Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2016) Volume 7, Issue 1
Mussel (Mytilus edulis) farms in Newfoundland, Canada were investigated as potential sites to hold adult lobsters Homarus americanus in inshore benthic cages. The goals of this project were to determine if lobsters can be maintained for prolonged periods in cages and survive and grow by feeding on mussels dropping-off culture lines. The effects of biotic and abiotic factors on the moulting, growth rates and serum protein concentrations were monitored at regular intervals in both the field and the lab over 6 months. Although survival rates were high under mussel lines, the moulting rate was low and analysis of serum protein concentration showed they were in a poorer condition than fed lobsters in lab experiments. In the laboratory diet type, temperature, feeding frequency and compartment size were manipulated to determine possible factors influencing survival and growth of the lobsters in the field. In the lab, moulting was highest at 15°C and survival lowest at 5ºC; lobsters fed a mixed versus a mussel only diet were healthier. In a separate lab experiment, lobsters that were fed twice weekly attained a larger size at post-moult than those fed once per month. However, feeding frequency did not affect survival or the number of animals moulting. The lab experiments suggested that the combination of low temperature and infrequent food input was the cause of the low moulting rate and overall quality of the lobsters in the field. This project showed although lobsters can be stored in benthic cages in the field for up to 6 months, relying on mussel drop-off alone is limited, and lobsters may need supplemental feeding in order to produce a larger, higher quality product for market. Initial results also suggest the promise of incorporating lobsters into a multitrophic aquaculture system as a means to remove moribund mussels underneath culture lines.
Keywords: Aquaculture; Blue mussel; Growth; Lobster; Moulting; Serum protein; Benthic cages; Homarus americanus
The American lobster, Homarus americanus (Milne-Edwards) supports is a multi-billion dollar fishery in Atlantic Canada and New England. Generally it takes between 4 and 12 years for H. americanus to reach marketable size [1]. Because of this long growth time the fishery is highly regulated. In Atlantic Canada, the lobster fishing season varies in duration among the lobster fishing areas, ranging from 2 to 6 months [2]. On the island of Newfoundland, the lobster fishing season is comparatively short compared with the rest of Canada and New England, starting between April and May and closing between June and July. Because the lobster fishing seasons are regulated, the commercial trade of adult American lobsters has largely focused on development of holding methods so that live lobsters are available year round. For example, most of the lobster marketing companies in Nova Scotia and New Brunswick hold lobsters in indoor tanks or outdoor impoundments. In the indoor facilities the lobsters are held at temperatures of 1-3°C. This low temperature reduces the lobster’s metabolic rate and maintains them in the intermoult (hard shell) stage, allowing the animals to be held for several months with minimal loss of product. The lobsters are not fed during this time and in order to combat the effects of starvation only lobsters with high serum protein concentrations can be stored in this way (Stewart Lamont, Tangier Lobster Company Limited, pers. comm.). However, the maintenance and logistics required for these holding methods are expensive, and cannot be employed on a small scale or in remote locations.
Although there have been efforts to raise and release juvenile clawed lobsters to repopulate areas [3,4] and there has been some work on on-growth of juvenile clawed lobsters in the field [5-8] they have had limited success. Therefore, the harvest of both H. americanus and its European counterpart, Homarus gammarus (Linnaeus) primarily remain a wild capture fishery. In contrast there has been much more research directed towards the potential for aquaculture and on-growth in spiny lobsters (Genus Jasus and Panulirus) [9]. These can either be cultured all the way from the larval pueruli stage [10-12], or sub-adults can be “fattened” for market [13,14]. The animals are held in cages, and although feeding with commercial pellet meals has met with some success, the highest growth and survival rates have been obtained when the lobsters are fed fresh mussels [10,15,16].
The high survival and growth of spiny lobsters on mussel flesh is echoed by the fact that populations of H. americanus have been enriched below or near commercial blue mussel (Mytilus edulis, Linnaeus) operations. The anchor line buoys provide shelter for the animals and the mussels dropping off the culture lines may be a potential food source for these lobsters [17-20]. Although the presence of bivalve farms has been reported to enrich lobster populations, the rapid expansion of this sector in recent years has led to concerns about its sustainability and in particular the problems with the input of excess nutrients into the environment and the impacts on local fauna [21-23].
In a pilot study, lobsters maintained for 6 months in cages in 6-8 m of water and fed twice weekly by hand had an 85% survival rate. 40% of the animals moulted and increased their body mass by approximately 35% [24]. However, the logistics and costs associated with hand feeding precluded the development of this method on a large scale. The goal of the present study was to investigate an alternative method for storage to determine if lobsters can survive and grow when held for extended periods. Lobsters were held in the field under blue mussel farms with the idea that mussels dropping off the culture lines could supply a food resource for lobster growth. In turn the lobsters could help remove moribund mussels that would otherwise rot and stagnate on the bottom. Because of the logistics associated with constant monitoring of animals and environmental conditions in the field, experiments were also conducted in the laboratory. Using the current literature, feedback from the mussel growers, and diver observations, the potential variables that lobsters may experience in the field such as the temperature change, the frequency of mussel drop-off and type of food items reaching the cages (mussels only or mussels with supplemental items) were manipulated in the laboratory. These experiments allowed us to more accurately determine how these variables potentially affected the moulting, survival and health of the cage-held lobsters in the field.
Housing and management
Three series of experiments were carried out, two were performed in the laboratory (Department of Ocean Sciences, Memorial University) where factors could be manipulated and a field experiment was conducted at Sunrise Fish Farms (Triton, NL). The animals used in the laboratory experiments were purchased from Clearwater Seafood, Nova Scotia, and both male and female intermoult animals were used. Due to permitting regulations, the lobsters used in the field experiments were purchased from local harvesters at Triton and only male intermoult lobsters were used. All the experimental animals (both in the lab and field) were held in individual compartments in plastic coated 2.5 cm wire mesh cages during the 6 month experimental period. The cages measured 1.20 × 0.90 × 0.30 m in depth either with 24 separate compartments, individually measuring 0.30 × 0.15 × 0.30 m in depth, or with 12 larger compartments, individually measuring 0.45 × 0.20 × 0.30 m in depth, with 2 cages held side by side (total n=24). Each individual lobster, isolated from other lobsters in separate compartments of the cage acted as a replicate. This cage design and experimental set-up was chosen as it represented the exact protocol for lobster on-growth in Newfoundland that would be used by Jerseyman Island Fisheries Ltd [24].
Sampling methods
The lobsters in the two laboratory experiments were checked every other day and any mortalities were recorded and removed, at the same time any lobsters undergoing moulting were noted. The following parameters were measured once every 2 months in the laboratory and once every 3 months in the field.
Lobster growth was measured by recording body mass and carapace length. For body mass the lobsters were removed from the cages and the water was allowed to drain from the bronchial chambers for 3 to 5 mins. The animals were then wiped dry and measured to the nearest 0.1 g. The carapace length was measured along the dorsal line between the eye socket and the posterior margin of the carapace.
