Pancreatic Disorders & Therapy
Open Access
ISSN: 2165-7092
ISSN: 2165-7092
Research Article - (2013) Volume 3, Issue 1
Introduction: Pancreatic ductal adenocarcinoma (PDA) has the worst prognosis among cancers, mainly due to the high incidence of early metastases. RAN small GTPase (RAN) is a protein that plays physiological roles in the regulation of nuclear transport and microtubule spindle assembly. RAN was recently shown to mediate the invasive functions of the prometastatic protein osteopontin (OPN) in breast cancer cells. We and others have shown previously that high levels of OPN are present in PDA. In this study, we analyzed the expression and correlation of RAN with OPN in human pancreatic lesions, and explored their regulation in PDA cell lines. Methods: Real time PCR was used to analyze RAN and OPN mRNA levels in PDA, adjacent non-malignant, and benign pancreatic tissues. Expression levels were correlated with survival and different clinicopathological parameters using different statistical methods. Transient transfection studies using OPN and RAN plasmids, and knockdown experiments using siRNA were used to examine their mutual regulation. Results: OPN and RAN levels highly correlated with each other (p<0.0001). OPN or RAN levels did not correlate with venous lymphatic invasion, diabetes, obesity, T stage, BMI, or survival. However, we found a significant association between RAN levels and perineural invasion (HR=0.79, 95% CI 0.59, 1.07; p=0.0378.). OPN and RAN colocalized in PDA tissues and cell lines. Increasing RAN expression in PDA cells induced OPN transcription and RAN silencing reduced total OPN levels. OPN did not have any significant effect on RAN transcription. Conclusions: The high levels of RAN in PDA and its correlation with OPN and with perineural invasion suggest that RAN may contribute to PDA metastasis and progression through the induction of OPN. RAN’s role in the regulation of OPN in PDA is unique and could provide potential novel therapeutic strategies to combat PDA aggressiveness.
<Keywords: Pancreatic cancer, RAN, OPN
Pancreatic ductal adenocarcinoma (PDA) is the fourth leading cause of cancer death in the United States. Most patients with PDA have a poor prognosis that includes a median survival rate of less than 6 months and a 5-year survival rate of 3-5% [1,2]. At the time of diagnosis, the disease is often discovered to be in its late stages, as more than 85% of patients have tumors that have metastasized [2]. The aggressive nature of PDA can be attributed not only to its metastatic and invasive properties, but also to its resistance to chemotherapy and radiation. Currently, surgery remains the only option to decrease mortality in PDA patients. However, PDA still remains an elusive cancer that requires further studies to understand its molecular mechanisms and investigate possible therapeutic options.
Genetic instability is a known hallmark of cancer. One protein implicated in the mitotic instability of cancer cells is the RAN small GTPase (RAN). RAN is a universal nuclear-cytoplasmic transport protein that alters between two conformations–RAN-GTP and RAN-GDP. Specifically, RAN-GTP is localized to the nucleus of cells whereas RAN-GDP is found exclusively in the cytosol. This GTP/GDP switch allows RAN to migrate across nuclear pore complexes and transport protein and mRNA along with it [3-10]. Any disturbances in RAN expression could therefore lead to the abnormal transport of tumor oncogenes such as Akt or NF-κΒ [10]. RAN has also been shown to play an active role in microtubule polymerization, mitotic spindle formation, and cytoskeleton organization during mitosis. As a result, dysregulation of RAN has been shown to cause aneuploidy, microtubule dysfunction, and disturbances in cell migration [11,12].
High levels of RAN in tumors have been associated with poor prognosis in renal cell carcinoma, breast cancer, and epithelial ovarian cancer [13-15]. RAN expression has also correlated with tumor grade, incidence of metastasis, and overall survival rates in renal cell cancer. Experimental silencing of RAN has been shown to decrease cell proliferation and cell adhesion in vitro [15].
Osteopontin (OPN) is an integrin binding glycoprotein that derives its name from its original discovery in the bone matrix [16]. However, OPN is present in a wide variety of tissues of the body and has been implicated in various pathologic states such as neoplasia and inflammation [17]. Elevated levels of OPN were found in gastric, hepatocellular, and breast malignancies [18-21]. Most importantly, OPN has been shown to promote inflammation and metastasis by binding to multiple integrins and ligands in the CD44 receptor family. This ligand-receptor interaction results in the activation of various effector molecules including Ras, Src, MAPK, Akt, NF-κB, and matrix metalloproteinases (MMPs) that contribute to cancer development [22,23].
