Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2014) Volume 2, Issue 2
Forages tend to accumulate elevated levels of nitrate when fields are heavily fertilized with nitrogenous fertilizers or are environmentally stressed due to drought, cold, frost, hail, etc. Elevated levels of nitrate have detrimental effects on animal health and are regarded as a causative factor for several mass cattle-death incidents. It was observed that nitrate concentration in Sorghum bicolor L. obtained from local fields of Uttar Pradesh (Ghaziabad and Meerut) and Haryana (Gurgaon and Faridabad) exceeded the safe limit (2500 mg of nitrate kg-1 of fresh wt.) in a significant number of samples (31.7%) studied. Given this, the investigation was conducted in earthen pots to determine nitrate contents in 16 genotypes of S. bicolor L. A significant difference in nitrate content was observed among genotypes, many of which accumulated nitrate to toxic levels. POP-52 (V9), a high nitrate reductase (HNR) genotype and EB-15 (V7), a low nitrate reductase (LNR) genotype of sorghum were selected to study the effect of potassium (K) application on nitrate accumulation in specially designed PVC-drums. The minimum nitrate concentration (V9= 816.6 mg/kg fresh wt. and V7= 2691.8 mg/kg fresh wt.), coupled with maximum NR activity (V9= 9.916 μmol NO2 –1 h–1 g–1 fresh wt. and V7= 5.018 μmol NO2 –1 h–1 g–1 fresh wt.) were observed in 60 day old K60 treated plants. K application reduced the nitrate concentration by 35.24% in V9 and by 25.54% in V7 genotypes by increasing nitrate reductase (NR) activity by 86.23 % in V9 and lesser increase of 32.07% in V7 genotype of sorghum at 30 days. A two- fold (approx.) decrease in nitrate concentration was observed at K60 in both genotypes from 30 to 60-days-after-sowing. K application also reduced considerably the nitrate in the leachate indicating that K is effective in mitigating nitrate pollution in plants and soil. The data emphasizes the importance of K in increasing the nitrogen use efficiency and of balanced fertilization in combating the nitrate-related implications on human beings, animals and environment.
Keywords: Balanced fertilization; Forages; Genotypes; Nitrate toxicity; Potassium, Sorghum bicolor L
Agricultural development relies largely on the production and consumption of fertilizers; however, addition of fertilizers does not ensure the enhanced crop production. Excessive input of fertilizers and poor application methods can cause fertilizer movement into the ground and surface waters [1]. In order to feed the increasing population and maintain a sustainable development, the amount of nitrogen (N) to be used should be double that of the amount currently in use, if the N use efficacy of crops is not going to be improved [2]. The wrong notion of achieving increased growth and yield by applying excessive N-fertilizers alone, with inadequate supply of other nutrients, especially potassium, has led to imbalanced fertilization in the developing countries [3-6].
Nitrate is the main form of N to be taken up and assimilated by annual crops [7]. In many parts of the world, nitrate concentrations in ground water exceed the maximum limit of 50 mg l-1, equivalent to 11.3 mg l–1 as NO3– -N as recommended by the WHO [8]. Intensive agricultural production, domestic and industrial wastes, sewage and atmospheric nitrogen pollution are the main sources of nitrate contamination in water [9]. Inefficient use of available N leads to accumulation of nitrate in plants that causes toxicity to animals, which feed on them. When the input of nitrogen exceeds the demand, plants are no longer able to absorb it, and N then builds up in the soil, mostly as nitrates, and leaches into the groundwater [10,11]. Nitrate leaching, due to excessive N fertilization, leads to eutrophication of freshwater bodies [12,13] and the marine ecosystems [14,15]. Nitrate (NO3-) accumulation in plant tissue is usually observed during drought, long periods of cloudy or cool weather, or following a heavy fertilization with manures and nitrogen-containing fertilizers and due to herbicide applications [16]. Anything that slows down the rate of plant growth can lead to increased nitrate levels in well-fertilized plants. Nitrate per se is not very toxic to animals. Inside the body, nitrates are converted to nitrites and then to nitrosamines that result in gastric cancer and other diseases in the consumers [17]. Methemoglobinemia is another adverse effect of nitrate poisoning [18,19].
Increase in N at the cost of decrease in K has major repercussion on the utilization of N itself. The depletion of K in agricultural ecosystems has, in fact, prevented increases in crop yields [4,5,20]. K is an essential mineral element for plant growth and development [21] and plays a key role at various points in N metabolism. Zhang et al. [3] reported that rapid nitrate uptake depends upon adequate K in the soil solution. The significance of potassium in affecting the uptake, translocation and reduction of NO3− is well documented by Ruiz and Romero [22]. Shrotriya [23] reported that balanced application of N, P and K might cause up to 122% increase in sorghum yield in India. Therefore, improving the crop performance and wisely handling the nutrient management are prerequisites for optimizing crop production and minimizing environmental risks due to N losses.
