apical leaves emerged in about 4 weeks following culture. The genotype TMS 60444 was used as a control in transformation experiments.
Agrobacterium preparation
Agrobacterium strain LBA4404 harbouring the binary vector plasmid pCAMBIA 2301 with the RNAi construct of CYP79D1/D2 was streaked and grown for 2 days on solid YEP medium supplemented with 50 mg/L streptomycin and 50 mg/L kanamycin. Cultures were incubated at 28°C. After 2 days, single colonies were used to inoculate 20 mL of liquid YEP medium supplemented with 200 μM acetosyringone, 50 mg/L streptomycin, 50 mg/L kanamycin and incubated at 28°C on a shaker for 2 hours. Kanamycin resistance gene had been inserted in the transformation gene cassette between the left and right borders. Kanamycin serves as a marker for transformation. The bacteria were then pelleted by centrifugation (10,000 g) for 10 min, and resuspended in 20 mL of MS basal salt (co-culture) medium devoid of nitrogen, but supplemented with 1% (w/v) of glucose and 1% (w/v) of galactose, and the pH adjusted to 5.5. At this stage the bacteria were ready for inoculating the explants.
Explant inoculation and co-cultivation
Young actively growing apical leaves were excised from clean cultures, placed onto a sterile filter paper and wounded using a sterile needle. Wounded apical leaves were then placed with the adaxial surface on co-cultivation medium (MS basal medium supplemented with 2% (w/v) sucrose, B5 vitamins, 50 mg/L casein hydrolysate, 0.5 mg/L CuSO4 and 8 mg/L 2,4-dichlorophenoxyacetic acid supplemented with 200 μM acetosyringone). Two drops of the bacterial suspension in liquid MS (to which was added two drops of Tween-20) were applied to the explant tissue, which was then incubated in the dark for 2 days at 28°C for co-culture. Subsequently, the explants were transferred to a new co-cultivation medium for another 3 days of co-cultivation. Following co-cultivation with A. tumefaciens, explants were transferred to selection media (MS basal medium supplemented with 2% (w/v) sucrose, B5 vitamins, 50 mg/L casein hydrolysate, 0.5 mg/L CuSO4 and 8 mg/L 2,4-dichlorophenoxyacetic acid supplemented with 50 mg/L kanamycin and 500 mg/L of carbenicillin) to induce embryogenesis. For negative control, a separate set of explants was not co-cultivated with A. tumefaciens but was transferred into the selection media. The cultures were incubated on a 12 hr day/12 hr night cycle for 4 weeks. The cultures were maintained on selection medium for 4 weeks.
Selection and regeneration of putative transformants
Somatic embryos produced on selection media were transferred to selection shoot regeneration medium (MS basal medium with 1 mg/L thiamine, 100 mg/L myo-inositol, 2% (w/v) sucrose, 1.0 mg/L BAP and 0.5 mg/L GA3 at pH 5.7 supplemented with 50 mg/L kanamycin and 500 mg/L of carbenicillin) to induce the development of cotyledonary leaves. Survival of regenerants in the medium containing kanamycin is indicative of transgenicity because the transformation gene construct contained kanamycin resistance gene. Once the individual somatic embryos germinated and formed shoots they were transferred to cassava micropropagation medium (MS salts, 2% (w/v) sucrose and activated charcoal, pH 5.7) for plant recovery for four weeks. All the cultures were incubated at 28°C under a 12-hour light/12-hour dark regime. Regenerated putative transgenic plantlets were transferred to sterile peatmoss and grown in confinement within the glasshouse to adhere to biosafety rules under natural lighting conditions and a temperature range of 22-30°C. The transferred plants were covered with transparent plastic bags to maintain high humidity. The bags were removed after 2 weeks and the plants were transferred to normal soil to expose them to normal environment. The transformed plants were given the same water treatment as the wild-type cassava plants.
