Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 2
Homeostasis of cellular Polyamines (PAs) is very important for proper growth and development of living organisms. Development of male sex organ and gametophyte in plants is a complex process involving many known and unknown pathways. PAs are actively involved in the development of anther and pollen. In the present study, oat arginine decarboxylase (ADC) gene has been over-expressed in tapetum of tomato using tapetum-specific TA29 promoter to perturb the PA homeostasis in order to see its implication on male fertility. Tapetum-specific ADC overexpression resulted in PA accumulation, mainly in free and bound fractions in the anther of transgenics. Transgenic lines displayed abnormal pollen and tapetum development. Pollen grains were aborted and distorted in morphology and tapetum showed premature degeneration in ADC transgenics. These transgenics failed to set seeds, but cross pollination with normal pollen from wild type plants restored the seed setting indicating that s ADC over-expression did not affect female fertility. ADC over-expression also did not affect the growth and morphology of transgenics. The present study suggests that the excess of PAs in anther is lethal for pollen and tapetum development and overexpression of ADC gene can be an effective approach for the engineering of male sterility in plants.
<Keywords: Polyamine, Arginine decarboxylase, Tapetum, Pollen development, Male sterility, Transgenic plants, Solanum lycopersicum
Polyamines (PAs), the low molecular weight, poly-cationic nitrogenous compounds are widely distributed in all the organisms. Putrescine (Put), Spermidine (Spd) and Spermine (Spm) are major PAs and they have been implicated in regulation of several important physiological and molecular processes including cell division, growth and development, flower and fruit development, stress responses, membrane integrity, senescence, stabilization of DNA, gene expression and many other cellular functions [1-6]. In animals and fungi, the essential Put, the precursor of the higher PAs, Spd and Spm is synthesized directly from ornithine by the enzyme Ornithine Decarboxylase (ODC). Plants and bacteria have an alternative route to the production of Put from arginine, which is catalyzed by Arginine Decarboxylase (ADC). Additional reactions convert Put into Spd and Spm. These steps are catalyzed by Spd and Spm synthases, which add propylamino group generated from S-adenosylmethionine (SAM) by SAM decarboxylase (SAMDC). Diamine oxidase (DAO) and Polyamine Oxidase (PAO) are involved in the degradation of diamines (Put) and PAs (Spd and Spm), respectively [2].
Pollen represents the entire male gametophyte. Its potential uses in genetic transformation and breeding programs have encouraged researchers to study pollen development in depth [7-9]. In flowering plants, development and maturation of pollen takes place in anthers. It starts with the meiosis of Pollen Mother Cells (PMC) to make haploid tetrad in the locule of anther. Haploid tetrad then gets released as uninucleate microspores [9] with the help of callase, an enzyme produced by tapetum. Tapetum is the innermost and most nutritive sporophytic tissue of the anther [10]. Further in the development process, uninucleate microspores undergo asymmetric division and forms bicellular pollen with a larger vegetative cell and a smaller generative cell [9]. Maturation of pollen grain is the final process in pollen development. A number of changes take place in the cytoplasm, including the deposition of reserve material and dehydration of the pollen grain. In addition, several changes occur at the surface of the pollen wall such as the deposition of substances involved in pollenpistil interaction. Various changes also occur in the nourishing layer of the anther, i.e., the tapetum to support the pollen maturation [11].
Tapetum provides various nutrients, enzymes and other molecules for the pollen development. During late pollen developmental stage, tapetum undergoes Program Cell Death (PCD) mediated degeneration (apoptosis) and releases the nutrients into anther locule. The significance of the tapetum in pollen development is evident from the facts that any abnormality in this tissue directly affects the male fertility of the plant [10]. In addition, timely apoptotic degeneration of the tapetum is also crucial for male fertility as abolition of apoptosis or premature PCD mediated degeneration [12,13] of tapetum results in male sterility. This indispensable involvement of tapetum in pollen development, encouraged the study of various tapetum- specific genes, promoters [14,15] and also elucidation of the functioning of tapetum [16-20]. Furthermore, various strategies, viz. expression of cytotoxic gene in tapetum, tapetum-specific down-regulation of genes involved in pollen development, abolition of PCD-mediated degeneration of tapetum, etc. have been adopted to genetically engineer male sterility in crops [21-25].