Sorghum is capable to accumulate toxic levels of nitrate even at moderate N fertility levels [24]. It is the fifth most important cereal crop grown in the world and is also valued for its fodder and stover [25]. According to Mahanta and Pachauri [26] in India, forage sorghum is grown in 2.6 m ha mainly in the states of western UP, Haryana, Punjab, Rajasthan and Delhi, which fulfill over two third of the fodder demand during Kharif (summer) season.
In order to minimize the risk of nitrate poisoning in ruminants and protect the soil and underground water from potential nitrate pollution without compromising with the yield, a balanced fertilization with application of K and maintaining a proper harvesting time may be beneficial [27]. The present study was undertaken on sorghum to investigate the role of K in increasing the utilization of applied nitrogen to increase plant growth and to prevent the environmental contamination.
Growth conditions and materials
Sorghum leaf samples (15 in number) were randomly collected from agricultural fields located at different districts in the states of Uttar Pradesh (Ghaziabad and Meerut) and Haryana (Gurgaon and Faridabad) just prior to an thesis. Leaf nitrate concentration was estimated in the fresh samples so as to evaluate their fitness for intake by livestock with reference to safe limits propounded by Evans et al. [28]; according to which, 0-2500 mg/kg of NO3-1 is the safe nitrate limit to feed all the classes of livestock, while 2500-5000 mg/kg of NO3-1 is the caution limit to feed the pregnant and young animals. Moreover, 5000-15,000 mg/kg of NO3-1 is considered as the dangerous toxic limit, while over 15,000 mg/kg of NO3-1 is extremely toxic and should not be fed to animals under any circumstances. Based on the results obtained further experiment was conducted in the Herbal Garden of Jamia Hamdard, New Delhi, during the kharif (summer) season of 2010- 2011. A pot-culture experiment was carried out on sixteen genotypes of sorghum (Sorghum bicolor L.) under naturally illuminated green house (photosynthetically activity radiation [PAR] >960 μmol m2 s-1, day/night temperature 25/20 ± 4°C and relative humidity of 70 ± 5%) in the Department of Botany, Hamdard University, New Delhi, India (28°38´ N, 77°11´ E and 228 m altitude). Authentic seeds were procured from Sorghum Research Institute, Hyderabad (India) (Table 1). The experiment was conducted in 10 inches diameter earthen pots according to randomized block design using three replicates in order to screen the genotypes of sorghum on the basis of NR activity and nitrate concentration in the leaves. Prior to sowing, the pots were lined with polythene bags provided with holes thereafter, the pots were filled with 8 kg soil taken from the Herbal Garden of Jamia Hamdard, New Delhi. The soil belonged to the Lukhi soil series of Gurgaon. It was sandy loam (83.6% sand, 6.8% silt and 9.6% clay) with a neutral pH 7.1. The available nitrogen (30 mg kg-1), phosphorus (4 mg kg-1) and potassium (40 mg kg-1) contents of the soil were low. The soil was air-dried, passed through 4 mm mesh for removing plant litter and mixed thoroughly. On the basis of soil test recommendations, a uniform basal dressing of fertilizers comprising nitrogen (120 mg kg-1), phosphorus (30 mg kg-1), potassium (80 mg kg-1) and zinc (25 mg kg-1) was applied to the soil before sowing. Sources of N, P, K and Zn were urea, single super phosphate, muriate of potash (KCl) and zinc sulphate, respectively. Fresh leaf samples were analyzed simultaneously for NR activity and nitrate concentration at 30 days after sowing (DAS). Two sorghum genotypes, viz. POP-52 and EB-15, were identified as high and low NRA genotypes, respectively, which were employed for further studies with various levels of potassium.
| Sl. no. | Code no. | Accession no. |
|---|---|---|
| 1 | V1 | CSV 15 |
| 2 | V2 | CSV 21F |
| 3 | V3 | CSV23 |
| 4 | V4 | E-68-1 |
| 5 | V5 | E73 |
| 6 | V6 | E77 |
| 7 | V7 | EB-15 |
| 8 | V8 | HC-308 |
| 9 | V9 | POP-52 |
| 10 | V10 | SPSSV 5 |
| 11 | V11 | SPSSV 6 |
| 12 | V12 | SPSSV 7 |
| 13 | V13 | SPSSV 20 |
| 14 | V14 | SPSS 422 |
| 15 | V15 | SPV-462 |
| 16 | V16 | SPV-913 |
Table 1: Accession numbers and codes used in the study for Sorghum bicolor L. genotypes obtained from Sorghum Research Institute, Hyderabad.
The high and low NRA genotypes were sown in 25 cm×120 cm (diameter × height) PVC drums with a total capacity of 60 liters. The experiment was conducted using the same soil as that of the pot experiment obtained from the Herbal Garden of Jamia Hamdard, New Delhi (India). The experimental drums had drainage system at three different places (30, 60 and 100-cm height of the soil column). The lower opening (drainage system) of the drum was filled with glass wool, followed by that at 10 cm height filled with washed fine gravels. Plastic funnels (10 cm diameter) with PVC tubes (5 mm diameter) were fitted with each drain to collect the leachate. The funnels were filled with washed fine sand after putting a piece of glass wool in the lower opening of the funnel. The Herbal Garden soil was filled in the drums above 10 cm gravel layer to reach the height of 100 cm after the saturation process for the filled soil. Before planting, the soil was irrigated to 20% water content. Ten seeds per pot were sown. The pots were weeded and scarified weekly. The experiment was laid out with randomized block design using three replications. The schematic diagram of the experimental set up is given (Figure 1).