Molecular analysis of putative transgenic cassava lines
Polymerase chain reaction (PCR) analysis: Genomic DNA was isolated from leaves of three months old grown wild-type plants and kanamycin-resistant putative transformants. Wild-type plants used were not transformed but grown in tissue culture media (without selection) similar to the putative transformants. The presence of the CYP79D1/D2 gene was detected by PCR amplification. Polymerase Chain Reaction (PCR) reactions were performed in a total volume of 25 μL containing: 1X PCR buffer, 20-100 ng of leaf DNA, 0.1 mM each dNTP, 1 unit Taq polymerase, 0.4 μM each primer. The CYP79D1/D2 gene was amplified using forward (5’-TGGCCATGAACGTCTCCACC-3’) and reverse (5’-TGGAGCCATTTGTGTCTTGC-3’) primers. The PCR amplification profile was as follows; 3 minutes at 94°C, 30 seconds at 94°C, and 45seconds at 58°C and 30 seconds at 72°C for 30 cycles. The PCR product was run on 1% (w/v) agarose gel at 70 V in 0.5X TBE buffer. Control experiments were carried out with DNA from the respective wild type cultivars.
Reverse transcriptase-PCR (RT-PCR) analysis: This was done to determine the abundance of transgene insertion in the putative transgenics. Total RNA was extracted from 100 mg of in vitro putative transformed root material using a Qiagen Plant RNA Extraction Kit (Qiagen Inc., Valencia, CA). The extract was treated with 1 unit DNase (Invitrogen, Carlsbad, CA) for 15 minutes at room temperature to eliminate DNA contamination. The DNase was inactivated according to manufacturer’s instructions. The first strand cDNA synthesis was carried out with 10 μg of total RNA using 1X reverse transcription buffer (50 mM Tris-HCl pH 8.0, 75 mM KCl, 3 mM MgCl2) 0.3 mM dNTP, 0.5 μg oligodT(12-18) primers, and 200 units of SuperScript II reverse transcriptase (Life Technologies, Rockville, MD). The mixture was incubated at 65°C for 5 minutes without reverse transcriptase followed by incubation at 42°C for 1 hour with reverse transcriptase. The synthesized cDNA was amplified by PCR using 1X PCR buffer, 1.5 mM MgCl2, 0.1 mM dNTP, 2.5 units of Taq polymerase, 0.4 μM of each primer specific for CYP79D1 gene. The forward primer sequence is 5’-TGGCCATGAACGTCTCCACC-3’ and the reverse primer sequence is 5’-TGGAGCCATTTGTGTCTTGC-3’. The PCR conditions were as follows: 3 minutes at 94°C, 30 seconds at 94°C, 45 seconds at 58°C and 30 seconds at 72°C for 30 cycles. The PCR product was run on 1% (w/v) agarose gel at 70 V in 0.5X TBE buffer. Control experiments were carried out without cDNA (negative control) and by amplification of the CYP79D1 gene. A separate PCR was conducted using primers specific to cassava starch branching enzyme- II (SBE-II) gene to serve as internal control.
Southern blot analysis of the transgenic plants: Genomic DNA was isolated from leaves of 3 months old grown wild-type plants and kanamycin-resistant putative transgenics using a protocol adapted from [21]. Upon DNA isolation, Southern blotting was undertaken following a standard protocol adapted from [22]. Approximately 10 μg of genomic DNA was digested with Kpn1 followed by fractionation on a 0.8% agarose gel. The fractionating gel was then soaked in HCl (250 mM) for 12 minutes, denaturation buffer comprising 1.5M NaCl and 0.5M NaOH for 50 minutes and neutralization buffer comprising 1M Tris-HCl and 1.5M NaCl for 35 minutes. Constant gentle agitation accompanied each soaking stage following, there was a short-lived rinsing with distilled water. The DNA was then transferred to nitrocellulose membrane for 20 hours. Transferred DNA was cross-linked to the membrane by UV light exposure for 40 seconds. Membrane prehybridization was done for a minimum of 1 hour, 45 minutes at 40°C in CHURCH buffer (0.5M NaHPO4 (pH 7.2), 1 mM EDTA, 1% (w/v) BSA, 7% (w/v) SDS). Hybridization of the membrane was conducted at 40°C with biotin-labeled CYP79D1 probe for 20 hours. The probe was made using PCR primers specific to CYP79D1 and the PCR reaction was performed using 1X PCR buffer, 1.5 mM MgCl2, 0.1 mM dNTP, 2.5 units of Taq polymerase, 0.4 μM of each primer specific for CYP79D1 gene. The biotin-labeled probe was then cleaned using Qiagen PCR cleaning kit (Qiagen Inc, Valencia, CA) and boiled for 5 minutes prior to addition to the membrane. Following hybridization, the membrane was washed with 1X Saline-Sodium Citrate (SSC) containing 0.1% (w/v) SDS at 45°C for 20 minutes followed by two washes with 0.1X SSC buffer containing 0.1% (w/v) SDS each at 45°C for 20 minutes. The membrane was then exposed toan imager overnight.