Recent studies have implicated RAN in the prometastatic invasive functions of OPN. Specifically, overexpression of OPN in breast cancer cell lines increases RAN expression, whereas RAN blockade induced an OPN-independent decrease in the cell metastatic properties [15]. We have recently established that OPN plays a critical role in mediating the proinflammatory and prometastatic effects of nicotine in PDA through inducing VEGF, MCP-1, and metalloproteinase MMP-9 [24-27]. However, the role of RAN in PDA and its relationship to OPN has not yet been elucidated. In this study, we analyze the expression of RAN in various pancreatic lesions, correlate its expression with OPN, and study their relationship in PDA cell lines.
Tissue acquisition
Histologically confirmed human invasive PDA samples (n=51) and benign lesions (n=7; IPMN, n=4; cystadenomas, n=3) were obtained from patients who underwent surgical resection at the Thomas Jefferson University Hospital between 2005 and 2011. Matching specimens from adjacent nonmalignant tissues were obtained and analyzed whenever available, allowing comparison to PDA (n=22). Tissue samples were either fixed in neutral formalin for histologic processing or incubated in RNA Later (Ambion) for RNA studies. All patients signed an appropriate informed consent for tissue acquisition and study. The study was approved by the Institutional Review Board at Thomas Jefferson University.
RNA extraction and real-time reverse transcription polymerase chain reaction (PCR)
Total RNA was isolated from PDA cells or pancreata using Trizol reagent (Life Technologies, Gaithersburg, MD). RNAs were quantified and input amounts were optimized for each amplicon. RAN, OPN and GAPDH (internal control) primers and Taqman probes were purchased by Applied Biosystems. Complementary DNA (cDNA) was prepared, diluted, and subjected to real-time PCR. Samples were measured in duplicates and the relative mRNA levels were presented as unit values of 2^[CT (GAPDH)-CT (Ran-OPN)], where CT represents the threshold cycle value defined as the fractional cycle number at which the target fluorescent signal passes a fixed threshold above the baseline.
Cell culture and treatment: Six PDA cell lines (AsPC-1, HPAF, MiaPaCa-2, Panc-1, PK-9, Hs-766T) were used to explore the basal levels of RAN and OPN. Eventually, two pancreatic cancer cell lines were used for the in vitro studies: MiaPaCa-2 and AsPC-1 (American Type Culture Collection, Manassas, VA). MiaPaca-2 cells, which express high levels of RAN and lower levels of OPN were used to explore the effect of knocking down RAN on OPN transcription, and the effect of overexpressing OPN on RAN transcription. In contrast, AsPC-1 cells, which express high levels of OPN and lower levels of RAN were used to explore the effect of knocking down OPN on RAN transcription, and the effect of overexpressing RAN on OPN transcription. Cells were cultured at 37°C and maintained in RPMI-1640 supplemented with 10% fetal bovine serum in a humid atmosphere of 5% CO2/95% air.
Transient transfection: To understand the interactions and mutual regulation of OPN and RAN in PDA cells, pGEM4–2rcDNA plasmid containing the full-length cDNA sequence that encodes mouse OPN was a generous gift from Dr. David Denhardt (Rutgers University, New Brunswick, NJ), and pCDNA.3-RAN plasmid was a generous gift from Dr. Mohamed El-Tanani (Centre for Cancer Research and Cell Biology School of Medicine, Dentistry & Biomedical Sciences, Queen’s University, Belfast, Northern Ireland) were used for overexpression studies. Cells were×plated10 at a concentration of 25/ml. At approximately 80% confluence, cells were depleted in a medium containing 0.1% BSA for 18 h. Cells were transfected using cationic liposome reagent TransFast (Promega, Madison, WI), with 10 μg of OPN or RAN cDNA or with the empty vectors. In addition, we performed parallel transfection experiments with a pGEM4- or pCDNA.3-enhanced green fluorescent protein (EGFP) plasmid and counted EGFP-expressing cells vs. total cell number to obtain an estimate of transfection efficiency (data not shown). We estimated our transfection efficiency at approximately 80%. After 3 h, the medium volume was increased to 2 ml. After 24 h, real time PCR was performed to compare gene levels in control and in OPN- and RAN-transfected cells.
