Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2015) Volume 3, Issue 2
The detriment of chemical fungicides and pesticides have proven a hindrance in sustainable agriculture and therefore motivated efforts in finding biologically natural alternatives to control plant pathogen infections. The effect of the plant growth promoting rhizobacteria (PGPR), Paenibacillus polymyxa, Bacillus licheniformis and Bradyrhizobium japonicum IRAT FA3 on seed germination and their biocontrol capability against Pseudomonas syringae pv malculiola in Arabidopsis thaliana were studied in this work. The data presented here demonstrates that the presence of these PGPR promotes the seed germination rate up to 145% higher than the untreated control. This study also describes the symptom isolation observed in Arabidopsis leaves infected with P. syringae, which prevented the infected plants to undergo full-fledged chlorosis suggesting that these PGPR can be possible options in acting as natural biocontrol agents against P. syringae and probably other plant pathogens.
Keywords: PGPR, Phytohormone production, Germination, Pathogenic pressure
A plant’s rhizosphere is home to numerous microbiota. Of these microbial populations, a promising aspect in farming sustainability is the advantageous relationships established between plants and plant growth promoting rhizobacteria (PGPR). These microbes colonize the root of the plant due to leaking photosynthates from the roots; 5-21% of the carbon fixed by plants is secreted as root exudates [1,2] such as L-leucine in tomatoes, which acts as a chemo attractant for Pseudomonas fluorescens WCS365 prompting its colonization of the tomato root [3]. Spanning several genera in the bacterial domain, such as Pseudomonas, Bacillus, Bradyrhizobium, Azospirillum among numerous others, are microorganisms that have both direct and indirect effects on plants. Known roles attributing to growth promotion are the formation of nodules, increased root development which further contributes to increased water and mineral uptake, solubilization of phosophates, siderophore production, phytohormone stimulation, biomass increase, stress tolerance, volatile compound production, signal interference, such as inhibition of biofilm formation, induced systemic resistance, competitive exclusion with pathogenic microbiota and production of antagonistic compounds such as antibiotics [4-15]. Pseudomonas syringae, is a well-known phytopathogen that exhibits spreading of chlorosis on leaves and elicits hypersensitive responses in its host plants, such as Arabidopsis thaliana [16]. It is known to infect a wide range of plant species. The pathovar used in this study, Pseudomonas syringae pv Malculiola ES4326’s virulence is rpoN dependent and its mode of attack is the synthesis of the phytotoxin, cornonatine [16]. Various approaches have been used in curbing phytopathogens such as P. syringae ES4326 and a suggested avenue in doing so is using soil-dwelling bacteria that do not pose a threat to the plant but can disseminate the pathogenic effect via competitive exclusion, production of inhibitory or bactericidal-specific compounds and/or induced systemic resistance [17]. Previously known as Bacillus polymyxa [18], Paenibacillus polymyxa is known to have a role in plant growth promotion due to its nitrogen-fixing capabilities [19], its role in phosphorus solubilization from the soil [20], cytokinin production [21], increased soil porosity [22] and production of compounds that attribute to the enhancing the biocontrol capabilities of plants such as production of hydrolytic enzymes [23]. Bradyrhizobium japonicum’s functional role in promoting root nodulation in leguminous plants has been well studied, to which the bacteria is responsive to root exudates which form Nod factors and the interaction between the root and bacteria resulting in nodulation [24]. Various strains have been seen to yield larger plants with increased productivity [13,24], stimulation of phytohormone production such as indole-3-acetic acid, zeatin, gibberellic acid, ethylene, abscisic acid [25], solubilization of phosphorous [26] and abiotic stress tolerance such as drought [13] and salinity [14]. Bacillus licheniformis’s mechanism of growth promotion effect has been well-studied in alder plants (Alnus glutinosa [L.] Gaertn) and highlighted to be responsible for stimulation of root nodulation [27], siderophore and cellulose production [28] and phytohormone production such as gibberellin [29] and auxin [28]. The biocontrol capabilities of these plant growth promoting rhizobacteria have not been fully investigated against P. syringae, a prevalent phytopathogen that attacks a vast array of hosts resulting in shoot and flower blights, cankers and diebacks in woody plants. This pathogen is easily spread by natural vectors such as wind-driven rain, insects and farming practices that involve the accumulation of water in the midst of harvesting [29,30]. Biocontrol technologies such as use of PGPR could prove beneficial and foster the next frontier in sustainable farming whereby handling of phytopathogens can be accomplished without chemical detriment [31]. Additionally, the effect of these plant growth-promoting rhizobacteria on a faster germination rate was investigated in this study.