The data on regeneration of the putative transformants was analysed in terms of regeneration frequency, regeneration efficiency and transformation frequency. Regeneration frequency was calculated as the number of germinated shoots divided by the total number of recovered calli events in germination medium (GM). Regeneration efficiency was calculated as the total number of regenerated shoots divided by the total number of infected explants. Transformation frequency was calculated as the total number of recovered calli events on GM divided by the total number of infected explants. Means of the number of explants, live calli on selection media, live calli on germination media and regenerated shoots were also computed. Minitab statistical software (version 2012) was used for data analysis.
Agrobacterium-mediated transformation and regeneration of cassava genotypes
Four cassava cultivars were transformed via Agrobacteriummediated gene transfer. Transformed cells underwent somatic embryogenesis in 4-6 weeks on embryogenesis medium supplemented with 50 mg/L kanamycin and 500 mg/L carbenicillin. Some embryogenic calli kanamycin survived selection, although some portions of embryogenic calli were dying (Figure 1). Kanamycin resistance indicates that most tissues that regenerate are transgenic. Kanamycinresistant embryos converted to plantlets in 1-2 months. Plantlets were then transferred to individual test culture bottles after 1 month. The individual plants in culture bottles were grown for 2 weeks and were then transferred to the greenhouse and later to the glass house (Figure 2). The negative control explants that were not agro-co-cultivated but were transferred to selection media, did not survive and they all died. The trend of transformation of the regenerable cassava cultivars was genotype dependent and consistent with regeneration data. Adhiambo lera showed the best response in terms of transformation frequency, regeneration frequency and regeneration efficiency, followed by Serere, TMS 60444and Kibanda meno (Table 1). There was regeneration of the transformed cultivars for all varieties.
| Cultivar | No. of explants | SM live calli | GM calli | Shoots | RF% | TF% | RE% |
|---|---|---|---|---|---|---|---|
| TMS 60444 Mean ± SEM |
126 29.9 ± 1.4 |
67 19.4 ± 1.1 |
38 5.8 ± 1.3 |
26 5.2 ± 0.5 |
38.81 | 30.16 | 2.06 |
| Adhiambo lera Mean ± SEM |
119 33.2 ± 1.1 |
80 23.4 ± 1.6 |
53 12.3 ± 0.6 |
39 8.7 ± 0.3 |
48.75 | 44.54 | 3.28 |
| Kibanda meno Mean ± SEM |
121 24.2 ± 1.7 |
62 15.3 ± 1.4 |
34 6.7 ± 1.1 |
29 4.0 ± 0.7 |
46.77 | 28.10 | 2.40 |
| Serere Mean ± SEM |
124 44.6 ± 1.4 |
71 18.3 ± 1.2 |
45 9.4 ± 0.8 |
32 4.2 ± 0.4 |
45.07 | 36.29 | 2.68 |
RF (%), Regeneration Frequency=Number of shoots germinated over total number of recovered calli events in germination medium (GM).
TF (%), Transformation Frequency=Total number of recovered calli events on GM over total of infected explants.
RE (%), Regeneration Efficiency=Total number of shoots regenerated over total number of explants infected.
Table 1: Transformation, callus recovery and regeneration of cassava lines.
