Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Review Article - (2013) Volume 4, Issue 1
Among the important challenges to shrimp aquaculture worldwide are diseases caused by viruses, in particular by White Spot Syndrome Virus (WSSV) whose genome of 305 kb has been recently sequenced. WSSV, also dubbed White Spot Bacilliform Virus (WSBV), is a major shrimp pathogen with a high mortality rate and a wide host range. The sequencing and characterization of different strains of WSSV has begun to reveal aspects of its biology, virulence and pathogenesis. Knowledge on these aspects is critical for developing effective control methods. The socioeconomic impacts of the diseases caused by the WSSV have been catastrophic in some shrimp producing countries of Asia and the Americas. Thus, these diseases were listed by the World Animal Health Organization (or Office International des Epizootics, OIE) as posing a significant threat to cultured and wild crustaceans as a consequence of international trade or movement of infected organisms. The aim of this review is to present a state-of-the-art knowledge in different aspects of WSSV like morphogenesis, pathogenesis, transmission risks, detection, bio-inoculation studies, and international rules and standards.
Keywords: WSSV, transmission risks, morphogenesis, pathogenesis, shrimp aquaculture, bio-inoculation studies.
The global shrimp production showed a declining trend from 1994 to 1997. The production figured 733,000 metric tons (MT) in 1994 and declined to 712,000 MT in 1995 and 693,000 MT in 1997 [1]. This declining trend was mainly attributed to diseases caused by WSSV. Shrimp farming in India showed an increasing growth from 35,000 MT in 1991-92 to 62,000 MT in 1993-94. However, the production was found to decline from 82,850 MT in 1994-95 to 66,875 MT in 1997-98 [2] and in Asian countries, the value was found declined from 36,104,518 (US$ '000) in 1997 to 35,839,499 (US$ '000) in 1998 [3]. The natural marine ecology is also threatened by WSSV as the virus has a wide host range, including salt and brackish water penaeids, crabs, spiny lobsters, freshwater shrimp and crayfish [4]. The outbreak of viral diseases during 1991 caused a major set back in the shrimp industry. WSSV was first described in China and Taiwan in 1991 in Penaeus japonicus. WSSV spread rapidly to shrimp farming areas throughout Asia associated with a widespread pandemic by 1994 [5]. The WSSV causes serious economic losses because of 100% mortality leading to total crop losses within 3-10 days under some farming conditions [6].
The spread of exotic viruses in cultured shrimps can be attributed to several avenues of introduction. History has documented the introduction of animal diseases to previously uninfected areas by transportation of infected stocks, by birds acting as vectors of diseases, and by import and processing of frozen food products [7,8]. Several developed countries like USA, Japan, EU and Australia import frozen shrimps from various Asian countries particularly from China, India, Thailand, Taiwan, Philippines and Indonesia, and Latin American countries like Brazil, Mexico and Ecuador [7]. Imports are typically repackaged at processing plants that are located in the importing countries.
Risk pathways for transmission of exotic shrimp viruses from imported fresh/frozen products intended for human consumption to wild and farmed shrimp have been proposed to be shrimp reprocessing waste, packaged shrimp diverted for recreational fishing bait, and packaged shrimp diverted for shrimp feed in shrimp hatcheries or in shrimp aquaculture ponds. Risks of shrimp virus introduction during trading of live shrimp for culture have also been described, but other potentially important sources of shrimp viruses are ship ballast water as well as frozen seafood products [9].
Fresh/frozen shrimp products imported into Australia for human consumption were subjected to mandatory testing using PCR technology for three major shrimp viruses - WSSV, Yellow Head Virus (YHV) and Infectious Hypodermal and Haematopoietic Necrosis Virus (IHHNV) at the level of 5% prevalence with 95% confidence from October 2007. Batches that tested positive are destroyed or cooked in an approved facility [10,11]. The requirement for IHHNV testing was dropped in September 2008, but testing requirements for YHV and WSSV remain unchanged. Export of processed shrimp products from India to other countries involves frequent detention of our commodity that causes great economic losses to cultured shrimp farms.
Viral disease diagnostics on imported fresh and frozen foods have been mainly implemented for those produced in Latin America or Asia [7,12-14]. Risks of shrimp virus introduction during trading of live shrimp for culture have been described. Other potentially important sources of shrimp viruses suggested by several authors include ship ballast water and frozen seafood products [8,12-14]. The WSSV encountered in shrimps is listed as a non-exotic disease in EC directive 2006/88 [15].