The serum protein concentration is a good indicator of quality (meat content and health) and physiological condition in lobsters. It was measured by withdrawing a 500 μl sample of haemolymph from arthrodial membrane at the base of the fourth walking legs. This sample was then injected onto the sample well of a pre-calibrated Brix/RI-Chek Digital Pocket Refractometer (Reichert Analytical Instruments, Depew, NY, USA). The time between withdrawal of the haemolymph and processing of the sample did not exceed 90 seconds. The total serum protein concentration was then calculated from the RI as outlined in Wang and McGaw [25].
Experimental protocols
Field experiment: Cage location and compartment size: The objective of the field experiment was to assess the input of blue mussels as a food source as well as the effect of compartment size on lobster survival, growth, and health status. In line with the Department of Fisheries and Oceans permitting requirements, all the lobsters used in the field experiments were males and had to be purchased from local harvesters; they had a body mass (mean ± SD) of 601.0 ± 92.8 g. The lobsters (n=192) were held in benthic cages (10 to 13 m depth) at Sunrise Fish Farms near Triton, NL (N49º 29’ 03.03’, W55° 45’ 03.58’). Half of the lobsters were maintained in cages with regular sized compartments, while the other half were kept in cages with the larger compartments. Each series of cages was strapped together to ensure they would remain in the same location. Temperature loggers (iBCod, type G, Ste-Juline, QC, Canada) were attached to the cages and recorded the temperature every 4 h during the 6 months (June to December 2013) experimental period. Half of the experimental animals (n=48 in regular compartments, 48 in large compartments) were placed directly under the mussel culture lines of Sunrise Fish Farms. The idea being that they would be able to feed on mussels dropping off the culture lines. The remaining cages (n=96 lobsters) were set approximately 15-25 m away from the mussel farm where there was no evidence of mussel drop-off from the culture lines.
Lab experiment 1: Temperature and diet type: The first lab experiment was designed to test the effects of temperature and diet type on survival and growth of cage-held lobsters. The lobsters were held in 3000 L flow-through seawater tanks maintained at either 5, 10 or 15ºC, representing typical temperature ranges experienced by lobsters in the wild [26,27]. The temperature in each tank was checked daily; the temperature in the 10°C and 15°C tanks varied ± 1°C, while the 5°C tanks fluctuated ± 2°C, during the 6 month experimental period. Each tank was equipped with air diffuser stones which maintained the water oxygen content between 92% and 98% saturation. The photoperiod was maintained on a 12L:12D cycle. Approximately equal number of lobsters (intermoult stage) of both sexes, with a body mass (mean ± SD) of 548.8 ± 53.3 g were purchased from Clearwater Seafood, NS. The lobsters were acclimated to laboratory conditions (and fed) for one month before the experiment commenced. The experiment was carried out between January and June 2013, 48 lobsters were held at each temperature (5°C,10°C,15°C) with 24 lobsters per cage, isolated in separate (small size) compartments; each cage (total of 6 cages) was housed in a separate tank. Each individual animal was fed to satiation twice weekly; one group of lobsters (n=24) in each temperature regime was fed a mixed diet (diet changed weekly-shrimp, squid, fish, blue mussel, scallop mantle or crab); while the other group (n=24) was only fed blue mussels. The tanks were cleaned at the end of each week, when any uneaten food and mortalities were removed.
The identification of the different moult stages of adult lobster was carried out by staging the developmental morphology of the pleopods following the methods outlined in reference [28]. In the present study, lobster pleopods were sampled at 1 month intervals and each sample was immediately photographed using Infinity Capture Imaging Software at 40X magnification. Haemolymph samples were collected weekly for lobsters in premoult and those that underwent moulting and the serum protein concentration was measured immediately; haemolymph samples continued to be collected from the moulted lobsters until the experiment was terminated.
Lab experiment 2: Feeding frequency and compartment size: In the second laboratory experiment the effect of feeding frequency and cage compartment size was investigated. The animals were maintained in cages (6 cages total) in the laboratory from June to December 2013 in a 45,000 l flow through tank, using ambient aerated sea water pumped from 5 m depth in Logy Bay, Newfoundland, Canada. Temperature data loggers (iBCod, type G, Ste-Juline, QC, Canada) were attached to cages and recorded water temperature every 4 h. Ninety six intermoult male and female lobsters with a body mass (mean ± SD) of 363.0 ± 59.3 g were purchased from Clearwater Seafoods. Half of the lobsters (n=48) were held in cages with 24 individual compartments of 0.20 × 0.30 × 0.30 m in depth, while the other half were held in cages with 12 larger compartments of 0.45 × 0.20 × 0.30 m in depth. Since a mussel only diet proved to be restrictive (lab experiment 1) and divers also noted other prey items in the vicinity of the cages each individual lobster was fed a mixed diet comprising of approximately 50% blue mussels supplemented with fish, squid, crab and scallop mantle. Following discussion with the mussel growers on potential timing and rates of drop-off, one group (n=24 lobsters per both compartment sizes) was fed to satiation twice weekly, while the remaining lobsters were fed once per month. To avoid fouling of the water, any remaining feed was removed at the end of each week.
Statistical analyses of lobsters mortality and moulting rates in various conditions, were analyzed using Kaplan-Meier survival curves (Prism v5.0, GraphPad Software Inc., La Jolla. CA). The Kaplan-Meier survival curves plot fractional survival/moulting (Y) as a function of time (X). It can be used to analyse the time to any event (usually death/ moult) that can only happen once. The data from the survival curves was then compared using a Mantel-Cox log rank test which examines the actual amount of events in relation to expected number of events and gives a Chi-squared statistic.
Changes in serum protein concentration as a function of diet type (Figure 1) were analysed with linear regression analyses using Prism (GraphPad Software Inc., La Jolla. CA). Growth (body mass and carapace length increments) and serum protein concentration were analysed using either ANOVA or repeated measures ANOVA (Sigma Stat). If significant differences were obtained they were further analysed with Tukey post-hoc tests. Statistical significance in all tests was accepted at the P<0.05 level. All the data is presented as the mean value ± the standard deviation.
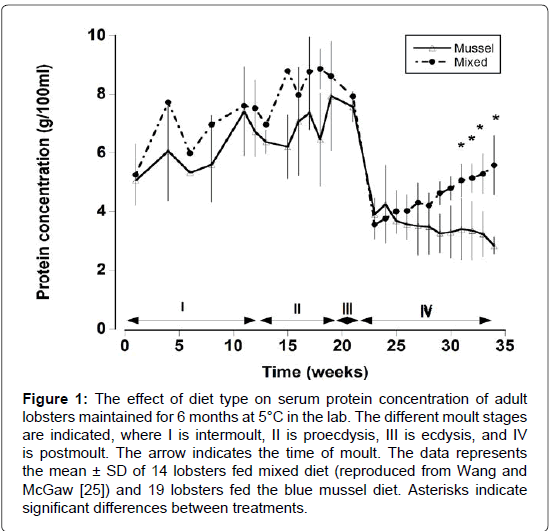
Figure 1: The effect of diet type on serum protein concentration of adult lobsters maintained for 6 months at 5°C in the lab. The different moult stages are indicated, where I is intermoult, II is proecdysis, III is ecdysis, and IV is postmoult. The arrow indicates the time of moult. The data represents the mean ± SD of 14 lobsters fed mixed diet (reproduced from Wang and McGaw [25]) and 19 lobsters fed the blue mussel diet. Asterisks indicate significant differences between treatments.