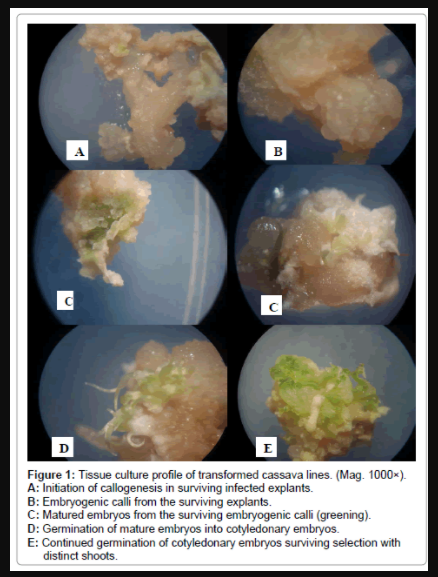
Figure 1: Tissue culture profile of transformed cassava lines. (Mag. 1000×).
A: Initiation of callogenesis in surviving infected explants.
B: Embryogenic calli from the surviving explants.
C: Matured embryos from the surviving embryogenic calli (greening).
D: Germination of mature embryos into cotyledonary embryos.
E: Continued germination of cotyledonary embryos surviving selection with distinct shoots.

Figure 2: Transgenic cassava lines at different stages of growth.
A: Young transgenic cassava plantlet growing in soil in glasshouse immediately after transfer from culture bottles.
B: Mature transgenic cassava plants continuing growth in glasshouse. C: Fully mature transgenic cassava plants in bio-containment field. Mature refers cassava plants growing but not to the level of producing tubers. Fully mature refers to cassava plants grown to the level of producing tubers.
Polymerase chain reaction (PCR) analysis for the T-DNA integration
The kanamycin-resistant putants were screened by PCR for the CYP79D1 gene. Each putant was obtained from a unique explant and not from a secondary somatic embryo. Similarly, untransformed wild-type plants used in all analyses were generated from independent explants via the same tissue culture procedures used to generate putative transformants. Twelve PCR-screened putants were positive for the CYP79D1/D2 genes (Figure 3).

Figure 3: PCR analysis of putative cassava transformants. Given below are three independent transgenic events for: TMS 1, TMS 2, TMS 3-TMS 60444 AL 1, AL 2, AL 3-Adhiambo Lera; Kb 1, Kb 2, Kb 3 -Kibanda Meno; Ser 1, Ser 2, Ser 3 -Serere.
Reverse transcriptase-polymerase chain reaction (RT-PCR) analysis
The cDNA from transformed plants had negative and reduced amplification of a CYP79D1/D2 gene reverse transcriptase product compared with the wild-types. Three transgenic plants (Kb 2, Ser 2 and Ser 3) had reduced levels of the CYP79D1/D2 transcripts. Conversely, nine transgenics (TMS 1, TMS 2, TMS 3, AL 1, AL 2, AL 3, Kb 1, Kb 3 and Ser 1) seldom had any detectable CYP79D1/D2 transcripts. Polymerase chain reaction (PCR) of the internal control was run in parallel (Figure 4).
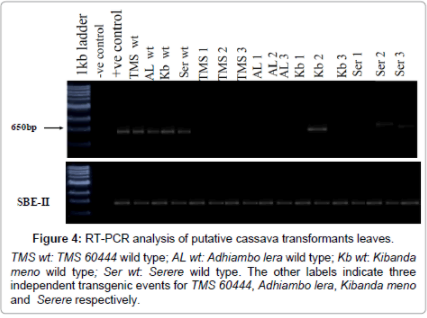
Figure 4: RT-PCR analysis of putative cassava transformants leaves.
Southern blot analysis
Southern blot analysis was undertaken to confirm the PCR and RT-PCR analysis results for transgene integration. Genomic DNA of twelve putative transgenics along with one wild type cultivar were restricted with Kpn1 and probed with biotin-labeled CYP79D1/D2. As (Figure 4) shows, the copy numbers of the inserted transgene ranged from one to three. TMS 2 and Ser 3 had one copy number of transgene insertion. TMS 1, TMS 3, AL 1, AL 2, Kb 2, Ser 1 and Ser 2 had two copy numbers of transgene insertion. Three copy numbers of transgene insertion occurred in AL 3, Kb 1 and Kb 3 (Figure 5).