PAs are well associated with the anther and male gametophyte development. The necessity of the PAs for pollen maturation, germination and tube growth has been examined by various experiments using PAs and their inhibitors in in vitro germination and also ex-vivo applications [26-29]. It has also been observed that decrease in PA levels in floral organs causes male sterility [25,29-31]. Interestingly, elevated levels of PAs are also seen to be associated with abnormal stamen development [32,33], pollen degeneration [34] and delayed flowering [35]. In view of these findings, it is important to determine the role of PAs in anther and pollen development and explore their usefulness in inducing male sterility, a useful tool in breeding through hybridization.
In the present study, we examined the influence of ADC overexpression on tapetum and pollen development in tomato. To achieve this, ADC gene from oat was placed under a tapetum-specific promoter TA29 in a binary vector and used to generate tomato transgenic plants. TA29 promoter, derived from tobacco, is specifically and strongly active in anthers and its activity starts early during flower growth and disappears as the tapetum degenerates at later stages of anther development [14]. Tomato transgenic plants over-expressing ADC gene in the present study, displayed aborted and non-viable pollen, resulting in male sterility.
Plant material and growth conditions
The seeds of tomato (Solanum lycopersicum Mill. cv. Pusa Ruby) were procured from National Seeds Corporation, New Delhi. Plants were grown under controlled growth conditions (26 ± 2°C, 16 h photoperiod with irradiance of 40 μE mol m-2 s-1).
Binary plant transformation vector
For the preparation of TA29-ADC construct, pBINAR binary vector harboring neomycin phosphotransferase (NPT-II) as a plant selection marker (obtained from Dr. P. Ananda Kumar, NRC on Plant Biotechnology, New Delhi) was digested with EcoRI and Kpn I restriction enzyme and end-filled for the cloning of TA29 promoter into it. The TA29 promoter was excised out from Litmus 28/TA29 plasmid (provided by Prof. R. B. Goldberg (Department of Biology, University of California, Los Angeles, California) using NcoI-HindIII restriction enzyme and end-filled before its blunt end ligation into pBINAR vector. Oat ADC gene (2.1 kb) was digested out from pRT101 (procured from Prof. R. L. Malmberg, Department of Botany, University of Georgia, Athens), end-filled and ligated into pBINAR (harboring TA29 promoter) using SmaI restriction site downstream of TA29 promoter. In both these cases, the right orientation was confirmed. E. coli strain DH5a was used for the maintenance and multiplication of the plasmid. The Agrobacterium tumefaciens strain LBA4404 was used for tomato transformation following the mobilization of the binary vector.
Tomato transformation and regeneration
Transformations were done by co-cultivating cotyledon explants, collected from about 10-12 days old axenic seedlings with A. tumefaciens (LBA4404 stain) harboring TA29-ADC binary vector following a protocol developed by Madhulatha et al. [36]. The co-cultivated explants were then selected on shoot regeneration medium [SRM-MS medium fortified with 3% maltose, 2.5 mg/l 6-benzylaminopurine (BAP), 0.5 mg/l indole-3-acetic acid (IAA), 0.5 mM Put] containing 30 mg/l kanamycin (selection agent) and 300 mg/l augmentin (bacteriostatic agent). It was cultured for about two months with subculture every fortnight. The shoots obtained on selection medium were subjected for proliferation on MS medium fortified with 0.5 mg/l BAP. The well-grown shoots were excised and transferred to the rooting medium (half-strength MS+250 mg/l augmentin). The rooted plants were transferred to pots containing vermiculite: soil in a 1:1 ratio and plants were covered with polythene bags for a week to maintain high humidity for hardening in the tissue culture room and then transferred to green-house.