siRNA sequences and constructs: Using GenBank™ sequence P62826 for human RAN and AK315461 for human OPN cDNAs and computer analysis software developed by Applied Biosystems/Ambion, candidate sequences in the RAN and OPN cDNA sequence for RNAi with no homology with other known human genes were selected and used during transient transfection experiments. Human mismatch or scrambled siRNA sequences (Applied Biosystems/Ambion; Austin, TX) possessing limited homology to human genes served as a negative control. Transfection was done with TransFast (Promega, Madison, WI) in AsPC-1 and Mia PaCa-2 cells as directed by the manufacturer. Cells were examined for RAN and OPN expression by real time PCR.
The relationships between OPN and RAN were examined using Spearman rank correlation. Tumor data only were examined for the relationships between RAN and OPN with other covariates, such as vascular lymphatic invasion, perineural invasion, diabetes, obesity, T stage, smoking status, and BMI. Summary statistics were calculated for each of the categories of the dichotomous covariates, using the Wilcoxon test for statistical tests.
The predictive ability of RAN and OPN was examined using ROC curves. Two logistic models were used, one with the dichotomous outcome of tumor vs. normal, and another with the dichotomous outcome of tumor vs. benign. OPN and RAN at their medians were analyzed using the Kaplan-Meier method with median survival and log-rank tests. Cox-proportional hazards regression was used with the continuous variables and hazard ratios. Robust variance analysis was conducted using Stata 11.2 (StataCorp LP, College Station, TX); the remainder of the analyses was carried out using SAS 9.3 (SAS Institute, Inc., Cary, NC). All in vitro experiments were performed 4 to 6 times. Data were analyzed for statistical significance by ANOVA with posthoc student t test analysis. Differences were considered significant at P ≤ 0.05.
Patient characteristics
An overview of the clinical patient data is summarized in table 1. The average age at the time of surgery was 67.5 years in the tumor group and 59.9 years in the benign group. The tumor group contained an equal number of males and females (51% and 49% respectively), whereas the benign group had a higher proportion of female patients (85.7%). BMI was also slightly lower in the tumor group when compared to the benign group (26.9 versus 30.9 respectively). The majority of patients with PDA presented at T3/T4 stage (81.4%) and showed evidence of both lymphatic invasion (83.9%) and perineural invasion (88.2%).
| All PDAs (n=51) | Matched normal tumor-adjacent tissue (n=22) | Benign (n=7) | |
|---|---|---|---|
| Gender [n (%)] | |||
| Male | 26 (51.0) | 9 (40.9) | 1 (16.7) |
| Female | 25 (49.0) | 13 (59.1) | 6 (83.3) |
| Age (years) [mean (SD)] | 67.5 (9.7) | 68.1 (9.7) | 59.9 (10.3) |
| BMI [mean (SD)] | 26.9 (5.5) | 27.3 (7.2) | 30.9 (8.9) |
| Staging [n (%)] | |||
| T1/2 | 8 (18.6) | 5(22.7) | |
| T3/4 | 43 (81.4) | 17 (77.3) | |
| Lymph node status [n (%)] | |||
| Positive | 41 (83.9) | 17 (76.7) | |
| Negative | 10 (16.1) | 5(23.3) | |
| Perineural invasion [n (%)] | |||
| Positive | 45 (88.2) | 18 (85.7) | |
| Negative | 6 (11.8) | 3(14.3) | |
| Differentiation [n (%)] | |||
| Well | 2 (3.9) | 1(4.5) | |
| Moderate | 36 (70.6) | 17 (77.3) | |
| Poor | 13 (25.5) | 4(18.2) | |
Table 1: Clinical characteristics of patients and tumors. BMI body mass index, PDA pancreatic ductal adenocarcinoma. PDA tissue (n=51) and if available, adjacent normal tissue (n=22) was anazlyed for RAN and OPN expression. Benign tissue comprises IPMN without invasion (n=4), and cystadenomas (n=3).