Different Nomenclatures for WSSV
The WSSV name is derived from the clinical sign that has been reported in some susceptible penaeid shrimp hosts, for example the presence of white spots associated with calcium deposition on the inner surface of cuticle [16]. The WSSV virus received different names during the first years of its appearance, due to its gross and clinical signs. Hypodermal and haematopoietic necrosis baculovirus (HHNBV), chinese baculovirus (CBV), systemic ectodermal and mesodermal baculovirus (SEMBV), penaeid rodshaped DNA virus (PRDV) or rod-shaped nuclear virus of Penaeus japonicus (RV-PJ), and white spot disease (WSD) are some of the terms used by shrimp disease investigation teams [5,15,17-19].
Taxonomy of WSSV
The WSSV was earlier known as white spot baculovirus (WSBV). However the ‘baculo’ prefix was later dropped in accordance with the sixth report of the International Committee on Taxonomy of Viruses (ICTV) that regrouped non-occluded baculovirus into unassigned invertebrate viruses. Recently, the phylogenetic studies on large (RR1) and small (RR2) subunits of ribonucleotide reductase genes showed that WSSV and baculovirus are not closely related. WSSV was assigned by ICTV as the only member of the genus Whispovirus in the family Nimaviridae [15,20,21]. Today, although various geographical isolates with genotypic variability have been identified, they all are classified as a single species within the genus Whispovirus [22].
Target Organs and Mechanism of Spread
The major targets for WSSV infection are tissues of ectodermal and mesodermal origin including those of the epidermis, gills, foregut, hindgut, antennal gland, lymphoid organ, gonads, haematopoietic cells and cells associated with the nervous system [23,24]. Epithelial cells of organs of endodermal origin such as the hepatopancreas, anterior and posterior midgut caeca and midgut trunk are refractory to WSSV infection [25]. In the late stages of infection, the epithelia of the stomach, gills and integument may become severely damaged. This may cause multiple organ dysfunctions and probably lead to death.
The portals of WSSV entry into the shrimp have not yet been clearly identified. Studies have shown variations in sites with respect to the entry. However, the primary sites of WSSV replication in early juvenile Penaeus monodon (per os challenge with WSSV infected tissues) were found out as subcuticular epithelial cells of the stomach and cells in the gills, in the integument and in connective tissue of the hepatopancreas determined by in situ hybridization [26]. Another study conducted on Marsupenaeus japonicus indicated that epithelial cells in the mid gut trunk may be a transient site for WSSV replication which would allow the virus to cross the underlying basal lamina [27]. However, WSSV challenge by immersion showed that haemocytes migrating to gills and midgut were WSSV negative at late stages of infection [28]. The sites of WSSV replication on per os challenge of shrimp using viral particles were shown to be cells in the antennal glands in addition to the epithelial cells in the foregut and cells in the gills (with a high dose of infection) [16].
Morphology of WSSV
WSSV virions are enveloped, rod-shaped nucleocapsids with a bacilliform to ovoid or ellipsoid shape [22]. In some virions, the most characteristic feature is the tail-like projections extending from one end [29,30]. The viral envelope, having a thickness of 6-7 nm, is a lipidic, trilaminar membranous structure with two electron transparent layers divided by an electron opaque layer. The nucleocapsid is located inside the envelope and has a striated appearance and a size of about 300 x 70 nm with a 6 nm thick external wall [23]. The size of the nucleocapsid is found to vary between isolates [16]. The striations are probably the result of stacked ring like structures consisting of rows of globular subunits of about 10 nm in diameter. These subunits are arranged in 14-15 vertical striations located every 22 nm along the long axis, giving it a cross-hatched appearance. When released from the envelope, the nucleocapsid increases in length indicating that it is tightly packed with the virion. The stages of WSSV morphogenesis are found to be directly related to the development of cellular lesions in the infected shrimps. There are 10 stages in the morphogenesis of WSSV [16].
1. Infectious WSSV particles.
2. An infectious WSSV virion attaches to a susceptible cell using envelope proteins with a cell attachment motif.
3. WSSV enters the cell.
4. The envelope of the WSSV virion probably fuses with the endosome and the naked nucleocapsid is transported to the nucleus, in a similar way as in baculoviruses.