Polymerase chain reaction (PCR)
Putative tomato transformants were checked for presence of the transgenes by PCR analysis. Genomic DNA was isolated from tomato leaf tissue by CTAB method [37]. PCR was done for 35 cycles with NPT-II and ADC gene specific primers at 59°C and 53°C annealing temperature, respectively. PCR products were checked on 1% agarose gel. The primer pairs for the amplification of a 750 bp fragment of NPT-II gene were 5’-TCAGAAGAACTCGTCAAGAA-3’ and ’-ATGGGGATTGAACAAGATGG-3’and for the amplification of a 1000 bp fragment of ADC gene were ’ CGGCCACCTCTGCGTCAGAATCTAT 3’ and ’ ACGCCGTGATAGCGCACTTGAGC 3’.
Southern hybridization
Transgene integration for T0 transformants (only PCR positives) was further confirmed by Southern hybridization using oat ADC and NPT-II gene specific probes. For Southern hybridization, 10 μg genomic DNA was digested with PstI restriction enzyme and separated on 0.8% agarose gel. Southern blots were prepared employing the standard protocol [38], using nylon membrane (Hybond N, Pharmacia). The ADC and NPT-II gene probes were prepared using 32P-labeled dCTP by random primed labeling kit as per the manufacturer’s instructions (Takara, Japan). Pre-hybridization and hybridization were carried out according to Sambrook et al. [38]. The membranes were washed and then exposed to X-ray film (XK- 5, Kodak film).
RNA extraction and semi-quantitative RT-PCR
Total RNA was isolated from 10 mm long anthers as per the protocol of Chomczynski and Sacchi [39]. Anthers (150 mg) were homogenized in liquid nitrogen using mortar and pestle and 400 μl of extraction buffer (4M GITC, 1M sodium citrate, 10% sarkosyl) was added to it along with 600 μl of phenol: chloroform: isoamyl alcohol (25:24:1). The homogenates were vortexed for 30 sec and the debris were pelleted by centrifugation at 15000 x g for 20 min at 4°C. Supernatant was carefully removed and extracted twice with an equal volume of chloroform: isoamyl alcohol (24:1). Supernatant was recovered in a fresh centrifuge tube and to it 1/10th volume of sodium acetate (3 M, pH 5.2) and 0.6 volume of isopropanol was added and the mixture was then incubated at -70°C for 1 h. Total RNA was precipitated by centrifugation at 4°C for 15 min. Pellet was re-suspended in 200 μl of DEPS treated water and then 50 μl of 10 M LiCl was added and incubated overnight at 4°C and centrifuged at 15000 x g at 4°C for 30 min. The pellet was again resuspended in 200 μl of DEPC treated water and again re-precipitated using 1/10th volume of sodium acetate (3 M, pH 5.2) and 0.6 volume of isopropanol. The mixture was then incubated at -70°C for 2-3 h and centrifuged at 15000 x g at 4°C for 30 min. Pellet was washed with 70% ethanol, dried and dissolved in an appropriate volume of DEPC treated distilled water.
In order to detect oat ADC transcripts in transgenic lines and untransformed control , RT-PCR was carried out with100 ng total RNA (treated with DNaseI, Fermentas, USA) using gene- specific primers (forward primer ’ CGGCCACCTCTGCGTCAGAATCTAT 3’ and reverse primer ’ ACGCCGTGATAGCGCACTTGAGC 3’) using one step RT-PCR kit (Qiagen Inc.) following manufacturer’s instruction for 24 cycles at 53°C.