Expression of RAN and OPN in pancreatic tissues
RNA was extracted and cDNA was prepared from 51 PDA tissue specimens with matching adjacent tissue when available (n=22) and 7 benign lesions (IPMN, n=4; cystadenomas, n=3). Real time PCR was done using RAN and OPN primers with GAPDH as the internal control. The distributions of OPN and RAN were examined. OPN was found to be highly skewed and these distributions were improved by log-transformation; log-transformed OPN was used in all subsequent analyses where appropriate. RAN and OPN were significantly (P<0.0002) higher in the tumors when compared to normal adjacent tissue (Figure 1). Benign lesions, on the other hand, expressed significantly (p=0.0054) lower levels of RAN and OPN when compared to PDA tumor tissue. A Spearman correlation analysis of ln(OPN) vs. RAN for all tissue samples demonstrated a significant correlation between the two genes, suggesting that RAN and OPN may influence each other’s behavior (p<0.0001) (Figure 2).
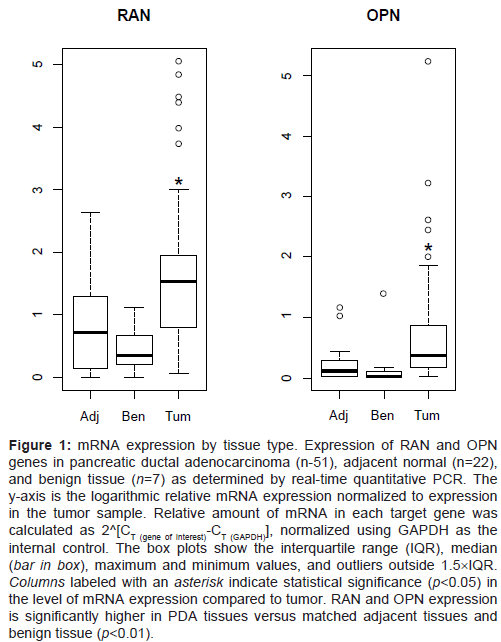
Figure 1: mRNA expression by tissue type. Expression of RAN and OPN genes in pancreatic ductal adenocarcinoma (n-51), adjacent normal (n=22), and benign tissue (n=7) as determined by real-time quantitative PCR. The y-axis is the logarithmic relative mRNA expression normalized to expression in the tumor sample. Relative amount of mRNA in each target gene was calculated as 2^[CT (gene of interest)-CT (GAPDH)], normalized using GAPDH as the internal control. The box plots show the interquartile range (IQR), median (bar in box), maximum and minimum values, and outliers outside 1.5×IQR. Columns labeled with an asterisk indicate statistical significance (p<0.05) in the level of mRNA expression compared to tumor. RAN and OPN expression is significantly higher in PDA tissues versus matched adjacent tissues and benign tissue (p<0.01).

Figure 2: Scatter plot illustrating RAN and OPN expression in all tissue types. The relationship between RAN and OPN was examined using Spearman rank correlation in PDA tumor tissue (n=52), normal adjacent tissue pairs (n=22) and benign IPMN lesions (n=7). A strong correlation was seen between RAN and OPN in all tissues with ρ=0.57 (p<0.0001).
Association with clinicopathological features
The expression of RAN and OPN were studied in relation to survival, T stage, resected lymph node status, perineural invasion, smoking, diabetes, and obesity. A significant association was seen between increased RAN expression and perineural invasion (HR=0.79, 95% CI 0.59, 1.07; p=0.0378.), while no other comparisons were significant. Although it did not reach statistical significance, a trend towards association between increased RAN and decreased survival was noted (p=0.107).
Predictive capability of RAN and OPN to distinguish tumor vs. non-tumor tissue
The predictive ability of RAN and OPN was examined using ROC curves. Two logistic models were used, one with the dichotomous outcome of tumor vs. normal and another with the dichotomous outcome of tumor vs. benign. Tables 2 and 3 and Figures 3A and 3B present the areas under the ROC curve for RAN, ln(OPN), and ln(OPN)+RAN. Ln(OPN)+RAN was the best predictor among the two-the combined predictive value for ln(OPN)+RAN was higher in discriminating between tumor vs. normal adjacent tissue (77%) (Table 2 and Figure 3A), and between tumor vs benign (89%) (Table 3 and Figure 3B).