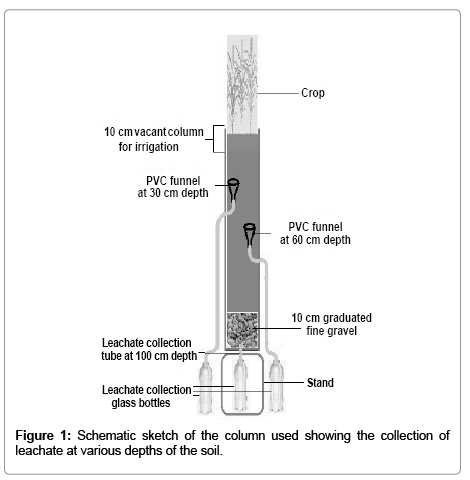
Figure 1: Schematic sketch of the column used showing the collection of leachate at various depths of the soil.
PVC drum experiment treatments
In addition to the uniform basal dressing of N, P and Zn, four levels of potassium, viz.0, 30, 60 and 120 mg K2O kg-1 soil (K0, K30, K60 and K120, respectively), as muriate of potash, were applied prior to sowing.
Sampling in PVC pot experiment
The youngest fully expanded leaves from the same position were sampled in triplicates from each treatment pot at two different stages, viz. 30 and 60 DAS. The leaf-tissue, lacking midrib, was used to determine NRA and nitrate concentration.
Determination of leaf nitrate concentration
Nitrate was extracted using the method of Grover et al. [29]. Ground dried leaf material (100 mg) was taken to which, 50 mg activated charcoal and 50 ml distilled water were added and boiled on a hot plate for 10 minutes. This was filtered through Whatman No. 42 filter paper, allowed to cool to room temperature and the volume was made to 100ml. Downe’s improved reduction hydrazine method [30] was adopted for the estimation of nitrate. An aliquot of 0.6 ml of the sample was drawn, mixed with 0.1 ml each of catalyst (0.0354 g CuSO4.5H2O + 0.9 g ZnSO4.7H2O in one liter of water), sodium hydroxide (1N) and hydrazine sulphate (1.71 g l-1) solution in a sequence. The reduction time was 7.5 min at a temperature of 33°C. Subsequent to reduction of nitrate to nitrite, it was diazotized by the addition of 0.32 ml of sulphanilamide and 0.1 ml of naphthyl ethylene diamine dihydrochloride and the pink colored solution was read in a spectrophotometer at 540 nm. The nitrate concentration was expressed as mg g-1 fresh weight of leaves.
Determination of in- vivo NRA
Random sampling of the youngest fully extended fresh leaves was carried out to estimate the activity of NR (E.C. 1.6.6.1) The NRA in the leaves was determined by the intact tissue assay method of Jaworski [31]. Chopped leaf pieces (200 mg) were incubated for 2 h at 30oC in a 5.5 mL reaction mixture, which contained 2.5 mL of 0.1 M phosphate buffer, 0.5 mL of 0.2 M potassium nitrate, and 2.5 mL of 5% isopropanol. The nitrite formed was subsequently determined by using the spectrophotometer recording the OD at 540 nm after azo-coupling with sulphanilamide and naphthylene diamine dihydrochloride. The enzyme activity was expressed as μmol NO2 h-1g-1 FW.
Determination of biomass
Fresh weight (g/plant) in triplicates was measured using a digital balance and the mean values obtained. This was followed by ovendrying the plants at 65°C ± 2°C for 48 hours to constant weight in order to record the corresponding plant dry weight (g/plant).
Determination of biomass nitrogen utilization efficiency
Whole plant (stem + leaf) nitrogen concentration (WPNC, %) was determined by using carbon- hydrogen- nitrogen-sulphur (CHNS) analyzer (Elementar, Analytic, Jena, Germany). Total nitrogen content (g plant-1) was calculated by multiplying whole plant N concentration with plant biomass (g plant-1) and dividing it by100 [i.e. (WPNC × plant biomass)/100]. Biomass nitrogen utilization efficiency was calculated using the formula: NutE = Biomass (dry matter) (g plant-1)/Total N content (g plant-1).
Collection and analysis of leachate
Each time, the leachate was collected after irrigating the plants; it was used for the analysis of content of N, using PC-based UV-vis double beam spectrophotometer (Systronics 2202) at 210 nm [32,33].
Statistical analysis
Data were analyzed statistically and given as mean ± Standard Error (SE). Analysis of variance was performed by SPSS (16.0 Inc., USA). Duncan multiple range tests (DMRT) were performed to test the significance of the differences between treatment means at a threshold P value of 0.05. Simple linear correlation and regression analysis was carried out using MS Excel 2007.