5. The naked WSSV nucleocapsid attaches to the nuclear membrane and the WSSV genome is released into the nucleus.
6. The WSSV genome replication starts in the host cytoplasm. The host mitochondria starts degenerating.
7. In the nucleus, the early virogenic stroma appears to be composed of loose granular material. Cellular chromatin accumulates near the nuclear membrane and the rough endoplasmic reticulum (RER) becomes enlarged and active.
8. The marginated chromatin is transformed in a dense ring zone (shaded area). The virogenic stroma is less dense and starts forming vesicles that will form the viral envelope. The vesicles are probably formed with membranous material found in the ring zone as in baculoviruses. A viral nucleosome is also observed as a filamentous structure in the virogenic stroma. This structure contains proteins that will form the nucleocapsid.
9. New WSSV particles are assembled in the nucleus within an electron-dense inclusion. The empty envelopes are filled with a nucleocapsid. In the cytoplasm, organelles become disintegrated and the cellular and nuclear membranes are disrupted.
10. WSSV virions are completely formed and released from the disrupted cell to begin the cycle in other susceptible cells.
The WSSV infects a wide range of aquatic crustaceans ranging from marine, brackish water and freshwater captured and cultured crustaceans and other arthropods, such as penaeid shrimps (Penaeus aztecus, P. chinensis, P. duorarum, P. indicus, P. japonicas, P. merguiensis, P. monodon, P. penicillatus, P. schmittii, P. semisulcatus, P. setiferus, P. stylirostris, P. vannamei); other shrimps (Acetes sp., Exopalaemon orientalis, Macrobrachium idella, M. lamerrae, M. rosenbergii, Metapenaeus dobsoni, M. ensis, Palaemon adspersus, P. sirrifer, P. styliferus. Parapenaeopsis stylifera, Scyllarus arctus, Solenocera indica, Squilla mantis, Trachypenaeus curvirostris); crabs (Calappa lophos, Portunus sanguinolentus, Charybdis sp., Helice tridens, Paratelphusa hydrodomous, P. pulvinata); wild lobster (Panulirus spp.); copepods; pupae of Ephydridian insects; crayfish (Orconectes punctimanus and Procambrus clarkii); pest crab (Sesama pictum); mud crab (Scylla serrata); and many other marine crustaceans [23,32,33].
The first publication on possible disease transmission risks from shrimp commodities was the seminal study of Lightner et al. [35]. This review attempted to examine possible routes of disease transmission and the authors concluded that mechanisms for transfer of exotic viruses such as WSSV, YHV, IHHNV and TSV in imported, frozen commodity shrimp from importer locations and distribution channels to shrimp aquaculture facilities or to wild crustaceans and shrimp may be common in the USA. Those mechanisms may include reprocessing of imported shrimp at processing plants located in fishing ports and release of untreated liquid and solid wastes from these plants into coastal waters, the disposal of solid wastes (heads, shells, etc.) in land-fills where seagulls and other shrimp-eating birds consume virus-infected tissues and then transport the virus to (and contaminate) shrimp farms or coastal estuaries through their faeces, the use of imported shrimp as bait by sports fishermen in coastal waters and the use of imported shrimp as ‘fresh food’ for the maintenance of other aquatic species. It is also known that shrimp eat appendages of dead shrimp and little of the remaining carcass, and that the carcass has the lowest transmission coefficient [35]. Subsequent publications reported experimental work on some aspects of the transmission routes proposed in this initial review. These major publications revealed that shrimp viruses may remain viable in frozen shrimp. The successful transmission of WSSV and YHV to experimental shrimp by the injection of viral extracts from some of the commodity shrimp samples that were having gross clinical signs of infection as well as highly positive for viral infection by PCR methods [7,8,14,36,37]. Briefly, the viral inoculation method consisted of tissue homogenization followed by differential centrifugation ending with ultracentrifugation of the supernatant solution of the infected shrimp homogenates. This was followed by re-suspension of the ultracentrifuge pellet into a small volume of buffer for injection into test shrimp [8,9,22,38]. Hill [39] reported the transmission of WTD through imports either in live or in dead conditions and observed that probably the factors like translocation of infected brood shrimp and post-larvae, highly virulent agent with a wide range of potential hosts and vectors, use of unprocessed shrimp meal in feed, vertical transmission and latency of the virus, poor understanding of the disease at the beginning by the researchers and farmers and the absence of adequate detection methods (i.e. PCR) at the early stage, may influence the transmission. The virus can also be transmitted through cold stored and cooked shrimp products [40,41].