| ROC | 95% ROC CI | OR | 95% OR CI | p-value | |
|---|---|---|---|---|---|
| RAN | 0.73 | (0.61, 0.85) | 2.68 | (1.60, 4.47) | 0.0002 |
| ln(OPN) | 0.74 | (0.63, 0.85) | 1.85 | (1.38, 2.48) | <0.0001 |
| ln(OPN)+RAN | 0.77 | (0.66, 0.88) |
Table 2: ROC values for tumor samples compared to matched adjacent normal (non-tumor) pancreatic tissue; 95% CI and OR are shown. RAN and OPN isoforms are overexpressed in tumor tissue relative to adjacent to normal expression.
| ROC | 95% ROC CI | OR | 95% OR CI | p-value | |
|---|---|---|---|---|---|
| RAN | 0.84 | (0.72, 0.97) | 2.68 | (1.34, 5.35) | 0.0054 |
| ln(OPN) | 0.83 | (0.61, 1.00) | 2.54 | (1.37, 4.74) | 0.0032 |
| ln(OPN)+RAN | 0.89 | (0.76, 1.00) |
Table 3: ROC values for tumor samples compared to benign pancreatic tissue; 95% CI and OR are shown. RAN and OPN isoforms demonstrated a strong association with the presence of malignancy (ROC=0.84, 0.83 respectively). Combining RAN and OPN slightly increased the predictive value for malignancy (ROC=0.89).
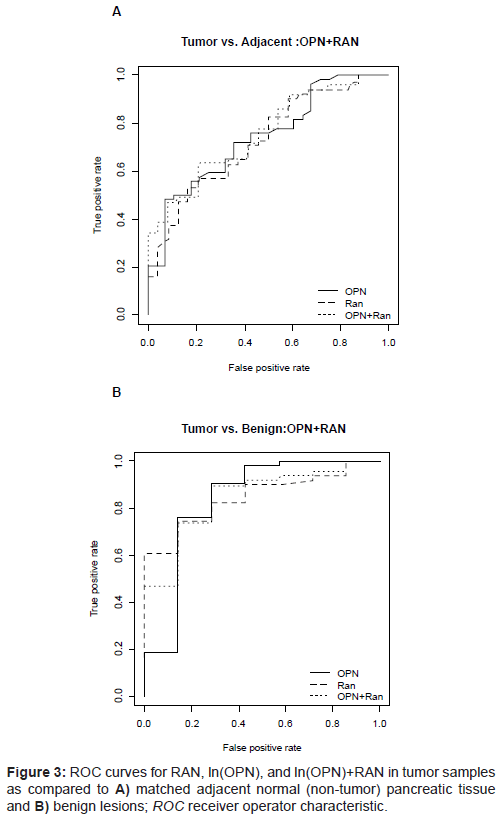
Figure 3: ROC curves for RAN, ln(OPN), and ln(OPN)+RAN in tumor samples as compared to A) matched adjacent normal (non-tumor) pancreatic tissue and B) benign lesions; ROC receiver operator characteristic.
Expression of RAN and OPN mRNA in different PDA cell lines
After extracting mRNA from the following cell lines (AsPC-1, PK- 9, Panc-1, HPAF, MiaPaCa-2, Hs-766T), baseline levels of RAN and OPN mRNA were evaluated for relative comparison. A wide range of RAN and OPN levels was noted (Figure 4). AsPC-1 cells express the least amount of RAN while MiaPaCa-2 cells contained the least amount of basal OPN mRNA. The Hs766T pancreatic cancer cell line contained the greatest amount of both RAN and OPN. We chose the two cell lines MiaPaCa-2 and AsPC-1 to conduct the following gainand loss-of function studies.
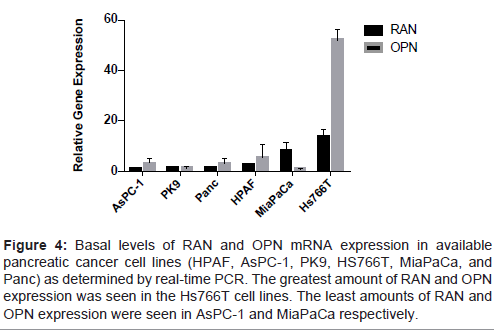
Figure 4: Basal levels of RAN and OPN mRNA expression in available pancreatic cancer cell lines (HPAF, AsPC-1, PK9, HS766T, MiaPaCa, and Panc) as determined by real-time PCR. The greatest amount of RAN and OPN expression was seen in the Hs766T cell lines. The least amounts of RAN and OPN expression were seen in AsPC-1 and MiaPaCa respectively.