Field experiment: Cage location and compartment size
The benthic water temperature (10-13 m) at Triton increased from 3.5°C in late June reaching between 5°C and 7°C, during the period between mid July to early November. Thereafter the water temperature dropped to around 2°C by mid-December (Figure 2). The temperatures recorded on cages under the mussel lines and those situated away from the mussel lines were almost identical to one another.
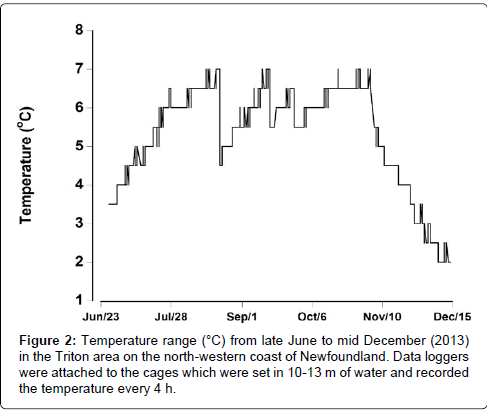
Figure 2: Temperature range (°C) from late June to mid December (2013) in the Triton area on the north-western coast of Newfoundland. Data loggers were attached to the cages which were set in 10-13 m of water and recorded the temperature every 4 h.
The lobsters held in cages in the field exhibited a low mortality rate, ranging from 2.1% to 4.2% (Table 1) and the majority of these mortalities occurred in the last 3 months (mid-September to mid-December). The moulting rate was comparatively low and ranged between 10.4% to 18.8% in all treatment groups (Table 1). Statistical analysis could not be performed on this data, because the limited inspection at the field sites (once per 3 months) did not allow accurate assessment of the exact time of each individual mortality/moulting event.
| Cage Location/ Compartment Size | Parameter | Number | Cumulative Mortality and Moulting Over 6 Months (Number and Percent) |
||
|---|---|---|---|---|---|
| June | September | December | |||
| Under Mussel Line (Large) | Mortality Moulting |
48 | 0 (0) 0 (0) |
1 (2.1%) 1 (2.1%) |
1 (2.1%) 6 (12.5%) |
| Under Mussel Line (Small) | Mortality Moulting |
48 | 0 (0) 0 (0) |
1 (2.1%) 3 (6.3%) |
2 (4.2%) 5 (10.4%) |
| Away Mussel Line (Large) | Mortality Moulting |
48 | 0 (0) 0 (0) |
0 (0) 9 (18.8%) |
2 (4.2%) 9 (18.8%) |
| Away Mussel Line (Small) | Mortality Moulting |
48 | 0 (0) 0 (0) |
0 (0) 4 (8.3%) |
2 (4.2%) 5 (10.4%) |
Table 1: Mortality and moulting rates of adult lobsters held in the field at Triton, NL. The animals were either held under mussel lines, or on the open sea bed and in two different compartment sizes and measurements taken at the start of the experiment (June) and at 3 month periods thereafter (September, December). Data is shown as cumulative numbers and these are expressed as a percentage of the total number in parentheses.
The moulted lobsters in cages on the open sea bottom exhibited an increase in body mass and carapace length of 22.32% ± 7.74% and 6.45% ± 1.53% respectively, while for those situated under mussel lines body mass and carapace length increased by mean levels of 20.33% ± 4.70% and 7.04% ± 2.26%, respectively. There was no significant effect of cage location or compartment size on these values (Figure 3) (Two way ANOVA, location, F=0.58, P=0.459; compartment size, F=0.12, P=0.738). Non-moulted lobsters were able to increase overall body mass during the 6 month period, nevertheless this increase was low, ranging between 0.39% and 1.92%, for both cage locations. The non-moulted lobsters in cages on the open seabed had a significantly higher increase in body mass (1.85% ± 1.15%), than those maintained under mussel lines (0.67% ± 1.29%) (Two way ANOVA, F=32.23, P<0.001). The compartment size did not have a significant impact on change in body mass of non-moulted lobsters (Two way ANOVA, F=0.92, P=0.34).
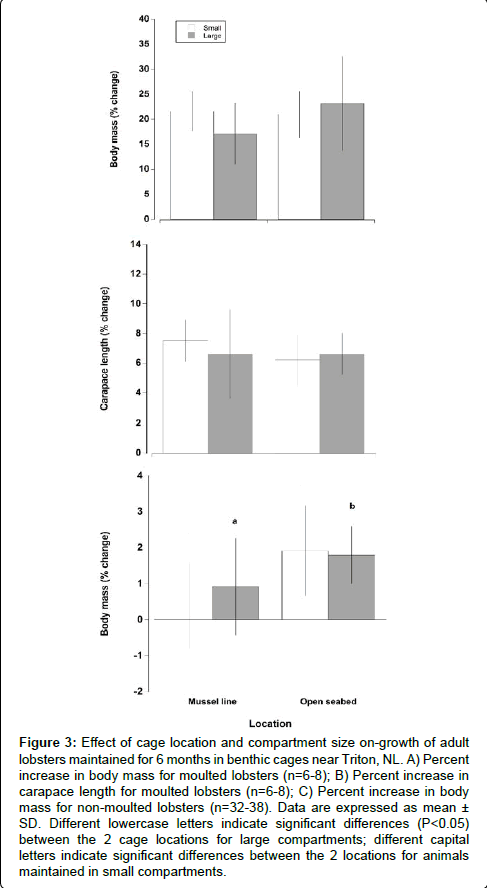
Figure 3: Effect of cage location and compartment size on-growth of adult lobsters maintained for 6 months in benthic cages near Triton, NL. A) Percent increase in body mass for moulted lobsters (n=6-8); B) Percent increase in carapace length for moulted lobsters (n=6-8); C) Percent increase in body mass for non-moulted lobsters (n=32-38). Data are expressed as mean ± SD. Different lowercase letters indicate significant differences (P< 0.05) between the 2 cage locations for large compartments; different capital letters indicate significant differences between the 2 locations for animals maintained in small compartments.
The serum protein levels decreased significantly over the 6 month study for both lobsters under mussel lines and on the open seabed (Table 2) (Two way RM ANOVA, F=279.3, P<0.001). The decrease in final serum protein levels was more pronounced in lobsters maintained on the open seabed (6.00 ± 1.39 to 2.35 ± 1.02 g/100 ml) compared with those held under mussel lines (5.83 ± 1.02 to 3.25 ± 0.911 g/100 ml) (Tukey post-hoc test, P<0.001). The compartment size also had a significant effect on the final serum protein concentration, but only for the lobsters ranched underneath mussel lines (Tukey post-hoc test, P<0.01; Table 2). The location of the compartment in the cage also had an effect on serum protein levels (Figure 4). For example, in the 24 compartment cages the 4 corner compartments had a large surface area that could potentially come into direct contact with organic material, while the lobsters in the middle 6 compartments were surrounded by animals in other compartments therefore had a much lower surface area (1 upper and 1 lower) directly in contact with the environment. On the open seabed, the lobsters held in corner compartments had significantly higher serum protein levels (2.70 ± 0.74 g/100 ml) than those held in the middle compartments (1.94 ± 0.56 g/100 ml) (Two way ANOVA, F=6.1, P<0.05). Although a similar trend was observed for lobsters held under the mussel lines (corner=3.45 ± 0.73 g/100 ml, middle=2.93 ± 0.9 g/100 ml), this proved to be statistically insignificant (Tukey post-hoc test, P=0.231).