The incidence of WSSV in cultured shrimp products was found to be high with 66% of showing positive in nested PCR reaction (Table 1). Nested PCR yielded good results than the single step PCR in which, the incidence was to a maximum of 29%. The incidence rate of WSSV in small sized shrimp product samples (71%) was more than in the big sized shrimp products [24]. Sritunyalucksana et al. [11] observed that marketed shrimps of sizes less than 15 g originated from shrimp ponds if they have undergone emergency harvest after the advent of high mortality due to known viral diseases including WSSV. Because of this reason only, small sized shrimps were selected by Durand et al. [7] from market samples for their studies on transmission of WSSV into native shrimps from imported frozen commodity shrimps. Jory and Dixon [42] also observed that small count size is an indication that the shrimps were from an emergency harvest, which is a common practice in Asian shrimp farms to save a crop once WSSV disease has been diagnosed.
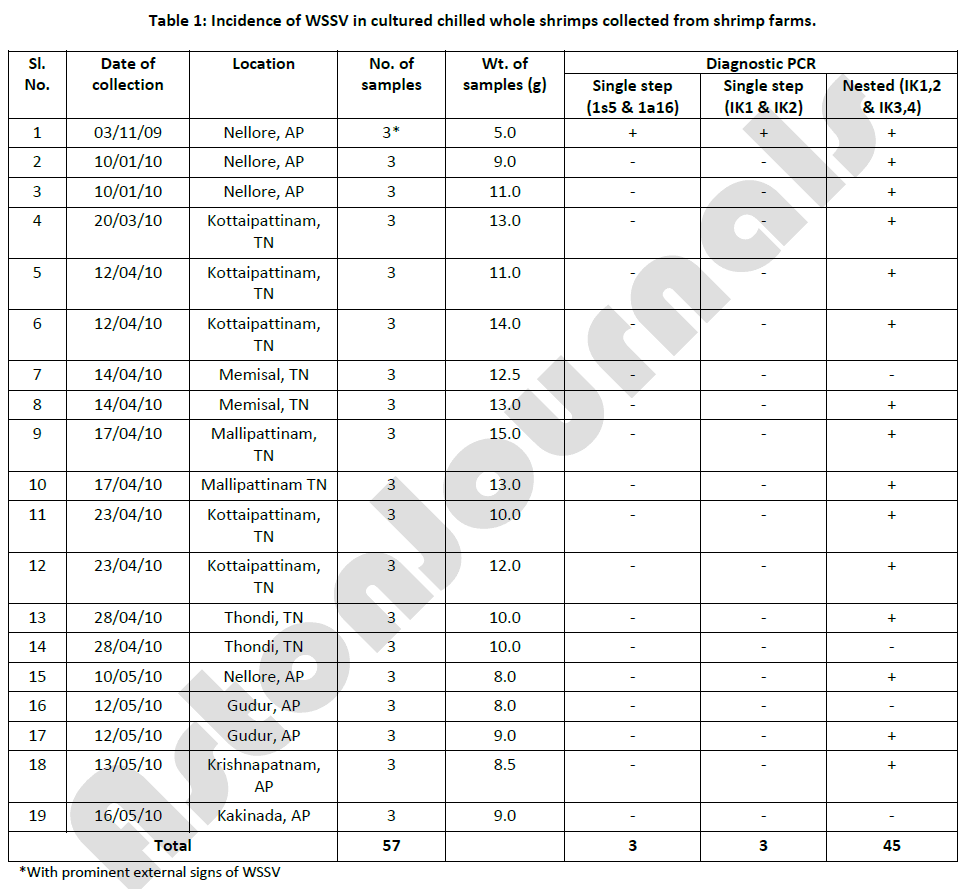
Table 1: Incidence of WSSV in cultured chilled whole shrimps collected from shrimp farms.