OPN over- or under-expression does not affect RAN transcription Previous studies on breast cancer cells implicated RAN as a downstream effector of OPN [15]. Here, the relationship between these two genes in PDA cell lines was explored. Transient transfection of OPN plasmid significantly increased OPN transcription, but had no effect on RAN gene expression (Fig 5A). When we used OPN siRNA, OPN transcription was significantly reduced after 24 h, but had no effect on RAN transcription (Figure 5B). Interestingly, when we over expressed RAN in AsPC-1 cells, OPN transcription was significantly increased (four-fold) (Figure 6A). When RAN was knocked down using siRNA, OPN gene expression levels were significantly reduced (75% reduction) (Figure 6B). These data show that in PDA cells, RAN acts upstream of OPN to regulate its transcription.
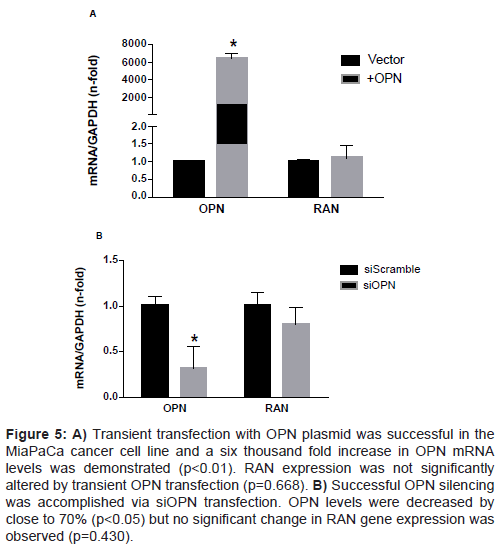
Figure 5: A) Transient transfection with OPN plasmid was successful in the MiaPaCa cancer cell line and a six thousand fold increase in OPN mRNA levels was demonstrated (p<0.01). RAN expression was not significantly altered by transient OPN transfection (p=0.668). B) Successful OPN silencing was accomplished via siOPN transfection. OPN levels were decreased by close to 70% (p<0.05) but no significant change in RAN gene expression was observed (p=0.430).
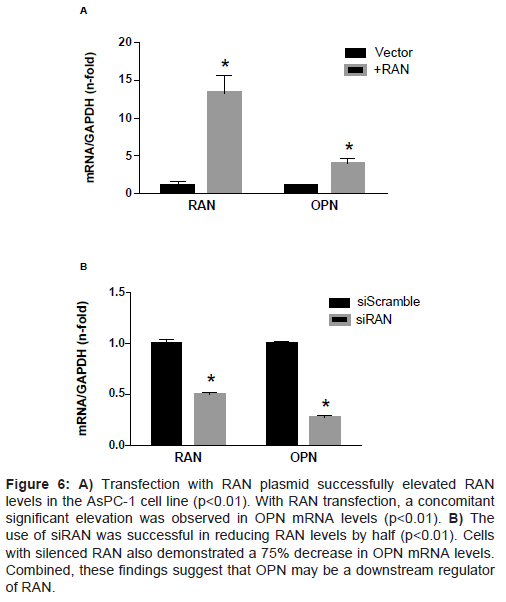
Figure 6: A) Transfection with RAN plasmid successfully elevated RAN levels in the AsPC-1 cell line (p<0.01). With RAN transfection, a concomitant significant elevation was observed in OPN mRNA levels (p<0.01). B) The use of siRAN was successful in reducing RAN levels by half (p<0.01). Cells with silenced RAN also demonstrated a 75% decrease in OPN mRNA levels. Combined, these findings suggest that OPN may be a downstream regulator of RAN.
The RAN small GTPase has classically been described as a Rasassociated protein involved in the nuclear-cytoplasmic transport of proteins and in the formation of the mitotic spindle [5-8]. Recently the focus on RAN has shifted to abnormalities arising from its dysregulation and subsequent genetic instability and cancer progression [3,4]. To our knowledge, this study is the first to investigate the expression of the RAN small GTPase in PDA. The significant elevation of RAN and OPN in PDA lesions when compared to normal adjacent tissue and benign tissue strongly implies that these proteins are involved in tumorigenesis and perhaps the progression from low to high grade dysplasia.