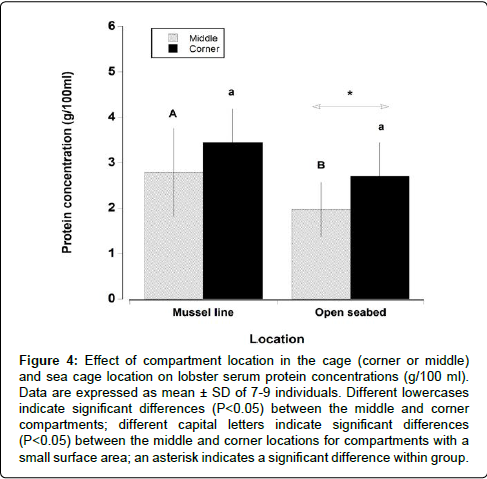
Figure 4: Effect of compartment location in the cage (corner or middle) and sea cage location on lobster serum protein concentrations (g/100 ml). Data are expressed as mean ± SD of 7-9 individuals. Different lowercases indicate significant differences (P < 0.05) between the middle and corner compartments; different capital letters indicate significant differences (P< 0.05) between the middle and corner locations for compartments with a small surface area; an asterisk indicates a significant difference within group.
Only a few lobsters moulted and because the cages were only checked every 3 months, the data for moulted lobsters was limited (shown in parantheses in Table 2). The general trend was that serum protein levels dropped after moulting, and serum protein levels (at both locations) continued to decrease thereafter. This decrease appeared to be more pronounced for lobsters settled on the open seabed (8.84 ± 1.08 to 1.46 ± 0.17 g/100 ml), than for lobsters settled under mussel lines, (7.12 ± 1.68 to 1.76 ± 0.57), however, this difference proved to be statistically insignificant (Student t test, T=1.62, P=0.131).
| Sea Cage Location | Compartment Size | Serum Protein (g/100 ml) Over 6 Months Mean ± SD |
||
|---|---|---|---|---|
| June | September | December | ||
| Under Mussel Line | Large | 5.86 ± 1.11 | 3.86 ± 0.92 | 3.50 ± 0.92aA |
| Under Mussel Line | Small | 5.73 ± 0.90 (7.12 ± 1.68, n=3) |
3.60 ± 0.94 (2.20 ± 0.511, n=3) |
2.98 ± 0.86bA (1.76 ± 0.57, n=3)A |
| Away Mussel Line | Large | 6.05 ± 1.39 | 2.65 ± 1.01 | 2.22 ± 0.88aB |
| Away Mussel Line | Small | 5.96 ± 1.41 (8.33 ± 1.08, n=11) |
2.95 ± 1.18 (1.99 ± 0.52, n=11) |
2.47 ± 1.13aB (1.46 ± 0.17, n=11)A |
Table 2: Serum protein concentration of adult lobsters at Triton area, at 0, 3 and 6 months (June-December). The animals were held under mussel lines or on the open seabed in 2 different compartment sizes. The data represent the mean ± SD of 46-48 individual lobsters at each time point. Different lowercase letters indicate significant differences (P<0.05) between the 2 compartment sizes in the same location; different capital letters indicate significant differences (P<0.05) between the 2 sea cage locations for the same compartment size. Data in parentheses are from molted lobsters.
Lab experiment 1: Temperature and diet type
Temperature had a significant effect on lobster survival rate (Table 3) (Mantel-Cox Test, Chi square=53.49, P<0.001). The lowest mortality occurred at 10°C (4.2%-12.5%), and at this temperature the diet did not have a significant effect on survival rate (Mantel-Cox Test, Chi square=1.09, P=0.297); these mortalities only occurred during the final 2 months of the experiment. The highest mortality rate (79.2%) was recorded at 5°C for lobsters fed the mussel only diet; this mortality rate was significantly higher than the 50% rate measured for the group fed a mixed diet at 5°C (Mantel-Cox Test, Chi square=4.11, P<0.05). The mortality rate was also relatively high (50%) for lobsters in 15°C fed on the mussel diet and this was significantly higher than its counterparts (12.5%) fed a mixed diet (Mantel-Cox Test, Chi square=6.58, P<0.05). The mortalities in the 15°C mussel diet nearly all occurred during the final month of the study and all these had recently moulted. In contrast, very few mortalities were observed in post-moult lobsters fed a mixed diet at 15°C.
The experimental temperature also influenced the incidence of moulting (Table 3) (Mantel-Cox Test, Chi square=71.70, P<0.001). In 5°C, only 2 lobsters moulted during the 6 month experimental period, these occurred at beginning of the study and both of these animals were maintained on a mixed diet. There was a significant effect of diet on moulting at 10°C (Mantel-Cox Test, Chi square=7.65, P<0.01). Moulting rates were similar (and low) between the 2 groups during the first 5 months; there was a substantial increase in the moulting rate for the mixed diet lobsters in the final month, but none of the animals fed the mussel only diet moulted at this time. The highest moulting rates occurred in lobsters maintained at 15°C with most of the animals undergoing this process during the final three months of the study (66.7% for mixed diet and 83.3% for mussel diet).
| Temperature and Diet | Parameter | Number | Monthly Cumulative Mortality and Moulting Over 6 Months | |||||
|---|---|---|---|---|---|---|---|---|
| February | March | April | May | June | July | |||
| 5°C Mussel | Mortality Moulting |
24 | 1 (4.2%) 0 (0) |
2 (8.3%) 0 (0) |
6 (25.0%) 0 (0) |
9 (37.5%) 0 (0) |
13 (54.2%) 0 (0) |
19 (79.2%)aA 0 (0)aB |
| 5°C Mixed | Mortality Moulting |
24 | 0 (0) 1 (4.2%) |
1 (4.2%) 2 (8.3%) |
2 (8.3%) 2 (8.3%) |
3 (12.5%) 2 (8.3%) |
8 (33.3%) 2 (8.3%) |
12 (50.0%)bA 2 (8.3%)aC |
| 10°C Mussel | Mortality Moulting |
24 | 0 (0) 0 (0) |
0 (0) 1 (4.2%) |
0 (0) 1 (4.2%) |
0 (0) 1 (4.2%) |
1 (4.2%) 1 (4.2%) |
1 (4.2%)aC 1 (4.2%)bB |
| 10°C Mixed | Mortality Moulting |
24 | 0 (0) 0 (0) |
0 (0) 1 (4.2%) |
0 (0) 1 (4.2%) |
0 (0) 2 (8.3%) |
2 (8.3%) 2 (8.3%) |
3 (12.5%)aB 9 (37.5%)aB |
| 15°C Mussel | Mortality Moulting |
24 | 0 (0) 0 (0) |
1 (4.2%) 0 (0) |
1 (4.2%) 3 (12.5%) |
1 (4.2%) 11 (45.8%) |
2 (8.3%) 15 (62.5%) |
12 (50.0%)aB 20 (83.3%)aA |
| 15°C Mixed | Mortality Moulting |
24 | 0 (0) 0 (0) |
0 (0) 0 (0) |
3 (12.5%) 2 (8.3%) |
3 (12.5%) 7 (29.2%) |
3 (12.5%) 11 (45.8%) |
3 (12.5%)bB 16 (66.7%)aA |
Table 3: Mortality and moulting rates of adult lobsters maintained in the lab at 3 temperatures (5, 10, 15°C) and fed 2 diet types (mussel and mixed) over a 6-month period in the lab. The monthly recorded data are shown as cumulative numbers and these are expressed as a percentage of the total number in parentheses. Different lowercase letters indicate significant differences (P<0.05) between the 2 diets in the same temperature; different capital letters indicate significant differences (P<0.05) among the 3 temperatures for the same diet.