Polyamine estimation
Polyamines were extracted and analyzed by thin-layer chromatography (TLC) following Bajaj and Rajam with minor modifications [40]. PAs from 10 mm long anthers were extracted in 5% (v/v) cold perchloric acid (PCA) at a ratio of 500 mg tissue / ml PCA and centrifuged at 20000 x g for 20 min at 4°C. The pellet (containing PCA insoluble bound PAs) was re-suspended in 1 ml of 1N NaOH. Soluble conjugated and bound PAs were released from 200 μl of supernatant and pellet suspension, respectively, by acid hydrolysis with 200 μl of 12 N HCl in sealed glass ampules and incubated overnight at 110°C. The hydrolysate was filtered through glass wool in small drying vials and dried on a dry bath at 70°C for 10-15 min. The residue was dissolved in 300 μl of PCA (5%) and used for dansylation. Aliquots of 200 μl of supernatant (free PAs), supernatant hydrolysate (conjugated PAs) and the pellet hydrolysate (bound PAs) along with PA standards were treated with equal volume of dansyl chloride (5 mg/ ml acetone) overnight. After 16 h, 100 μl proline (100 mg/ml, ~87 μM) was added to each of the samples to remove excess dansyl chloride and the dansylated PAs were extracted in 100 μl of benzene, which were analyzed by TLC using ethylacetate and cyclohexane (4:5 v/v) as the solvent system for 2 h. Individual PA bands were scrapped off from the plate, re-suspended in 3 ml ethyl acetate and were analyzed by a spectroflurometer (VersaFlour TM, BioRad) at excitation wavelength of 350 nm and emission wavelength of 495 nm. Concentration of PAs was calculated by extrapolating from standard curve of known concentrations of standard PAs.
Pollen viability and structure analysis
The anthers were collected from transgenic and untransformed control tomato plants at the stage of dehiscence. Pollen viability was checked by squashing anthers in 2% acetocarmine stain. The viable pollens stained red and non-viable pollens failed to take the stain.
For Scanning Electron Microscope (SEM) and Transmission Electron Microscopy (TEM), anthers were fixed in 3% glutaraldehyde, postfixed in 1% osmium tetroxide, and dehydrated in a graded acetone series. For SEM samples were critical point dried, mounted on aluminium stubs, and sputter coated with gold. Specimens were viewed on a LEO 435 VP variable pressure scanning electron microscope at an accelerating voltage of 30 kV.
For TEM, 50-120 nm sections of the anthers were cut with using diamond knife. The sections were picked up on 70×300 mesh copper grids, and stained with uranyl acetate and lead citrate. The sections were viewed in a Phillips 300 TEM at 60 kV and images photographed on Kodak 4489 electron microscope film.
Generation of Oat-ADC tomato transgenic plants and molecular characterization
A construct was prepared by cloning TA29 tapetum-specific promoter and Oat ADC gene in pBINAR binay vector as a prerequisite for the generation of ADC over-expressing transgenic tomato plants (Figure 1A). The TA29-ADC construct was then used for A. tumefaciens mediated transformation of tomato. Thirteen putative tomato transformants were checked for presence of the transgenes by PCR analysis of the genomic DNA using ADC as well as NPT-II gene specific primers (Figure 1a and b). The only PCR positive transgenic plants were further confirmed for the transgene integration and copy number by Southern hybridization using ADC and NPT-II gene specific probes (Figure 1b- 1d).
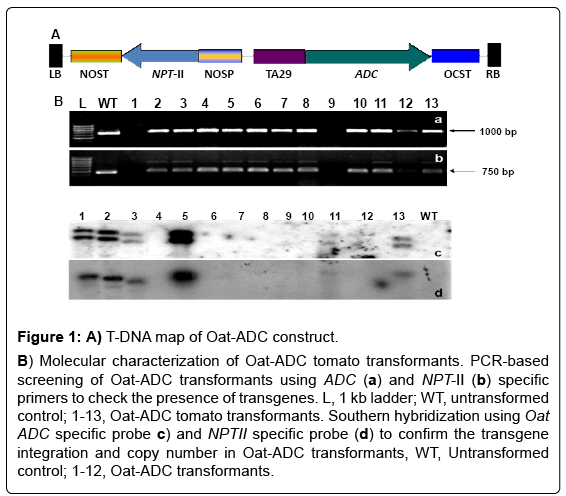
Figure 1: A) T-DNA map of Oat-ADC construct.