Here, we introduce RAN and OPN as novel tissue biomarkers for PDA. ROC analysis of RAN and OPN demonstrated a 77% accuracy when the predictive value of both markers is used together to distinguish between PDA and normal adjacent tissue, and an 89% accuracy in distinguishing between tumor and benign tissue. Hence, these two proteins have the potential to serve as useful markers in differentiating neoplastic pancreatic pathology. Although there are limited studies on the use of RAN as a biomarker, one group analyzed RAN expression in squamous cell carcinoma of the lung and found significantly increased levels of RAN in tumor tissue when compared to normal adjacent tissue. RAN also correlated with lung cancer stage, giving further support to its possible role as a tissue biomarker [28]. Many studies have considered OPN as a possible biomarker for many cancers. In gastric cancer, high levels of OPN have been associated with increased proliferation, increased stage, and metastasis [18]. Another study observed that OPN levels became elevated in the serum of patients one year before the diagnosis of hepatocellular carcinoma [29]. Similar results in pancreatic, prostate, and breast cancers, as well as in gliomas have further corroborated OPN as an important biomarker for identifying malignancy [30-36].
Metastasis is the most significant contributor of death due to cancer in a host. Although we found no significant correlation between RAN and OPN expression and venous/lymphatic invasion, there was a significant association between RAN expression and perineural invasion. Perineural invasion is a poorly understood process by which cancer cells metastasize to nerves and their surrounding neural sheaths [37]. Although metastatic spread via neural invasion is often overlooked, PDA has among the highest rates of perineural invasion when compared with other malignancies. Out of 51 PDA patients in our study, 45 (88%) showed evidence of perineural invasion. Perineural invasion has been associated with decreased survival and pain in PDA patients [38]. It has been hypothesized that the extremely high rates of perineural invasion in PDA may be attributed to prominent neurotropic effects on PDA cells and the proximity of the pancreas to prominent nerve plexuses (such as the plexuses at the celiac axis and the superior mesenteric artery). The molecular mechanisms mediating perineural invasion have implicated several important cytokines–NGF (nerve growth factor), BDNF (brain-derived neurotrophic factor), TGF-α (transforming growth factor), and EGFR (epidermal growth factor receptor) [37,39]. Many of these molecules promote the activity of MMP-2 and MMP-9, two important metalloproteinases that not only promote perineural invasion, but also have been shown to be downstream effectors of OPN [40,41]. High levels of RAN showed strong association with reduced survival although it did not reach significance (p=0.107). Nonetheless, the interaction between RAN and the various cytokines responsible for perineural invasion may be an interesting avenue for exploration in the future.
Although the mechanism by which invasion is regulated by RAN has not been resolved, a previous study in breast cancer cell lines has implicated OPN as coordinating with RAN to enhance metastatic and invasive potential (15). We were intrigued to notice that contrary to prior studies in the rat mammary cell lines, altering OPN levels in PDA cells did not significantly alter levels of RAN. Instead, the opposite was found–altering RAN produced corresponding changes in OPN expression. Furthermore, data from our lab have shown that overexpressing RAN also induced a significant (p<0.05) stimulation of the expression of CD44, a cell surface receptor for OPN (unpublished observation). This association suggests that in addition to increasing OPN, RAN may also support transcriptional activity of this OPN receptor that allows Ras, Akt, and other oncogenes to become more active [22]. An extremely strong correlation between OPN and RAN expression in human pancreatic tissue specimens further suggests that these two proteins are intimately related. Nonetheless, the signaling pathways involved in the RAN-mediated regulation of OPN are yet to be studied. Furthermore, whether RAN directly impacts OPN transcription by activating its promoter or via stimulation of other signaling pathways known to activate or deactivate OPN is currently unknown. Studies to answer these questions are currently ongoing in our laboratory.
In conclusion, the high expression of RAN and OPN mRNA in human PDA tissue suggests that these two genes may serve as important discriminating biomarkers between benign and malignant pancreatic pathology. The strong association between increased RAN and perineural invasion further suggests a novel role for this small GTPase in the aggressive nature of PDA. Finally, the strong correlation between RAN and OPN in pancreatic tissue and the hypothesis of OPN as a downstream effector of RAN is unique and can provide a novel therapeutic target in PDA patients.
This work was supported by NIH grant 1R21 CA133753-02 as well as funds conated in support of the Jefferson Pancreas, Biliary and Related Cancer Center.