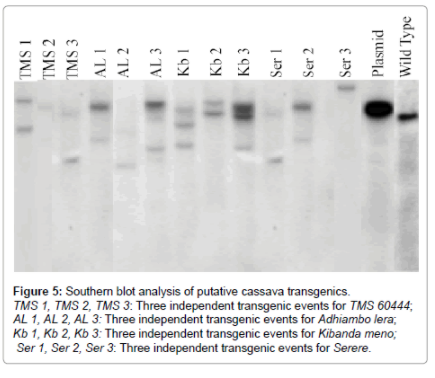
Figure 5: Southern blot analysis of putative cassava transgenics.
Successful transformation and regeneration of four cassava lines and a model line, was demonstrated in this study. The genotype TMS 60444 was used as model cultivar while the others were Kenyan cassava lines. The study results indicate that immature leaf lobes are also good explants for transformation of cassava. Friable embryonic calli (FEC) and germinated somatic embryos have previously been used as explants in cassava transformation [7,23-25]. The transformation and regeneration frequencies of the transformed cassava lines were genotype dependent and consistent. Similar findings were observed by [25] who reported genotypic related transformation and regeneration frequencies and efficiencies of cassava genotypes TME 13, TME 127, TME 8, TME 1, TMS I 91/02327 and TMS 60444. Agrobacterium strain LBA4404 harbouring pCAMBIA 2301 was used for transformation cassava in this study. This has previously been found to be more efficient in T-DNA transfer than other strains. Previous reports describing cassava transformation have shown that the Agrobacterium strain LBA4404 was suitable for the transformation of the cassava genotype MCol 22 [15] due to the presence of duplicated Vir D region on the co-integrated plasmid pTOK233 of LBA4404 that enhanced the virulence of such Agrobacterium strains in rice [26] and cassava [1]. This may account for the slightly higher transformation frequencies and efficiencies observed in this study. In embryogenic callus of cotton, LBA4404 was also shown to be more efficient in producing transformants than C58C3 [27]. In this study, LBA4404 harbors pCambia2301 instead of pDgusbin19 in the work of [27]. This, therefore, demonstrates that the superiority of LBA4404 strain in infecting both cassava and cotton tissues are independent of the binary plasmid type, but it is rather associated with the nature of the LBA4404 genome. Agrobacterium strain EHA105, a derivative of A281, has previously been shown to be unsuitable for cassava transformation [15]. However, it was surprisingly found to be highly virulent with the cassava genotypes TME 13, TME 127, TME 8, TME 1, TMS I 91/02327 and 60444 [26]. These observations clearly demonstrate that in cassava, the virulence of Agrobacterium strains varies based on the Agrobacterium strain, cassava genotype, tissue type, and the T-DNA transfer ability of the binary or co-integrated resident plasmids present in the disarmed Agrobacterium vectors as previously demonstrated in Chrysanthemum moriflolium, Ramat [28]. Although the results of transformation frequencies and efficiencies of the transformed cassava in this study were slightly higher than other studies, it is clear that the transformation frequencies and efficiencies achieved in this study were still low (Table 1). The reason for the low efficiency might be attributed to the selection pressure of transformed tissues. The antibiotic typically used in selection of transformed tissues in this study is kanamycin, which, together with hygromycin, are considered very toxic to cassava tissues. This can possible be ameliorated by use of the kanamycin analogue, paromomycin, in selecting the transformed cassava tissues [17]. Polymerase chain reaction (PCR) and RT-PCR positive, transgenic cassava plants have been recovered in this study. This indicates that this transformation system has significant replicable potential for use in genetic transformation programmes at least for the apparently recalcitrant African cassava genotypes. Results of southern blot analyses show that there was minimal transgene insertion in the transgenic studies. This makes it possible for further segregation analysis on the transgenic varieties generated in this study. This also underscores the advantages of Agrobacterium-mediated plant transformation over particle bombardment. Particle bombardment leads to multiple gene insertion, thus making segregation analysis difficult [7].
In conclusion, this study has demonstrated successful and reproducible transformation of three Kenyan cassava landraces and the model cultivar TMS 60444 using immature leaf lobes as explants. This is considered a major impetus towards genetic improvement of cassava to introduce and/or improve the desired traits. Further, the findings of this study grossly dispute the general paradigm of thought that African cassava genotypes are seldom transformable.