Some of our laboratory experiments showed the incidence of WSSV in the cultured shrimp products obtained from seafood processing plants was 66% (Table 2), while it was 79% in cultured chilled whole shrimps procured from shrimp farms [24]. Nunan et al. [36] reported a prevalence of WSSV in 50% of frozen shrimps sold in grocery stores in Texas, USA. Durand et al. [7] reported that the incidence of WSSV in various retail outlets in Tucson, Arizona, USA was 80%. It has also been found that WSSV was present in frozen shrimp (Penaeus monodon) imported into Australia from Asian countries [8]. Ostrowski [43] found that 73% of the shrimp samples surveyed from grocery-store in Hawaii by PCR were positive for WSSV. The present results are in line with the earlier findings. But, Reville et al. [12] have detected WSSV in only 4.7% of shrimp products in supermarkets of Massachusetts, USA. In Mexico, Reyes-Lopez et al. [14] reported the incidence of WSSV in frozen shrimp products as 32%. In India, Reddy et al. [24] found WSSV in 73% of the frozen shrimp products surveyed from different processing plants. The variation in prevalence of WSSV in shrimp products observed by different workers might be due to the differences in the pattern of outbreaks of WSSV in shrimp farms at different point of time of sampling.
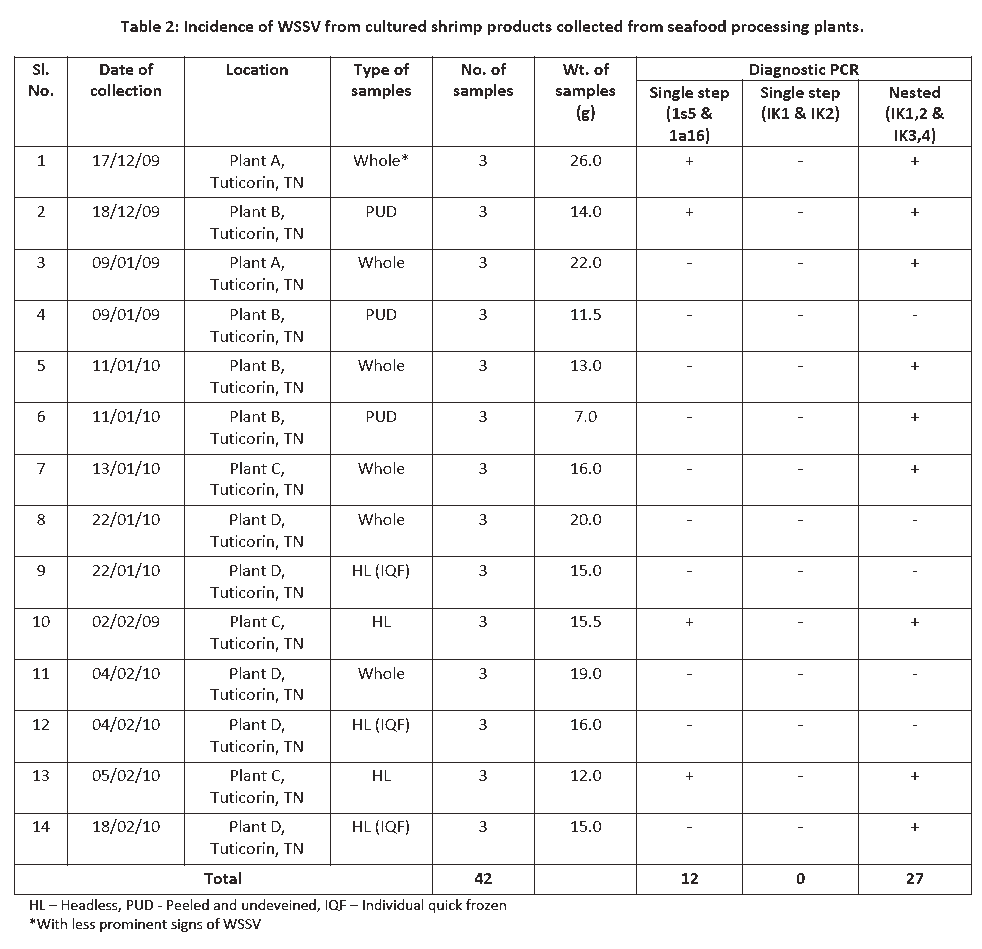
Table 2: Incidence of WSSV from cultured shrimp products collected from seafood processing plants.