Plant material and growth conditions
Arabidopsis thaliana seeds, ecotype Col-0, from the Arabidopsis Biological Research Center (ARBC) were used in this study. The seeds were sown in soil in 8 cm wide clay pots and left to grow in gnotobiotic greenhouse conditions of 12 h/12 h day/night at a temperature of 25°C. The soil was autoclaved twice for 60 minutes at 121°C, 15 psi within a 24-hour period to eliminate the influence of pre-existing microflora on the outcome of the study.
Inoculum preparation and plant inoculation
Pure inoculum of B. japonicum IRAT FA3, P. polymxa and B. licheniformis, isolated from maize rhizospheres, were acquired from 3-day old Luria-Broth (LB) liquid cultures grown at 28°C under shaking conditions at 120 rpm. Each bacterial inoculum’s concentration was maintained at 108 cfu/ml. Prior to sowing of seeds, the soil for each treatment was soaked in its respective inoculum, whereas the control treatment soil was soaked in water.
Administration of virulent Pseudomonas syringae pv maculicola ES4326 to plants
For the pathogen infection, following the procedure elucidated by Barth et al. [32] and Gachomo et al. [33], 2 week-old seedlings of each treatment were infected with P. syringae pv maculicola ES4326.
Experimental design and statistical analysis
The experiments were completely randomized in nature and each experiment comprised of nine replicates. Data were expressed as mean values ± SE. P values were determined by One-way ANOVA analysis using Graph Pad Prism 6 Software for Mac OS X.
Arabidopsis plants typically germinate within 3-5 days. However, the A. thaliana plants that were treated with P. polymyxa, B. licheniformis and B. japonicum IRAT FA3 exhibited early germination in comparison to the untreated control. The plants treated with B.licheniformis averaged germination at 1.8 days, whereas those treated with P.polymyxa germinated at 2 days. B. japonicum IRAT FA3 had an average germination time span of 2.8 days and the untreated control had one of 4 days (Figure 1). This yielded the statistically significant value of p<0.0001 obtained from the One-Way ANOVA statistical test for the bacteria-treated plants.
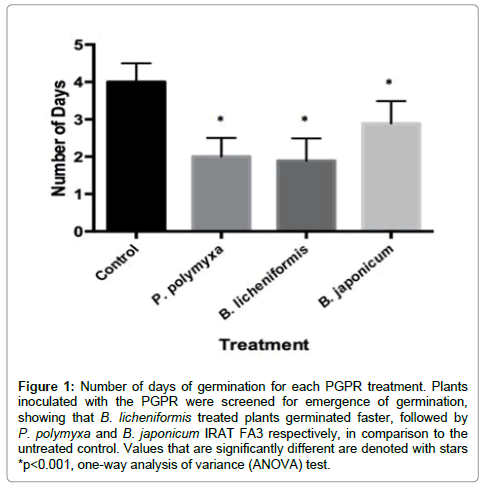
Figure 1: Number of days of germination for each PGPR treatment. Plants inoculated with the PGPR were screened for emergence of germination, showing that B. licheniformis treated plants germinated faster, followed by P. polymyxa and B. japonicum IRAT FA3 respectively, in comparison to the untreated control. Values that are significantly different are denoted with stars *p< 0.001, one-way analysis of variance (ANOVA) test.
The A. thaliana plants treated with P.polymyxa, B. licheniformis and B. japonicum IRAT FA3 were significantly larger in size, as indicated by (Figures 2 and 3), where the plants were imaged at 1 week post sowing. The B. licheniformis treatment yielded the largest plants of all the treatments, followed by P. polymyxa and B. japonicum IRAT FA3. We noticed that all three bacteria-treated plants looked 2-3 weeks old when in essence they were one week old, which is indicatory of a faster growth rate. Additionally, the leaves in all the PGPR-treated plants were significantly larger in comparison to the untreated control (Figures 4 and 5), whereby the B. licheniformis leaves were the largest of the treatments, followed by P. polymyxa then B. japonicum. In the face of pathogenic pressure, all three PGPR treatments did yield plants with larger leaf surface areas, which also displayed symptom isolation at the final stages of the disease, indicated by the speckled yellowing, unlike the untreated control. B. japonicum IRAT FA3, however, yielded plants with the largest leaf surface area of the three PGPR treatments (Figure 6) displaying that it promotes significant growth despite being under attack. The plants from the untreated control displayed a 100% infection in its leaves. The PGPR-treated plants, however, displayed symptomic isolation.