B) Molecular characterization of Oat-ADC tomato transformants. PCR-based screening of Oat-ADC transformants using ADC (a) and NPT-II (b) specific primers to check the presence of transgenes. L, 1 kb ladder; WT, untransformed control; 1-13, Oat-ADC tomato transformants. Southern hybridization using Oat ADC specific probe c) and NPTII specific probe (d) to confirm the transgene integration and copy number in Oat-ADC transformants, WT, Untransformed control; 1-12, Oat-ADC transformants.
Oat ADC over-expression impairs pollen viability in transgenics
Tapetum-specific expression of gene did not affect the growth and morphology of the ADC tomato transgenic plants, except pollen viability as all the transgenic plants were phenotypically similar to untransformed control plants and had no apparent morphological aberrations (Data not shown). However, the effect of ADC over-expression on pollen development became evident with the acetocarmine staining, which differentiates the red viable pollen from non-stained colorless nonviable pollen. All the transgenics were either completely (all non-viable pollen) or partially (mixed population of viable and non-viable pollen) sterile showing their pollen viability in the range of 0% - 25% (Figure 2B and 2D) compared to 90-100% pollen viability in untransformed control plants (Figure 2A and 2C). Further, pollen from ADC transgenic lines when observed under scanning electron microscope (SEM) were found to be shrunken and distorted in morphology (Figure 2F and 2H) unlike round and intact pollen of untransformed control plants (Figure 2E and 2G). The completely sterile lines failed to set fruits. Moreover, the buds turned brown and dried up after few days of development, but the female fertility was unaffected as crossing of male sterile lines with untransformed control pollen restored the fertilization resulting in fruit setting (data not shown). Partially male sterile lines had less number of fruits when compared to control. It ranged from two fruits to six fruits per plant.
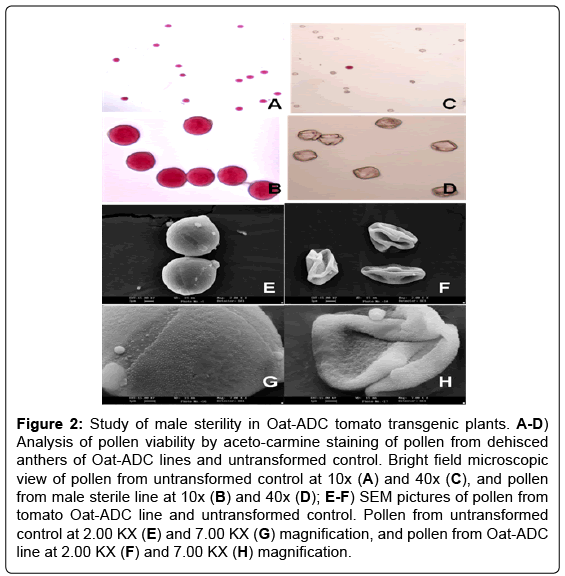
Figure 2: Study of male sterility in Oat-ADC tomato transgenic plants. A-D) Analysis of pollen viability by aceto-carmine staining of pollen from dehisced anthers of Oat-ADC lines and untransformed control. Bright field microscopic view of pollen from untransformed control at 10x (A) and 40x (C), and pollen from male sterile line at 10x (B) and 40x (D); E-F) SEM pictures of pollen from tomato Oat-ADC line and untransformed control. Pollen from untransformed control at 2.00 KX (E) and 7.00 KX (G) magnification, and pollen from Oat-ADC line at 2.00 KX (F) and 7.00 KX (H) magnification.