The limited amount of data for the 5°C treatment and 10°C mussel diet precluded statistical analysis on all combinations. Analysis of the remaining data showed no significant effect of temperature or diet on growth (Two way ANOVA, F=0.21, P=0.818). Following moulting lobsters increased in body mass by 29.32% ± 7.21% while an increase of carapace length of 8.89% ± 1.36% was observed (Figures 5A and 5B). There were only slight increases (1.3%-2.5%) in body mass for non-moulted lobsters (Figure 5C) and there was no consistent trend in change in body mass as a function of temperature or diet in these animals.
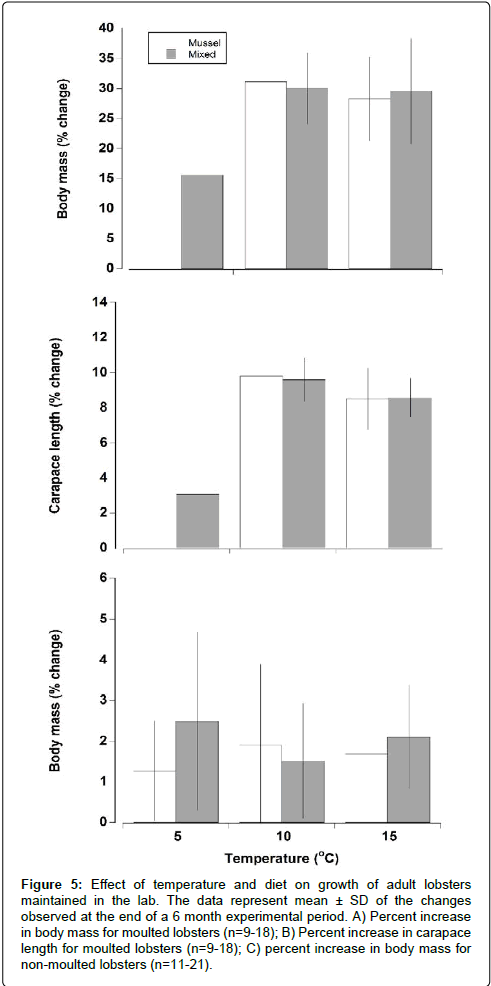
Figure 5: Effect of temperature and diet on growth of adult lobsters maintained in the lab. The data represent mean ± SD of the changes observed at the end of a 6 month experimental period. A) Percent increase in body mass for moulted lobsters (n=9-18); B) Percent increase in carapace length for moulted lobsters (n=9-18); C) percent increase in body mass for non-moulted lobsters (n=11-21).
Serum protein concentration was used as an indicator of the physiological and nutritional status of the animal. Temperature and diet had a significant effect on the final serum protein concentration (Table 4). Serum protein concentration increased with increasing temperature in non-moulted lobsters fed both diets after 4 months (2 way RM ANOVA, F=105.1, P<0.001). Lobsters in 5°C and 15°C fed a mixed diet had higher serum protein levels than those fed the mussel diet, but there was no effect of diet at 10°C (Two way ANOVA; diet, F=5.96, P<0.05; interaction, diet and temperature, F=4.25, P<0.05) in the final measurement of serum protein concentration of non-moulted animals (5°C and 10°C in the 6th month; 15°C in the 4th month).
| Temperature | Diet | Serum Protein (g/100ml) over 6 Months Mean ± SD | |||
|---|---|---|---|---|---|
| January | March | May | July | ||
| 5°C | Mussel | 4.62 ±0.75 | 4.99 ± 0.88 | 4.85 ± 0.80aC | 4.98 ± 1.44bB |
| 5°C | Mixed | 4.62 ± 1.76 | 4.96 ± 1.24 | 5.67 ±1.10aC | 6.32 ± 0.85aB |
| 10°C | Mussel | 3.97 ± 1.18 | 5.55 ± 1.03 | 6.77 ± 1.00aA | 7.92 ± 1.05aA |
| 10°C | Mixed | 5.12 ± 0.99 | 5.89 ± 1.06 | 6.69 ± 0.75aB | 7.65 ± 0.78aA |
| 15°C | Mussel | 5.20 ± 0.90 | 6.37 ± 1.25 | 7.73 ± 1.85bAB | *3.47 ± 0.48aC |
| 15°C | Mixed | 5.22 ± 1.26 | 6.91 ± 0.79 | 8.35 ± 0.85aA | *4.16 ± 1.83aC |
Table 4: Serum protein concentrations (g/100 ml) in 3 different temperature and 2 diet treatments. Samples were taken at 2 month intervals between January and July, data represents the mean + SD of 10-24 individuals at each time point. For the 4th and 6th month, different lowercase letters indicate significant differences (P<0.05) between the 2 diets in the same temperature regime; different capital letters indicate significant differences (P<0.05) among 3 temperatures in the same feed type condition. At 15, the data for months 0, 2 and 4 were from non-molt lobsters; at 6th month, all previously sampled lobsters had molted. *indicates serum protein values of post-molted lobsters.
In addition to temperature and diet type, the lobster serum protein concentration changed over time for both diet types (Two way RM ANOVA, time, F=15.19, P<0.001; interaction of temperature and time, F=16.63, P<0.001). At 5°C, the serum protein concentration was maintained at stable levels during the first 4 months, but increased significantly during the last 2 months of the study (Tukey post-hoc test, P<0.05). At 10°C, the serum protein concentration increased significantly at each two month sampling period, reaching its highest level at the end of the 6 month experimental period (Tukey post-hoc test, P<0.001). Serum protein concentrations at 15°C also increased significantly during the first 4 months, thereafter a significant decrease in serum protein concentration occurred. This was due to lower serum protein levels measured in post-moulted lobsters (Tukey post-hoc test, P<0.001).