In our laboratory, bio-inoculation studies were conducted on the healthy WSSV-free shrimps using viral inoculum from infected shrimp product tissues showed 100% mortality within 28-45 h of P.I. Negative control group were injected with inoculum from negative shrimp extract. The progress of WSSV infection in the bio-inoculated shrimps showed that WSSV first affected the pleopods followed by uropods. The injected shrimps showed gross clinical signs of WSSV infection. The WSSV infected animals on bio-inoculation exhibited pinkish red discolouration on the uropods and pleopods [24]. Sudha et al. [44] also reported that the shrimps with reddish discolouration without white spots were a preacute form of the disease caused by WSSV. The disease process may be completed in a short time period because of high viral load and small pore size, and therefore, the principal clinical sign of white spot appearance normally associated with WSSV syndrome may not develop at all. The successful WSSV infection on infectivity bioassays in all of the experimental animals caused rapid reduction in feed intake and lethargy [45], which was also observed in the present study. Of the 5 animals injected with the viral inoculum from frozen cold stored shrimps, the first mortality was observed at 28 h P.I. No clinical signs of disease or mortality were observed among negative control shrimps during the experimental period. Lightner [46] also reported the appearance of reddish discoloration or small white spots in the WSSV infected P. monodon. Durand et al. [7] observed 100% mortality in the indicator shrimp [specific pathogen free (SPF) P. vannamei] by injection of WSSV inoculum from frozen shrimp products. All the 5 animals in the present study died within 45 h after P.I., which showed that the viability of WSSV in shrimps was not affected by freezing cold storage.
Nunan et al. [36] reported that the bioassay using 15 P. stylirostris indicator shrimps produced 100% mortality within 8 day P.I. of WSSV inoculum from frozen shrimp products. The series of experiments conducted by Nunan et al. [36] also showed that frozen product imported from Asia contains the viral pathogens, WSSV and YHV and through the use of bioassay, they demonstrated that the viruses in the frozen imported product were infectious. Durand et al. [7] observed that the challenge bioassay of WSSV by shrimp injection produced the first mortality in indicator shrimp on day 3, but, the entire indicator shrimps had died by day 4. Hasson et al. [13] observed the clinical signs of disease including lethargy, anorexia and chromatophore expansion resulting in dark body coloration and reddening of both the uropods and antennae, when SPF Litopenaeus vannamei juveniles were injected with WSSV PCR-positive Parapenaeopsis spp. tissue homogenates resulting in 100% mortality in the experimental shrimps within 48 to 72 h of P.I.
The WSSV takes some time to express itself but once after its expression, infected animals die within 3-8 days resulting in high mortality [45]. The WSSV infected shrimp in the field is found to gather near the pond edge and display clinical signs in 1 or 2 days before occurrence of any mortality. Cumulative mortality may reach 100% within 10 days after the onset of disease [47,48]. In grow-out ponds, juvenile shrimp of all age and sizes are susceptible to the disease but massive mortality occurs 1 or 2 months after stocking [49]. The most obvious symptom of WSSV infection is the presence of circular white spots or patches of 0.5-3.0 mm in diameter most prominent in the cuticle of cephalothorax and tail part [5,50]. Expression of WSSV infection furnished with some characters as white to reddish-brown / reddish / pinkish / to discoloration [5,45] over the head and carapace, low appetite, gather near the embarkment etc., which are not distinct from some other viral infection of bacterial white spot syndrome [51].
Even though the exact mechanism of white spot formation is not known, it is possible that the WSSV infection may induce the dysfunction of the integument resulting in the accumulation of calcium salts within the cuticle and giving rise to white spots [52]. Other symptoms of the disease include a reddish discolouration of body and appendages because of the expansion of chromatophores [53,54], less intake of feed [55,56], reduced preening and low response to stimulus [19,57], loose cuticle [58], swelling of branchiostegites because of accumulation of fluid [25,59] and thinning and delayed clotting of haemolymph [60]. Histologically, infection is characterized by eosinophilic to progressively more basophilic inclusion bodies in the hypertrophied nuclei of infected cells [58]. Infected nuclei become progressively more basophilic and enlarged [59,61-63]. In the late stages of infection, karyorrhexis and cellular disintegration may occur, leading to the formation of necrotic areas characterized by vacuolization [47,49].