ADC over-expression increased polyamine titre in transgenic anthers
The transgene expression was analyzed in the anthers of three lines i.e., 2, 3 and 5 along with the untransformed control by semiquantitative RT-PCR using Oat ADC gene-specific primers. The RTPCR results showed variable expression of Oat ADC gene in different transgenic lines (Figure 3), which may be due to the position effect of transgene integration. Further, to check the effect of Oat ADC transcript accumulation on PA levels in anther, we quantified PAs in anther of ADC lines and untransformed control by TLC. The ADC transgenic lines showed increased PA levels, particularly the bound fractions of Put and Spd and free fraction of Spm (Figure 4). Free and conjugated Put concentration in ADC lines was not different from the untransformed control. However, transgenic anthers had 3-5 fold increase in bound Putas compared to non-transformed controls. Overexpression of ADC gene showed around 1.5 fold increase in the free form and 6-7 fold increase in bound form of Spd concentration, which further led to 3-4 fold increase in free and 2-fold increase in bound forms of Spm (Figure 4).
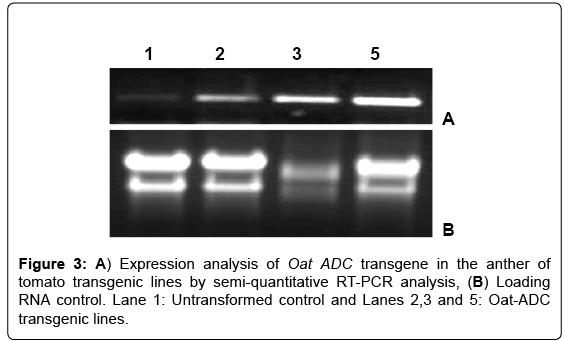
Figure 3: A) Expression analysis of Oat ADC transgene in the anther of tomato transgenic lines by semi-quantitative RT-PCR analysis, (B) Loading RNA control. Lane 1: Untransformed control and Lanes 2,3 and 5: Oat-ADC transgenic lines.
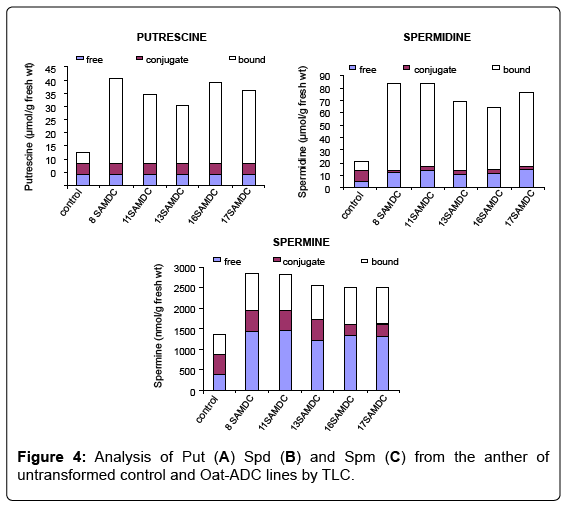
Figure 4: Analysis of Put (A) Spd (B) and Spm (C) from the anther of untransformed control and Oat-ADC lines by TLC.
PA accumulation in tapetum affected pollen development
In order to investigate the effect of ADC over-expression on microsporogenesis in the developed ADC lines, we examined pollen cellular constitution by transmission electron microscopy (TEM). The pollen from untransformed control tomato plant displayed dense cytoplasm with well developed pollen wall. On the other hand, pollen from transgenic ADC lines were found to be empty and highly vacuolated with thick pollen wall (Figure 5). Moreover, because of the empty cytoplasm pollen from transgenic lines were distorted and collapsed as observed by SEM (Figures 2F and H). The 1 μm thin transverse section (TS) of transgenic anther when observed under bright field microspore displayed aborted and lysed pollen grains at the anthesis stage in comparison to the round and fully developed pollen in untransformed control plants (data not shown).