Leaf nitrate levels in the farmers’ fields
The range and mean values of nitrate concentrations in the leaf samples of Sorghum bicolor L., collected from the farmers’ fields located in the districts of Uttar Pradesh (Ghaziabad and Meerut) and Haryana (Gurgaon and Faridabad), are summarized in Table 2. On fresh weight basis, the nitrate content in the leaf-samples ranged from 3500 to 7102 mg kg-1 (Ghaziabad), 1549 to 4981 mg kg-1 (Meerut), 528 to 2822 mg kg-1 (Gurgaon), and from 794 to 2919 mg kg-1 (Faridabad). The highest nitrate concentration was recorded in Ghaziabad samples (7102 mg kg- 1). This is far beyond the safe limit of nitrate concentration (2500 mg kg-1) reported in the forage crops [28]. Nitrate concentration in Meerut samples (4981 mg kg-1) was also relatively high, almost reaching the caution limit of 5000 mg kg-1. On the other hand, the samples from Gurgaon and Faridabad were lower in nitrate concentrations, though all of them were not safe for consumption as some of them exceeded the safe limit. Thus, many of the collected forage samples were not suitable for consumption by the animals as they could result in toxicity.
| Location | Number of samples | NO3(mg kg-1fresh wt of samples) Mean (range) |
|---|---|---|
| Ghaziabad | 15 | 5316 (3500-7102) |
| Meerut | 15 | 2769 (1549-4981) |
| Gurgaon | 15 | 1621 (528-2822) |
| Faridabad | 15 | 1771 (794-2912) |
Table 2: Nitrate concentration in fodder samples collected from different districts of
Uttar Pradesh and Haryana.
Pot experiment results
Leaf nitrate concentrations in 30 days old plants of sorghum genotypes: Of the sixteen sorghum genotypes grown in earthen pots, leaf nitrate concentrations varied significantly in 30 days old plants (Figure 2). The minimum and maximum nitrate concentration was 2308.67 and 7121.3 mg kg-1 fresh wt. noted in V9 (POP-52) and V7 (EB-15) genotypes, respectively. Leaf nitrate concentration of three sorghum genotypes V2 (CSV 21F), V9 (POP-52) and V13 (SPSSV 20) were under safe limit (below 2500 mg of nitrate kg-1 fresh wt.), while seven of the genotypes V1 (CSV 15), V3 (CSV 23), V4 (E-68-1), V8 (HC-308), V11 (SPSS V6), V14 (SPSS 422) and V16 (SPV-913) were not fit for feeding the pregnant and young animals as they reached the caution limit of 2500 - 5000 mg of nitrate kg-1 fresh wt. Out of the genotypes under investigation, six genotypes V5 (E-73), V6 (E-77), V7 (EB-15), V10 (SPSS V5), V12 (SPSS V7) and V15 (SPV-462) exceeded the safe limit and were categorized under the dangerous toxic level (5000 - 15,000 mg of nitrate kg-1 fresh wt.).
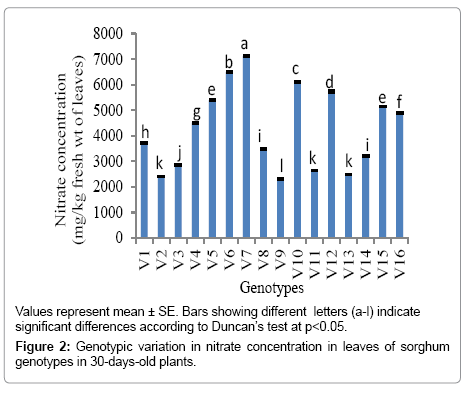
Figure 2: Genotypic variation in nitrate concentration in leaves of sorghum genotypes in 30-days-old plants.
Leaf NR activity in 30 days old plants of sorghum genotypes: Genotypic variation was evident in the leaf NR activity in 30 days old plants (Figure 3). V9 genotype had the highest level of NR activity (5.99 μmol NO2–1 h–1 g–1 fresh wt.) with minimum nitrate concentration, while the minimum level of NR activity was observed in V7 genotype (1.813 μmol NO2–1 h–1 g–1 fresh wt.). There was a significant negative relationship (r = -0.913) between level of NR activity and leaf nitrate concentration among the sixteen genotypes (Figure 4).
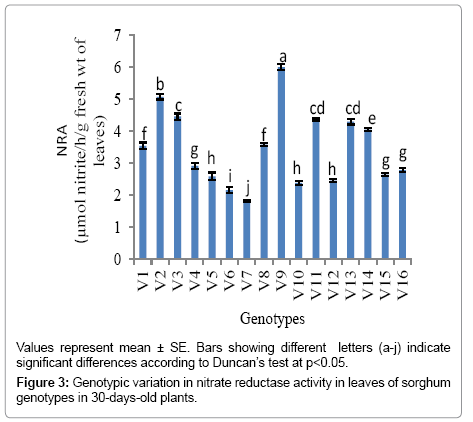
Figure 3: Genotypic variation in nitrate reductase activity in leaves of sorghum genotypes in 30-days-old plants.