The WSSV is viable for at least 30 days at 300C in seawater under laboratory conditions [64] and is viable in ponds for atleast 3-4 days [22,65]. The WSSV has been found to be inactivated in <120 min. at 500C and <1 min. at 600C [65]. According to Maeda et al. [66], sodium hypochlorite inactivated PRDV at 1ppm for 30 min. and at 5ppm for 10 min. SEMBV at 10 ppm for 30 min. Povidone-iodine inactivated these viruses at a concentration of 10 ppm for 30 min. A high concentration of NaCl (12.5%) inactivated PRDV in 24 h at 250C and 15% NaCl inactivated SEMBV in 24 h at 280C. PRDV was inactivated by heating at 500C for 20 min, by drying at 300C for 1h and by using ethyl ether at 40C for 18 h. The PRDV in the seawater maintained its infectivity for at least 120 days at 40C and for more than 60 days but less than 120 days when kept at 250C. According to Reddy et al. [40], various processing treatments such as icing, freezing, cooking, cooking followed by slow freezing, cooking followed by quick freezing, canning, and cold storage were employed to destruct the WSSV DNA. But, among all these processing treatments, cooking followed by quick freezing found to be very effective in destruction of WSSV DNA.
The WSSV genome is a circular dsDNA molecule and is one of the largest animal virus genomes that has been entirely sequenced [67-69]. The genome size varies according to the viral isolate. Three complete WSSV sequences (accession numbers AF369029, AF332093, AF440570) identified and the sizes of the genomes were found to be 292,967 bp, 305,107 bp, 307,287 bp for the Thailand, China and Taiwan isolates, respectively [69,70]. The nucleotide sequence analysis revealed that the WSSV genome encodes approximately 185 open reading frames (ORFs) of 50 amino acids or more. With completion of the WSSV genomic DNA sequence, research has now been focused on the functional analysis of the gene products, especially, on the functions of the viral envelope proteins. A few structural genes and DNA metabolism related genes have also been identified and characterized [71-78].
A virion is a complex assembly of macromolecules exquisitely suited for the protection and delivery of viral genomes. Its structural proteins are found to be highly important, since these proteins are the first molecules to interact with the host, and they therefore play critical roles in cell targeting as well as in the triggering of host defences [79]. The characterization of structural proteins and their genomic sequence is of major importance to determine the taxonomic position of viruses. Furthermore, the structure and interaction of the WSSV virion proteins may explain the unique morphological features of the virus. Diagnostic tests could therefore be designed based on one or more of these structural proteins [68] Several WSSV proteins have been characterized. Some non-structural proteins are found to be involved in transcriptional regulation (VP9) [80], virus proliferation (WSV 021) and / or regulation of DNA replication (WSV 477). Of the total proteins, 21 have been found in the envelope, 10 in the nucleocapsid and 5 in the tegument (putative structure located between the envelope and nucleocapsid).
The non-structural proteins identified in the WSSV genome are found to be required for the replication of the viral genome, production of the virus particle, and inhibition of the cell functions. These proteins therefore prove as potential candidates for drug design and the development of vaccine. VP9, a full length protein of WSSV, encoded by ORF115, was identified for the first time in the infected P. monodon shrimp tissues, gills and stomach as a novel, non-structural protein by Western blotting, mass spectroscopy and immunoelectron microscopy. Although, the exact cellular function of VP9 is not clear, studies have identified VP9 as an abundant protein in WSSV infected host tissue [80]. X-ray and NMR based structural studies revealed that VP9 possesses a DNA recognition fold with Zn ions specific binding sites [81,82]. All these results indicate that VP9 could act as a transcriptional regulator of WSSV.
The envelope proteins are particularly important for enveloped viruses, because these proteins often play vital roles in viral entry, assembly and budding [83]. The envelope proteins VP31, VP110 and VP281 show the presence of a cell attachment motif which has a role in viral entry [79,84-86]. The tegument proteins VP36A and the nucleocapsid proteins VP664 [79,87] and VP136 A [79,88,89], are also characterized by the presence of a cell attachment motif. Other proteins such as VP28, VP39B, VP31A, VP41B, VP51A, VP51B, VP68, VP124, VP150, VP187, VP281, VP292 and a collagen like protein [85] have been located in the envelope [68, 70, 84, 88], whereas the proteins VP35 [71], VP466 [89], VP15, VP51 and VP76 [90] are found to be located in the nucleocapsid and may have different putative functions [68].