Figure 2: PGPR treated plants yielded plants further in their leaf development stage. PGPR-inoculated plants displayed a faster development rate, with oneweek old plants appearing as if they are 2 weeks old with B. japonicum IRAT FA3 and 3 weeks old for those treated with B.licheniformis and P. polymyxa.
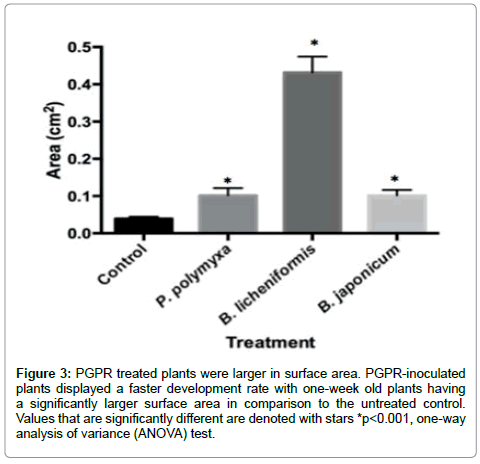
Figure 3: PGPR treated plants were larger in surface area. PGPR-inoculated plants displayed a faster development rate with one-week old plants having a significantly larger surface area in comparison to the untreated control. Values that are significantly different are denoted with stars *p< 0.001, one-way analysis of variance (ANOVA) test.

Figure 4: PGPR treated plants resulted in leaves with larger surface areas.3a: A: Bradyrhizobium japonicum IRAT FA3, B: Paenibacillus polymyxa, C: Bacillus licheniformis. Plants inoculated with the PGPR yielded leaves with larger surface areas in comparison to the untreated control. The 4-week old leaves display that B. licheniformis yielded the plants with the largest surface areas followed by P. polymyxa then B. japonicum IRAT FA3.

Figure 5: PGPR treated plants resulted in leaves with larger surface areas. PGPR-inoculated plants yielded leaves with larger surface areas in comparison to the untreated control. Values that are significantly different are denoted with stars *p< 0.001, one-way analysis of variance (ANOVA) test.

Figure 6: PGPR treated plants maintained symptom isolation in leaves. A: Bradyrhizobium japonicum IRAT FA3, B: Paenibacillus polymyxa, C: Bacillus licheniformis. 4-week old PGPR-inoculated plants isolated the symptoms of the P. syringae pv. maculicola in the leaves whilst promoting further growth of the plants as depicted by the larger leaves, whereas the control underwent fully fledged chlorosis. B. japonicum IRAT FA3 promoted the most growth despite being under pathogenic pressure followed by B. licheniformis and P. polymyxa.
P. polymyxa had an infection rate of 37.74% of its total leaf surface area. The infection on the leaves of the B. licheniformis-treated plants displayed an infection that comprised 25.63% of the total leaf surface area. B. japonicum displayed the best symptom isolation capability with the chlorosis displayed on the leaf making up 21.38% of the total leaf surface area. It is evident that the PGPR-treated plants yielded larger plants. P. polymyxa, as clarified by Linderg et al. [34], displayed that the capability of nitrogen fixation holds no direct correlation to the rate of growth promotion. However, phytohormone stimulation of cytokinin has been observed [20]. Cytokinins, often found in low concentrations, possess crucial and significant roles in the growth and development of a plant. Such roles include the promotion of seed germination, leaf size expansion, and stimulation of reproductive development, senescence retardation, de novo bud release and formation [20,35]. P. polymyxa’s ability to synthesize cytokinins is most likely what resulted in the faster germination rate of the plants. Additionally, the large leaf surface area of the A. thaliana, whose soil was treated with P. polymyxa, could be attributed in the cytokinin synthesis that occurs in the presence of these bacteria. The antagonistic capability of P. polymyxa as observed against oomycetic pathogens namely Phytophthora palmivora and Pythium aphanidermatum [36] could be due to the bacteria colonizing the root tip of the plant and forming biofilms as well as entering intercellular spaces but not spreading through the plant as studied by Timmusk et al. [37]. Additionally, it was shown that P. polymyxa produce hydrolytic enzymes in barley rhizospheres antagonizing pathogenic soil-dwelling fungal populations [22]. This is most likely why plants treated with P. polymyxa showed symptomic isolation, rather than a spreading chlorosis that engulfed the whole leaf as observed in the untreated control when inoculated with P. syringae.