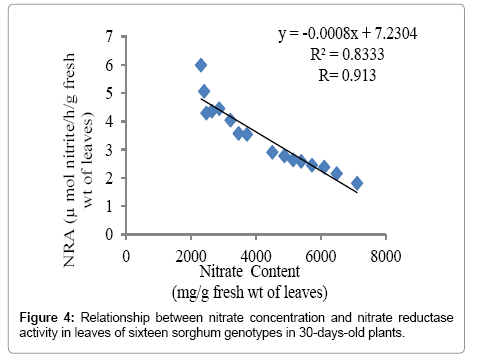
Figure 4: Relationship between nitrate concentration and nitrate reductase activity in leaves of sixteen sorghum genotypes in 30-days-old plants.
Leaf nitrate concentration under potassium application: Application of potassium decreased the nitrate accumulation significantly (p<0.05) in the leaves of both the sorghum genotypes studied (V9 and V7 genotypes). The minimum and maximum nitrate concentration was associated with the application of K60 and K0, respectively. In V9 genotype, there was recorded 2118.89 mg of nitrate kg-1 fresh wt. at K60; whereas at K0, it was 3272.04 mg of nitrate kg-1 fresh wt. In V7 genotype, there was 7104.06 mg of nitrate kg-1 fresh wt. at K60, while it was 9600.97 mg of nitrate kg-1 fresh wt. at K0 in 30 days old plants (Figure 5). However, this decrease in nitrate concentration with potassium application increased maximally only at K60, after which the decrease was substantial. The result showed that application of K60 brought the nitrate concentration under safe limit only in V9 genotype at 30 DAS.
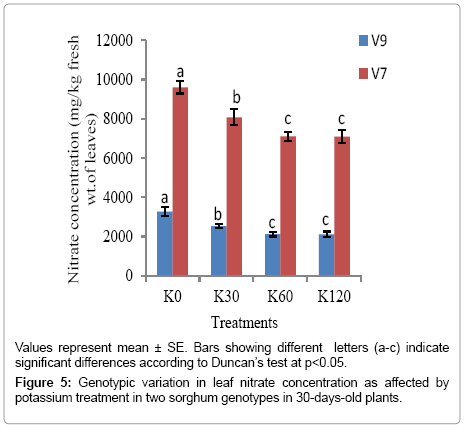
Figure 5: Genotypic variation in leaf nitrate concentration as affected by potassium treatment in two sorghum genotypes in 30-days-old plants.
The nitrate concentrations in the leaves were lower at 60 DAS than at 30 DAS (varying from 1.68- 3 folds). The lowest nitrate concentration, occurring at K60 in V9 genotype (816.6 mg of nitrate kg-1 fresh wt.) was safe for animal consumption. V7 genotype was however, not safe for consumption under natural conditions in spite of the considerable reduction in nitrate concentration with K application (Figure 6).
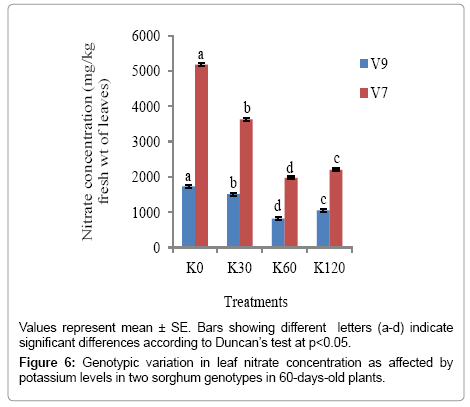
Figure 6: Genotypic variation in leaf nitrate concentration as affected by potassium levels in two sorghum genotypes in 60-days-old plants.
Leaf NRA under potassium application: There was a positive and linear relationship between NR activity and K-fertilization rate up to K60 in both the genotypes at both the stages, after which the increase was not distinct. The increase in NR activity was significant at (p<0.05) with K application. In 30-day old plants, the maximum NR activity appeared with K60 in V9 genotype (5.978 μmol NO2-1 h-1 g-1 fresh wt of leaves), the genotype that carried the minimum leaf nitrate content. Similar result was obtained in 60-day old plants as well, with the maximum NR activity being attained in V9 genotype (9.916 μmol NO2-1 h-1 g-1 fresh wt of leaves) in comparison to V7 genotype (5.018 μmol NO2-1 h-1 g-1 fresh wt of leaves). Minimum level of NR activity was obtained in the control (K0) in 30 as well as 60 days old plants of both the genotypes (Figures 7 and 8).
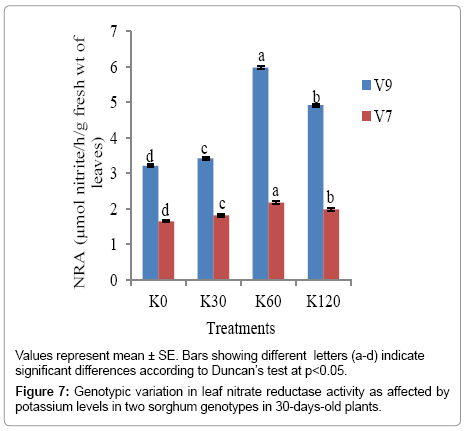
Figure 7: Genotypic variation in leaf nitrate reductase activity as affected by potassium levels in two sorghum genotypes in 30-days-old plants.