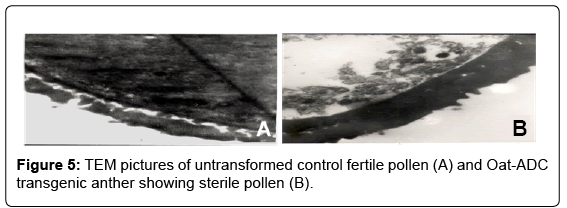
Figure 5: TEM pictures of untransformed control fertile pollen (A) and Oat-ADC transgenic anther showing sterile pollen (B).
PAs play pivotal role in regulation of fertility. They, especially their conjugates with hydroxycinnamic acid are abundant in flowers compared to leaves or other vegetative tissues [41]. The Phenylpropanoid PA conjugates like acylated Spd, N1, N5, N10-trihydroxyferuloyl Spd and N1, N5-dihydroxyferuloyl-N10-sinapoyl Spd are major constituents of the outermost protecting layer of pollen called pollen-kit in A. thaliana [42,43]. Also, PAs are indispensable for pollen germination and tube growth [28,29,44]. They facilitate pollen tube growth by inducing the release of cytosolic Ca2+ ion concentration via one of their catabolic byproducts, hydrogen peroxide (H2O)H2O2 activates the opening of Ca2+ permeable channels in pollen [45].
PAs are maintained in a relatively narrow concentration range in the cell. They cause cellular toxicity beyond a threshold level [46,47] whereas, their deficit negatively affects growth and development of plants [3]. Cell maintains the homeostatic pool of intracellular PAs by regulating the cyclic process of their metabolism, conjugation, binding and transport [48]. PAs generate reactive oxygen species (ROS) particularly H2O2 during their catabolism [48]. ROS act as signaling molecules in various stress conditions and developmental processes [49]. In plants, various stress and developmental phenomenon are associated with the perturbed ROS levels. During developmental process like root development, root hair growth, stress, ROS increases in a spatial and temporal manner [50-52]. They are also known to regulate program cell death (PCD) [53,54] and therefore, a balanced titre of intracellular ROS is crucial for proper cellular function. This balance is maintained by interplay between its production and scavenging mechanism [49,55,56]. Recently Hu et al. reported that timely production of ROS, particularly superoxide anion is associated with the tapetal PCD initiation during anther development in rice [57]. They found that MADS gene product (involved in ROS homeostasis) modulated ROS homeostasis is crucial for late anther development. Moreover, disturbance in ROS homeostasis results in aborted anther wall and disrupted pollen development due to oxidative stress [57].
In the present study, over-expression of ADC gene in tapetal tissue of tomato resulted in increased levels of PAs in transgenic anthers, and this in turn interfered with the pollen and tapetum development with the loss of pollen viability. Transgenics displayed significant increase in bound fraction of Put and Spd, suggesting the conversion of free form of PAs to bound fraction. This could be due to the much regulated homeostasis of the PAs in cellular machinery to avoid the toxicity [48]. Although, how PA accumulation due to the over-expression ADC gene interferes with tapetum and pollen development remains unknown, we speculate that accumulation of PAs beyond a threshold in tapetal tissue might have caused toxicity which must have caused disturbance in tapetum and pollen development. However, this warrants further investigation.
Similar to our observations, Rastogi and Sawhney, while working on a male sterile mutant sl-2/sl-2 observed that increase in PA levels in floral organ causes abnormality in stamen development and decline of the same restores the normal stamen development [32]. They also reported that the exogenous PAs interferes with the in vitro growth of stamen and gynoecium [33]. Interestingly, the constitutive overexpression of ADC2 in Arabidopsis resulted in host dwarfism and delayed induction of flowering [35]. In case of kiwifruits, which is morphologically hermaphrodite but functionally unisex, the high level of free- and conjugated PAs was found to be the reason for the pollen degeneration as well as male sterility [34].
In conclusion, the present results provide an efficient approach to generate male sterile lines in an important crop plant, tomato, thus paving the way to generate hybrid vigor lines and containment of transgene flow via pollen dispersal into the environment.