B. licheniformis has been noted to have growth promotion capabilities as well. It was previously understood that much of the reason that plants treated by B. licheniformis yielded larger plants was due to biosynthetic levels of auxin being higher in the presence of the bacteria [38]. However, auxin is a phytohormone that attributes to seed dormancy through the stimulation of abscisic acid signaling, which plays a dominant role in seed dormancy, by the induction of the ARF-mediated (particularly the AUXIN RESPONSE FACTOR 10 and AUXIN RESPONSE FACTOR 16 pathways) ABSCISIC ACID INSENSITIVE 3 (ABI3) activation in A. thaliana [39]. Gutierrez- Manero et al. [38] saw that in alder (Alnus glutionosa [L.] Gaertn). Seedlings with a dwarf phenotype induced with the gibberillic acid (GA) biosynthesis-inhibiting compound namely, paclobutrazol yielded phenotypically normal plants when treated with B. licheniformis whilst stimulating stem elongation. The results were synonymous with plants that underwent exogenous application of GA. This is evidentiary that B. licheniformis does promote the production of GAs. Their ability to produce such phytohormone compounds is most likely what resulted in the faster germination rate of the A. thaliana plants treated with the bacteria. This suggests that the amount of auxin produced in comparison to gibberellin production in the presence of B. licheniformis is low enough that the function of GAs in breaking seed dormancy is effective, thus resulting in a faster germination rate than in the untreated control. Additionally, the bacteria’s ability to possess biocontrol properties are not extensively understood, however a study conducted by Lee et al. [40] displayed that a formulation of B. licheniformis proved to have a higher disease control value than those of chemical fungicides against Botrytis cinerea as well as having an additional contribution to promoting tomato growth. This is most likely why plants treated with B. licheniformis did not only show symptomic isolation of the Pseudomonas syringae pv maculicola ES4326 but also yielded larger plants in comparison to the untreated control.
According to Gachomo et al. [13], B. japonicum IRAT FA3 resulted in larger Zea mays (maize) and A. thaliana plants. For maize, the treatment of B. japonicum IRAT FA3 yielded larger plants with broader and greener leaves and a denser, longer root system with more root hairs in comparison to the untreated control. These then attributed to a higher shoot and ratio biomass. For A. thaliana, the plants were visibly larger however, their growth stages as exhibited by their leaves showed that those treated with B. japonicum IRAT FA3 were in the fourth growth stage while the untreated control were in the second growth stage. Additionally, they had an increased number of siliques and the soil nutrients, nitrate, ammonia and phosphorous were found to have significantly increased in the soil. Some B. japonicum strains have been known to contribute to elevated production of phytohormones, particularly gibberellin and zeatin [41,42]. Zeatin is a phytohormone that belongs to the class namely, cytokinins. Both phytohormones play roles in breaking seed dormancy and most likely attributed to the faster germination rate as well as growth promotion capability. Inoculation with a combination of Trichoderma and Bradyrhizobium spp. lead to significant increase in percentage of Phytopathora sojae inhibition and increased yield in soybean plants due to the increase in peroxidase enzymes and phytoalexin activities [43,44]. This is most likely why plants treated with B. japonicum IRAT FA3 not only showed the best symptomic isolation of the P. syringae pv maculicola ES4326 but also yielded plants with the largest surface areas despite being under pathogenic pressure.
In summary, the treatment of A. thaliana with P. polymyxa, B. licheniformis and B. japonicum IRAT FA3 proved beneficial not only in plant growth promotion, but also in reduction of symptom spreading on infected foliage. Additionally, the observance that these plant growth promoting rhizobacteria play a role in faster germination rates could explain another method that facilitates their faster growth promotion, yielding mature plants in a shorter amount of time. Our findings suggest that in hopes to limit the detriment of chemical-based biocontrol agents, the use of these soil dwelling bacteria could pose a better option to not only act as biofertilizers but as natural biocontrol agents against certain pathogens.
In conclusion, plants treated with the PGPR Paenibacillus polymyxa, Bacillus licheniformis and Bradyrhizobium japonicum IRAT FA3 promote faster growth and seed germination rate in Arabidopsis thaliana, as well as prime them to fight phytopathogens such as Pseudomonas syringae pv malculicola.