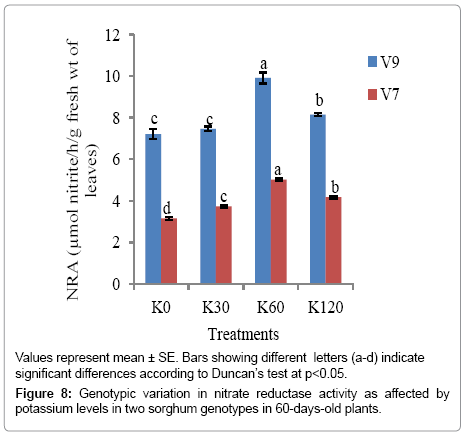
Figure 8: Genotypic variation in nitrate reductase activity as affected by potassium levels in two sorghum genotypes in 60-days-old plants.
Plant biomass
Genotypic variation was also evident in plant biomass (fresh and dry weight) of the sorghum plants at 60 DAS and was significant at (p<0.05) (Figure 9). The fresh weight was the highest with K60 (689.12 g plant-1) in V9 as compared to V7 genotype at K0 (356.85 g plant-1). A profound difference in dry weight between the two genotypes was also noted with the application of potassium. The dry weight was significantly lower in V7 genotype at all the K levels (Figure 10).
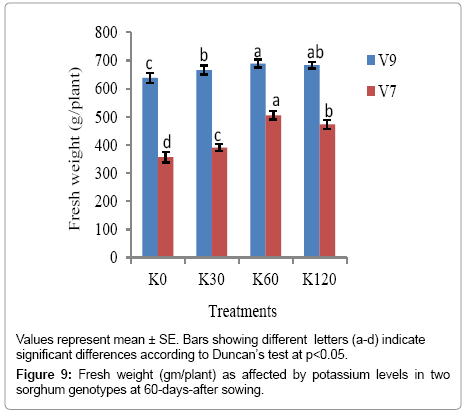
Figure 9: Fresh weight (gm/plant) as affected by potassium levels in two sorghum genotypes at 60-days-after sowing.
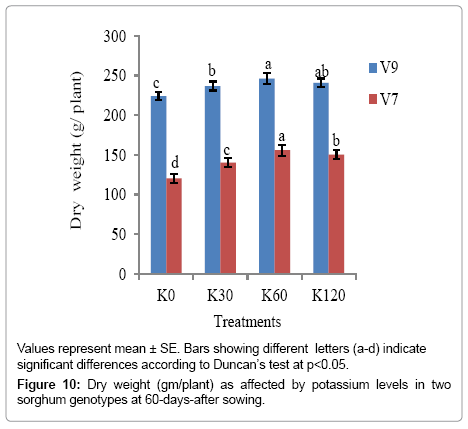
Figure 10: Dry weight (gm/plant) as affected by potassium levels in two sorghum genotypes at 60-days-after sowing.
Biomass nitrogen utilization efficiency in leaves at 60 DAS
The effect of potassium was apparent on the biomass nitrogen utilization efficiency in leaves after 60 days of growth (Figure 11). V9 had better biomass nitrogen utilization efficiency in comparison with V7 genotype at all the levels of potassium. There was a significant (p<0.05) increasing trend from K0 to K60 in both the genotypes; however, the increase was not evident at K120. The maximum biomass nitrogen utilization efficiency was recorded at K60 in V9 (45.93) and V7 genotypes (39.09), while the minimum biomass nitrogen utilization efficiency was recorded at K0 both in V9 (37.1) and V7 (31.05).
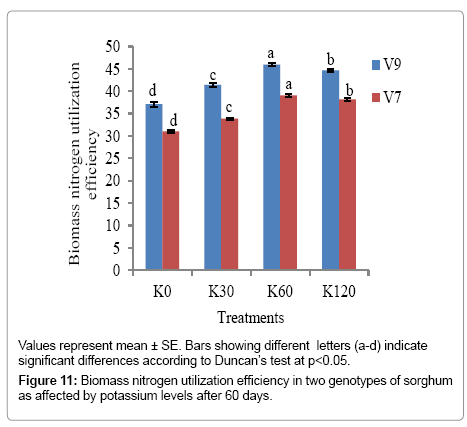
Figure 11: Biomass nitrogen utilization efficiency in two genotypes of sorghum as affected by potassium levels after 60 days.
Nitrate content in the leachate
As evident from Figures 12-14, nitrate content in leachate decreased significantly (p<0.05) with increase in K levels (from K0 to K60) at all the soil depths. The minimum nitrate content was recorded with K60 in both the genotypes at all the soil depths. However, V9 exhibited comparatively a higher reduction in nitrate content in the leachate, suggesting that nitrate had been used by the plants.