Haemolymph samples were collected at weekly intervals in both pre and post-moult animals in 15°C (Figure 1). Serum protein concentrations (both diet types) increased steadily during intermoult and early proecdysis, reaching a peak in late proecdysis. There was a trend for the lobsters fed a mixed diet to exhibit higher serum protein levels than those fed the mussel diet, however, this difference proved to be statistically insignificant (Linear regression, F=2.98, P=0.088). The majority of the lobsters (n=33) moulted during the 22nd week. Following moulting, serum protein levels dropped to their lowest levels of between 3.55 g and 3.91 g/100 ml. Thereafter there was a significant effect of diet type on serum protein levels. Serum protein concentration slowly increased over the following 12 weeks in lobsters fed a mixed diet (Two way RM ANOVA, F=6.6, P<0.001). In contrast, serum protein levels of lobsters fed a mussel only diet declined steadily reaching levels that were significantly lower than those of the mixed diet animals after 30 weeks (Two way RM ANOVA, diet type, F=56.0, P<0.001; time, F=28.34, P<0.001; interaction, F=2.14, P<.01). Eighty nine percent (n=19) of post-moulted lobsters fed on a mussel diet died, while only 7% of post-moulted lobsters (n=14) fed a mixed diet died during the same time period.
Lab experiment 2: Feeding frequency and compartment size
There was a significant variation in ambient water temperature during the 6 month experimental period (Figure 6). Water temperature increased from around 5°C at the start of the experiment in early June, reaching maximal levels of approximately 15.5°C at the end of July. The temperature remained steady for around 3 months, after which the water temperature decreased to 10°C in early October. There was a further decrease from late October onwards, reaching the lowest measured temperature of 2°C in mid-December.
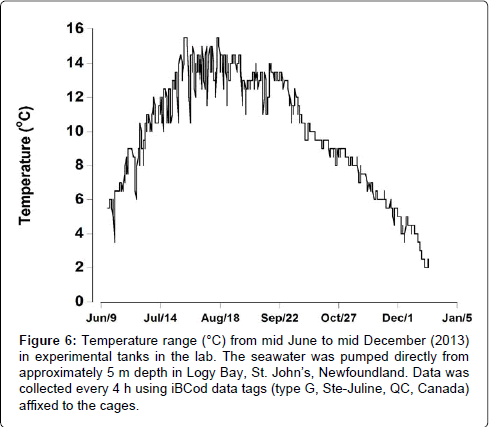
Figure 6: Temperature range (°C) from mid June to mid December (2013) in experimental tanks in the lab. The seawater was pumped directly from approximately 5 m depth in Logy Bay, St. John’s, Newfoundland. Data was collected every 4 h using iBCod data tags (type G, Ste-Juline, QC, Canada) affixed to the cages.
The mortality rate ranged between 12.5% and 37.5% and a large proportion of these mortalities occurred in last 60 days in all treatments (Table 5). There was no significant effect of feeding frequency or compartment size on mortality rates (Mantel-Cox Test, Chi square=6.588, P=0.086). Feeding frequency and compartment size did not have any significant effect on moulting rate, which ranged from 20.8% to 37.5% among the different treatments (Table 5) (Mantel-Cox Test, Chi square=0.09, P=0.761). Moulting started in mid-August, peaked during September to October and declined substantially during November and December.
| Feed Frequency/ Compartment Size |
Parameter | Number | Cumulative Mortality and Moulting (Number and Percent) Over 6 Months | |||||
|---|---|---|---|---|---|---|---|---|
| July | August | September | October | November | December | |||
| 1 Feeding/Month Large |
Mortality Moulting | 24 | 0 (0) 0 (0) |
0 (0) 2 (8.3%) | 0 (0) 4 (16.7%) | 0 (0) 4 (16.7%) | 1 (4.2%) 5 (20.8%) |
3 (12.5%) 5 (20.8%) |
| 1 Feeding/Month Small |
Mortality Moulting | 24 | 0 (0) 0 (0) |
1 (4.2%) 1 (4.2%) |
1 (4.2%) 4 (16.7%) |
1 (4.2%) 5 (20.8%) |
2 (8.3%) 8 (33.3%) |
3 (12.5%) 9 (37.5%) |
| 2 Feeding/Week Large |
Mortality Moulting | 24 | 1 (4.2%) 0 (0) | 3 (12.5%) 2 (8.3%) |
3 (12.5%) 3 (12.5%) |
3 (12.5%) 5 (20.8%) |
5 (20.8%) 6 (25%) |
9 (37.5%) 7 (29.2%) |
| 2 Feeding/Week Small |
Mortality Moulting | 24 | 0 (0) 0 (0) |
1 (4.2%) 1 (4.2%) |
1 (4.2%) 2 (8.3%) |
1 (4.2%) 2 (8.3%) |
3 (12.5%) 4 (16.7%) |
5 (20.8%) 6 (25%) |
Table 5: Mortality and moulting rates of adult lobsters held in the lab from June to December 2013 on an ambient temperature cycle. The animals were held in 2 different compartment sizes and fed either twice per week or once per month. Monthly data are shown as cumulative numbers and these are expressed as a percentage of the total number in parentheses.
Feeding frequency and to a lesser degree compartment size did have a significant effect on growth of moulted lobsters (Figures 7A-7C). Lobsters fed in the high feeding frequency treatment had a significantly higher increment of both body mass and carapace length (37.07% ± 10.94% and 10.03% ± 2.07%, respectively) compared with lobsters in the low feeding frequency treatment (20.49% ± 7.39% and 6.85% ± 1.84%, respectively) in both compartment sizes (Two way ANOVA, F=22.11, P<0.001; F=13.03, P<0.01 for body mass and carapace length respectively). Lobsters maintained in large compartments (both frequent and infrequent feeding) exhibited a trend of a larger increment in body mass but this was only statistically significant for body mass in the low feeding frequency treatment (Two way ANOVA, F=7.61, P<0.05). There was also a change in body mass for non-moulted lobsters. Lobsters in the low feeding frequency could not maintain their body mass and it decreased on average by 1.9% during the 6 month trial. Lobsters in high feeding frequency treatment maintained their body mass with a mean increase of 0.5%. However, the effect of feeding frequency on non-moulted lobster growth was only statistically significant in the small compartment sizes (Two way ANOVA, feeding frequency, F=20.91, P<0.001).
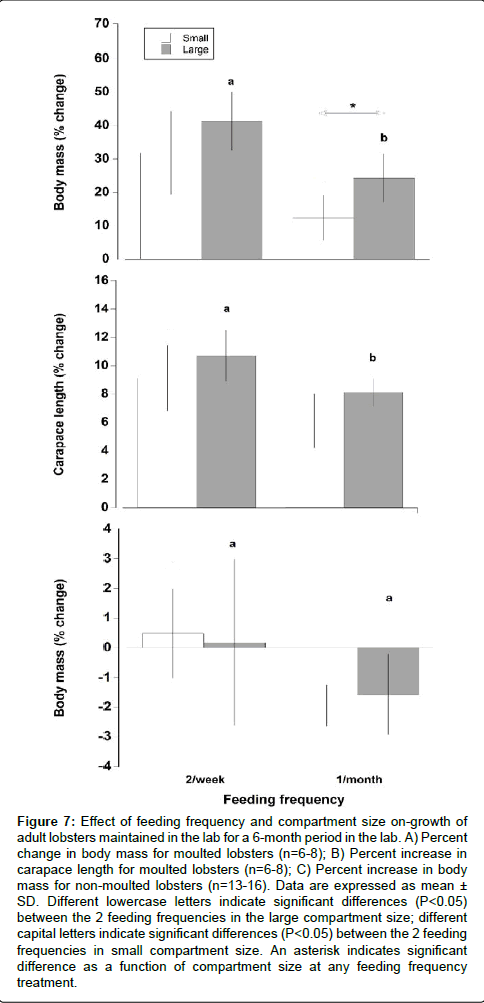
Figure 7: Effect of feeding frequency and compartment size on-growth of adult lobsters maintained in the lab for a 6-month period in the lab. A) Percent change in body mass for moulted lobsters (n=6-8); B) Percent increase in carapace length for moulted lobsters (n=6-8); C) Percent increase in body mass for non-moulted lobsters (n=13-16). Data are expressed as mean ± SD. Different lowercase letters indicate significant differences (P< 0.05) between the 2 feeding frequencies in the large compartment size; different capital letters indicate significant differences (P< 0.05) between the 2 feeding frequencies in small compartment size. An asterisk indicates significant difference as a function of compartment size at any feeding frequency treatment.