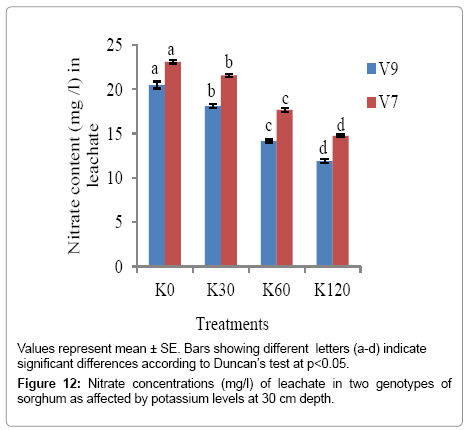
Figure 12: Nitrate concentrations (mg/l) of leachate in two genotypes of sorghum as affected by potassium levels at 30 cm depth.
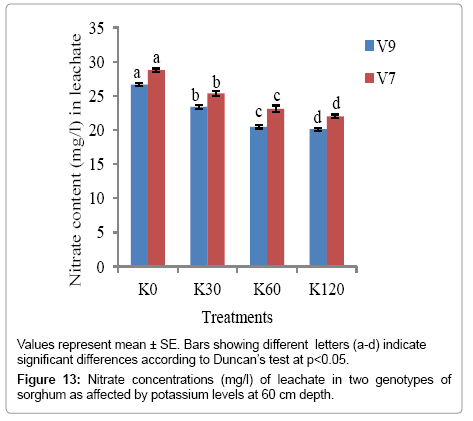
Figure 13: Nitrate concentrations (mg/l) of leachate in two genotypes of sorghum as affected by potassium levels at 60 cm depth.
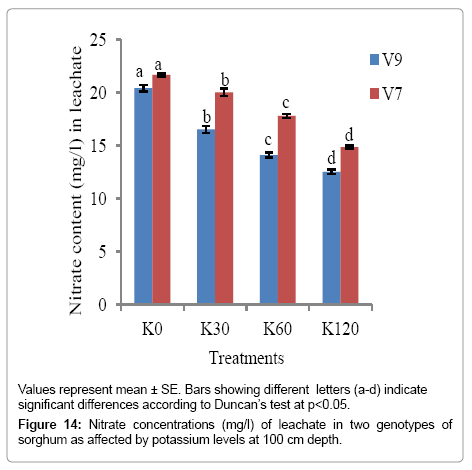
Figure 14: Nitrate concentrations (mg/l) of leachate in two genotypes of sorghum as affected by potassium levels at 100 cm depth.
Nitrogen is an essential element in biological materials and nitrate is the most common form of N taken up by the plants [34]. Changes in the availability and metabolism of N are of particular importance for maintaining high productivity of crops [35]. However, excessive use of nitrogenous fertilizers in order to attain maximum yield is leading to accumulation of nitrates above the safe limits causing toxicity to ruminants [36]. Nitrate is easily leached because it shows negligible interaction with negatively charged matrix of most topsoil and is therefore very mobile in the soil [37]. Further, loss of nutrients due to intensive agricultural practice is of great concern as it may contribute to a variety of secondary effects in the soil, plant and environment [38]. The sandy soils, with minimal nutrient retention capacity, needs attention in this regard. The nutrients not absorbed by the plant roots in these soils tend to move freely with soil-water resulting in their leaching to deeper soil strata. In the present study it was observed that nitrate content in sorghum exceeded the safe limit in 13 out of the 16 genotypes studied as shown in the Figure 2. The leachates also showed the presence of nitrate.
Mondal and Nad [39] reported that nitrate assimilation in forage crops is an acute problem, causing toxicity to animals that feed on them; it may accumulate in plant parts to toxic levels due to generous nitrogen application. The inter- and intra- species variations in leaf nitrate content have been reported in many crop plants [40]. These variations may occur due to difference in the efficiency of nutrient uptake system and nitrate reduction system of plants or a combination of both, or due to genetic differences among genotypes in the N assimilation pathway. The N assimilation pathway might differ due to differences in the rate of NO3 uptake and the uptake of other elements needed for enzyme activities, or to the differences in the generation of electron donors needed by the nitrate assimilation pathway [36].
To reduce nitrate accumulation in plants its efficient utilization is required. Strategies for improving nitrogen assimilation require the knowledge of N-assimilation pathway. NR activity is assumed to be the rate-limiting step for nitrate assimilation in plants [41,42]; hence, there exists a close relationship between NR activity and nitrate concentration in plants [43]. Nitrate is reduced to nitrite by NR activity in plant system. Consequently, there is a reverse relationship between nitrate content and NR activity [44]. The findings of our study also showed a negative relationship between NR activity and nitrate concentration in leaves (Figure 4).
On comparison of the nitrate content and NR activity in the two genotypes, it was observed that the increased NR activity decreased the nitrate accumulation in the leaves of sorghum at both the growth stages (Figures 7 and 8). Further, NR activity increased and nitrate content in the leaves decreased with plant age. It has been reported that the NR activity varies with the plant species, age and growth conditions [45,46]. Kapoor and Paul [47] studied that in potato tuber the NR activity began to decline at about 50 days after emergence, and this may be a useful indicator of when to apply nitrogen fertilizer so as to increase tuber protein content.