The serum protein concentration of the lobsters was significantly impacted by feeding frequency, but was not affected by the compartment size (Table 6) (Two way RM ANOVA, feeding frequency, F=54.81, P<0.001; compartment size, F=1.89, P=0.175). Non-moulted lobsters with a high feeding frequency exhibited an increase in serum protein levels during the 6 month period. The highest levels were measured in mid-October and although serum protein levels decreased slightly in the last 2 months, they were still significantly higher (7.08 g ± 2.06 g/100 ml) than levels measured at the start of the experiment (5.26 g ± 0.92 g/100 ml) (Student t test, T=3.533, P<0.01). In contrast, the lobsters fed once per month were unable to maintain serum protein levels and they declined significantly from initial levels of 5.58 g ± 0.94 g/100 ml, reaching their lowest levels of 3.79 g ± 1.09 g/100 ml at the end of the experimental period (Student t test, T=9.54, P<0.001).
| Feed Frequency | Compartment Size | Serum Protein (g/100 ml) Over 6 Months Mean ± SD |
|||
|---|---|---|---|---|---|
| June | August | October | December | ||
| 1 Feeding/Month | Large | 5.63 ± 0.97 | 4.86 ± 1.10 | 4.13 ± 1.05 | 3.81 ± 1.11aB |
| 1 Feeding/Month | Small | 5.52 ± 0.94 (8.75 ± 1.37, n=2) |
4.74 ± 1.06 (3.70 ± 1.14, n=2) |
4.08 ± 1.14 (2.07 ± 0.58, n=2) |
3.76 ± 1.12aB (1.84 ± 0.25, n=2)B |
| 2 Feeding/Week | Large | 5.40 ± 0.36 | 5.25 ± 0.67 | 7.46 ± 2.20 | 6.24 ± 2.33Aa |
| 2 Feeding/Week | Small | 5.17 ± 1.13 (8.68 ± 0.35, n=3) |
5.90 ± 1.04 (3.63 ± 0.21, n=3) |
7.65 ± 1.82 (6.03 ± 1.03, n=3) |
7.56 ± 1.80aA (5.84 ± 0.49, n=3)A |
Table 6: Serum protein concentration of adult lobsters held in the lab in two compartment sizes and on 2 feeding schedules over a 6 month period on an ambient temperature cycle. The data represents the mean + SD of 15-24 individuals at each time point. Different lowercase letters indicate significant differences (P<0.05) between the 2 compartment sizes in the same feeding frequency conditions; different capital letters indicate significant differences (P<0.05) between 2 feeding frequencies for the same compartment size. Data in parentheses are from molted lobsters.
Because of a large difference in the timing of the moult for individual lobsters and the close relationship between time after moulting and serum protein levels (Figure 1), there were not enough replicates to perform reliable statistically analysis. Nevertheless, the trend for moulted lobsters was consistent with the non-moulted lobsters. In the low feeding frequency group, the serum protein concentration of moulted lobsters decreased from 3.7 g ± 1.15 g/100 ml down to 1.84 g ± 0.25 g/100 ml (n=2) at the end of the experimental period. In the high feeding frequency group, the moulted lobster’s serum protein level increased from 3.63 g ± 0.21 g/100 ml, reaching levels as high as 6.03 g ± 1.03 g/100 ml (n=3) after 2 months.
Survival
The survival rates of adult lobsters stored in benthic cages under mussel farms were very high (>95%). In commercial holding facilities and during live transport, chilling coma (<1°C) is used to enhance survival rates [29,30]. The water temperature in the Triton area varied between 2°C-7°C, thus this relatively low temperature may have enhanced survival. However, the results of the laboratory experiments did not fully support this assumption. Lobsters held in the lab at 5°C exhibited a high mortality rate (>50%), especially those that were fed the mussel only diet. In the lab the haemolymph protein concentration decreased with decreasing water temperature (Table 3). The haemolymph contains important proteins which are involved in the immune response [31,32] and in lobsters, the rate of phagocytosis is positively related to temperature [33]. The reasons for the higher mortality in the lab are unclear. Although the experimental tanks were supplied with a constant flow of seawater and were cleaned weekly, there is the potential “wall effect” where bacterial build-up occurs on flat surfaces in these semi-enclosed laboratory systems [34-36]. Since serum protein concentrations were low in the animals maintained in 5°C, it would suggest that they may have compromised defense mechanisms, leaving them more vulnerable to infection from pathogens. Rao et al. also report higher mortality rates in tank versus cage-held spiny lobsters Panulirus homarus (Linnaeus) and suggest that this may be due to higher levels of stress in tank held animals [37]. This highlights some of the potential problems of extrapolating responses in the lab with those in the field.
There was also a high mortality rate in the lab at 15°C, but this was primarily for post moulted lobsters. Although mortality rates increase during moulting, and recently moulted lobsters are more physiologically sensitive and vulnerable [26,38,39], this was not the case here. The mortalities primarily occurred between 31 and 80 days after moulting and nearly all of them were lobsters fed the mussel diet at 15°C. The drop in serum protein levels in the lobsters fed mussels indicated that this diet was not sufficient to maintain health [25]. Amino acids such as asparagine, alanine and glutamic acid are deficient in mussels [40,41]. Astaxanthin is also lacking in the mussel diet and this plays an important role in immunocompetence and stress tolerance in crustaceans [42,43]. Post-moulted lobsters also required a higher levels of calcium intake for hardening of the shell [44]. This suggests the mussel only diet was not sufficient to provide nutrients for post-moult processes such as laying down muscle and hardening of the carapace and for dealing with increased pathogen loads in the experimental tanks.
The effects of a restricted diet (mussel only) diet on post-moult survival could be a potential concern when holding lobsters under mussel farms. The lobsters held under culture lines were likely feeding on mussels because large numbers of empty mussel shells were found in and around the cages when they were retrieved by the divers. However, the remains of gastropods, sea urchins and sea stars were also found inside the cages and green algae were growing on the cages. As lobsters are omnivorous, it was likely they were also feeding opportunistically on animals that entered the cages and therefore were not feeding exclusively on mussels, but rather getting a broad range of nutrients in their diet.