This study as indicated in Figures 5 and 6 also suggests that potassium application reduced the nitrate accumulation in leaves, which is in accordance with the findings of Ahmed et al. [48] and Ruiz and Romero [22]. They reported that increase in the rate of potassium application facilitated the uptake and transport of nitrate towards the aerial parts of the plant, promoted the metabolism and utilization of nitrate and ultimately, reduced the nitrate accumulation in plants. Phosphorus, potassium and sulfur have major roles in the production of proteins and, hence, their consumption in plants results in the decrease in nitrate content in the plants [49]. Therefore, deficiency of plant nutrients seems to favor accumulation of nitrate in plants, whereas integrated use of the fertilizers of potassium, phosphorus and N may improve nitrogen uptake as well as the N- assimilation efficiency. Studies have demonstrated that interaction between N and other nutrients, in particular, potassium and phosphorus impacts N efficiency and crop yield. Adequate and balanced use of fertilizer nutrients is the most effective way to improve the efficiency of N fertilizers [50]. Potassium effectively reduced nitrate content and enhanced maximally the activity of NR and the corresponding lowest nitrate content at K60 in both the genotypes. The difference in nitrate and NR activity in different treatments was apparent at both the growth stages.
The biomass nitrogen utilization efficiency in the studied samples also increased considerably with potassium application indicating that K played an important role in improving nitrogen use efficiency of plants by enhancing the NR activity as NR activity has been reported to be augmented by potassium application [51].
Singh et al. [52] reported that the rate and the total amount of the leached ions have been shown to be affected by a number of factors. One of them is the use of unbalanced N fertilization that leads to accumulation of nitrate in the soil beyond the safe limit. Various strategies are suggested in order to decrease leaching or increase the use efficiency of nutrients. These include application of different fertilizers in different modes, avoiding runoff, mitigating losses from soil and plants, using slow release fertilizers and nitrification inhibitors, use of organic or green manures, use of legumes in cropping systems, correction in their imbalanced use and integrated nutrient management. Among these measures, use of judicial K fertilization might be an efficient strategy to increase N assimilation and prevent the nitrate leaching. Brar et al. [53] reported that application of K effectively increased the NUE of maize. Brar and Imas [54] studied the effect of K fertilization on N uptake and efficiency in various crops and suggested that balanced fertilization with K increases crop yield while enhancing NUE for the protection of the environment. A higher NUE implies better utilization of N and therefore lesser wastage. Improving NUE with K implies that lesser N can be applied without affecting yield and thereby preventing land and water contamination. In the present study, nitrate content in the leachate decreased considerably with the increase in the level of potassium. The minimum nitrate content was recorded with K60 level in both the genotypes. This trend was consistent at all the soil depths, viz. at 30, 60 and 100 cm below the soil surface as depicted in Figures 12-14. The decrease in nitrate content in the leachate would have been due to better utilization of the available nitrate with K application. The potassium-nitrogen partnership is important for getting maximum N returns [55]. Increased K fertilization reduces nitrate accumulation in plants [56-58]. Silva et al. [59] suggested that the interaction of rates of N and K significantly affects the activity of NR. The application of double rate of K and phosphorus contributed to a reduction in nitrate content of rocket plants even at high rates of N [60]. Increasing amount of K caused reduction in N-nitrate in rocket leaves [61]. The present study focusses on the importance of K in increasing NUE and decreasing the nitrate content in plants to prevent nitrate toxicity in human and nitrate pollution of environment.
The samples from the local fields as well as those from experimental sorghum genotypes showed a significant accumulation of nitrate, exceeding the safe limit (2500 mg kg-1 fresh wt.). Nitrate concentration in the leaves was the highest at the younger stage (30 DAS). However, with K application, especially with K60, leaf nitrate concentration decreased considerably, while the NR activity appeared at the highest level in both the genotypes (V9 and V7). Genotype V9 exhibited better growth seemingly through increased NR activity and decreased nitrate concentration in leaves and leachate compared to V7 at both the growth stages. The maximum K application effect appeared after 60 days of growth. Thus, a balanced nutrient management with the K application at 60 mg K2O kg-1 soil (K60) could help in reducing nitrate accumulation and sustaining growth and productivity of sorghum. This would help in minimizing the nitrate toxicity in animals and its load on ground water. Further, this would help the farmer to attain a cost effective strategy of fertilizer management without wastage of fertilizers. This should be validated, however, with further studies at large scale employing different crops at various locations with different climatic conditions.
We gratefully acknowledge Prof. M. Iqbal, Dept. of Botany, Jamia Hamdard and Dr. Patricia Imas, for their assistance in shaping the manuscript. Generous help rendered by the Sorghum Research Institute, Hyderabad, in providing seeds of sorghum is gratefully acknowledged. The authors also thank the International Potash Institute (IPI), Switzerland, for funding this study.