Moulting
The low moulting rate in Triton area (13%) was probably due to the cooler water temperatures (2°C to 7°C). In the lab, moulting rate was also very low in 5°C (4%) and most animals remained in the intermoult stage throughout the 6 month experimental period. The European lobster H. gammarus moults when the temperature reaches between 12°C-14°C [45]. The present results suggest that for H. americanus, rather than needing to be exposed to a certain temperature to induce moulting, the lobsters might need to be exposed for a number of degree days. The growing degree day (thermal integral) is used as a reliable predictor of growth and development in fish species and this likely also applies to crustacean moulting and growth [46].
The low moulting rate observed at Triton could also be due to food limitation since changes in food abundance impact moulting frequency in larval and juvenile stages of clawed, rock and spiny lobsters [47-49]. Crustaceans can refrain from moulting during starvation in order to save energy to maintain basal metabolic functions [50]. However, this did not appear to be the case here for adult lobsters. Lobsters in the lab fed once per month had similar moulting rates to those fed twice weekly. In addition, lobsters maintained in the laboratory at 5°C and fed had similar low moulting rates to animals at Triton where food input was limited. Crustaceans expend energy at moult and the hepatopancreas functions as a major source of energy during moulting [51]. Even though the hepatopancreas was smaller in infrequently fed animals and those at Triton it suggests that the lipid and glyceride stores would still have been sufficient to facilitate moulting [25].
Growth
The growth increment of moulted lobsters at Triton was lower than those maintained in the lab. The cooler temperatures suppress the lobster’s metabolism [52,53], and subsequently the lobsters would have consumed less food. In support of this, post moult lobsters fed frequently in the lab were significantly larger than infrequently fed lobsters. Lobsters with access to enough food would have enough reserves to lay down more muscle tissue and have energy adequate energy reserves to produce larger organs.
The effect of feeding frequency on-growth of non-moulted lobsters was somewhat different. Although one feeding per month was adequate to keep the lobsters alive, the mass of non-moulted lobsters in the lab experiment tended to decrease. In contrast, the lobsters at Triton (without artificial feeding) were able to maintain or even increase their body mass slightly. The lower temperatures at Triton probably slowed the lobsters metabolic rate and use of stored nutrients [27,54]. In spite of this, the lobsters at Triton had a more pronounced decrease in hepatopancreas size and edible meat content than those fed once per month in the lab [25]. During starvation, crustaceans metabolize their tissues, resulting in a decrease in organ mass [55,56]. One possible explanation for the starved lobsters at Triton area maintaining or even slightly increasing (1% to 2%) their body mass is that an increased water uptake would compensate for the decrease in organ mass. The body mass of white shrimp Litopenaeus vannamei (Boone) and king crabs Lithodes santolla (Molina) also remains constant during short-term starvation and is likely due to an increased water content in the body [50,57,58].
There was no effect of doubling cage size on growth of adult lobsters in the field or in the lab. In contrast, juvenile H. americanus respond to an increase in container size with a significant increase in carapace length and body mass [8,59,60]. The cages used by Beal and Protopopescu’s were large enough for juvenile lobsters to freely move around and the large surface area for settling organisms supplied plenty of food for the lobsters suggesting that in the current experiments a much larger increase in cage size relative to adult body size would be required in order to have any discernible effect [8].
Health and physiological condition
The serum protein concentration is a rapid and effective way of determining the quality and physiological condition of lobsters. There is a strong positive correlation between serum protein concentration and hepatopancreas size, heart size and edible meat content and a negative correlation with moisture content of the hepatopancreas and muscle tissue [25].
The final serum protein concentrations of the lobsters at Triton area were lower than any of the lab treatments, although the colder water temperatures may have contributed, the decrease would primarily be related to the lower food input, because lobsters maintained in the lab at 5°C and fed regularly exhibited an increase in serum protein concentration. In support of this assumption, the decrease in serum protein concentration was greater for lobsters set on open sea bottoms where they would not get the input of mussels. Interestingly, lobsters held in corner compartments had higher serum protein concentrations than lobsters held in the centre compartments. The corner compartments had a larger surface area in direct contact area with the surrounding environment, allowing more surface area to forage and these lobsters would be the first to come into contact with any organic material that drifted into the cages.
The slow increase in serum protein during post moult represents body tissue growth, which replaces the water [55,61]. Serum protein concentration declined in post-moulted lobsters in 15°C fed mussels, but increased in those fed the mixed diet. Post-moulted lobsters appeared to have a poor appetite for the mussel diet, while those fed the mixed diet continued feeding. Low serum protein concentration from the mussel diet could also be attributed the lower energetic content of molluscs when compared with other benthic invertebrates [40]. This suggests that although mussels are readily eaten by lobsters [62] and are a good source of calcium for exoskeleton hardening, they lack all the essential nutrients needed for survival [40,42,63]. This may be an important consideration when attempting to hold lobsters under mussel farms and additional feeding may be required for post-moulted lobsters if they are destined for market.
Inshore benthic storage cages could be useful in remote areas with short fishing seasons, enabling harvesters to hold lobsters and release them when market price dictates. Survival rates in the field will likely be high; the deeper cold water reduces a lobsters metabolism and need for food, thus extending their storage time [52,53]. After 3 months serum protein levels were still relatively high, indicating a healthy, quality product [25]. Taste tests showed that although people could discern a difference between the cage-held lobsters from Triton and store bought lobsters, there was no preference for either type [64]. Nevertheless, the benthic cage method may be limited for longer term storage (>3 months) and on-growth. Although cold water enhances survival [52], it reduces moulting (growth rate) and overall quality (edible meat content). After 6 months of storage the lobsters had a low serum protein concentration and were more susceptible to the effects of emersion during transport to market [24,25]. Longer term storage success could be remedied with supplemental feeding of the lobsters similar to that employed with on-growth of spiny lobsters [14]. However, the number of mussel farm sites in Newfoundland that are suitable for on-growth may be limited because the mussels lines are typically situated in deep, colder water (Laura Halfyard, Sunrise Fish Farms pers comm). Results from the laboratory experiments suggest that warmer shallow water sites typical of those found in Prince Edward Island or New Brunswick mussel aquaculture operations would be most effective at promoting moulting, and size at moult could be enhanced with supplementary feeding.
Multitrophic integrated aquaculture has typically focused on the use of bivalves and seaweeds to remove particulate and dissolved organic material around finfish farms [65-67]. There have also been recent advances in the use of sea cucumbers and sea urchins as potential vectors to control benthic deposits [68-70]. The present study showed that lobsters will survive in the vicinity of mussel farms, and when divers retrieved the cages they were full of broken mussel shells; both these factors suggest lobsters have the potential to be incorporated into a multitrophic system. Due to logistics associated with the remote location of the site, we were unable to fully assess the amount and frequency of mussel drop-off into the cages. Future work will be aimed at quantifying the amount of mussel drop-off, as well as the number of lobsters required to significantly impact the removal of benthic mussel deposits.
This research was supported by a NSERC Discovery Grant and a grant from the Canadian Centre for Fisheries Innovation (Memorial University) to IJM. We thank the staff from the workshop, dive team and the Joe Brown Aquatic Research Building (JBARB) (Ocean Sciences, Memorial University) for assistance with experiments. We thank the Halfyard family and the staff of Sunrise Fish Farms for access to their mussel farms. We also thank Stewart Lamont of Tangier Lobster, NS for providing helpful discussion on holding conditions for commercial lobsters.