Among these proteins, VP466 is found to have an important role in virus penetration [91]. P466 is designated by the ORF151, is one of the latency associated genes in WSSV, and is found to be a glutathione S- transferase (GST) fusion protein [88]. VP76 is designated by the ORF112, contains the conserved domain of Class I cytokine receptors [68, 69] and is involved in WSSV infection. ORF112 is 2025 nt long and encodes a protein of 675 aa with a theoretical molecular mass of 76 kDa [92]. Phylogenetic analysis has shown that ORF112 contains the conserved motif sequence of a Class I cytokine receptor and has several glycosylation sites and a signal peptide, but no transmembrane domain. This implies that WSSV has evolved mechanisms to evade the host defence system (cytokines) in favour of virus infection and replication.
Various diagnostic procedures have been developed for detection of WSSV. These include histopathological techniques [14,57], in situ hybridization [53], immunological methods such as Nitrocellose-enzyme immunoblot [93] and Western blot techniques [19,55] and more recently highly simple, sensitive and reliable technique such as Polymerase Chain Reaction based methods [59,94-96]. The level of sensitivity with the use of different DNA based techniques seems to vary. This is observed in a recent study of identification of WSSV latency-related genes by use of microarray [97]. However, from the practical point of view, among the various diagnostic techniques, PCR provides a high degree of sensitivity and specificity in detection of WSSV [95]. PCR has been used recently to detect WSSV in a very specific and sensitive manner. Nested or two step PCR has the advantage of increasing the level of sensitivity over single step PCR. Nested PCR consists of the reamplification of the PCR product obtained on a single step PCR reaction by using an aliquot of this first reaction product as a template in a second round of amplification. When a shrimp shows clinical signs of WSSV, it is easily detected by single step PCR. However, at low viral loads, WSSV is latent without causing disease symptoms in the shrimps, and can be detected by nested PCR [59,98]. Sensitivity comparison of several PCR methods commonly employed in Thailand such as Taq Man real-time PCR, one-step PCR, single-tube nested PCR, two tube nested PCR and triple blind ring test [96]. Several authors studied the use of reverse transcription PCR for the detection of WSSV [19,69,73,99]. Durand and Lightner [29] developed a quantitative real time PCR method for the detection and quantification of WSSV in samples.
A number of primers have been designed for PCR detection of WSSV by several research groups. Lo et al. [58] designed three sets of primers based on sequence of cloned WSSV Sal I 1461 bp DNA fragment. The primers 146F1/146R1 gave an amplicon of 1447 bp while primers 146F2/146R2 and 146F4 / 146R3 yielded amplicons of 941 and 775 bp, respectively. Another set of primers designed by Wongteerasupaya et al. [57] were based on the sequence of a specific probe yielding a 294 base pair fragment specific for WSSV DNA. Takahashi et al. [63] developed a primer pair which amplified a 643 bp fragment of RV-PJ of Japan and designated the primers as PJ1 and PJ2. Kim et al. [94] designed two primers named IF and IR based on the sequence data of WSSV DNA which gave an amplicon of 365 bp. Nunan et al. [36] designed two sets of primers based on sequence of WSSV genome. The first set (N/L primers) designated as 6581 and 7632 amplifies a 750 bp sequence of WSSV DNA while the second set of primers designated as 9987 and 9988 amplifies a 500 bp internal sequence to the first set of primers. Kasornchandra et al. [49] used two oligonucleotide primers designated 102 F1 and 102 R1 which amplified a 520 bp fragment of white spot virus genomic DNA. Magbanua et al. [100] used a set of DNA oligonucleotide primers originally designed by Tapay et al. [101] which gives an amplicon of 217 bp. Hossain et al. [102] and Pradeep et al. [103] used two sets of primers, designated IK1– IK2 and IK3– IK4 that were based on sequence from Gene Bank Accession No. U50923 giving an amplicon of 486 and 310 bp, respectively. A method for the detection of degraded WSSV DNA was developed by the Australian Animal Health Industries of CSIRO based on the primers 1s5 and 1a16, which gave an amplicon of any one of the following products size of 480, 420, 385 280 and 198 bp [104].
Shrimp farming is limited in the European Union (EU) in comparison to the world production estimates [105], but interest is increasing, particularly in the use of intensive re-circulation rearing systems and some pilot operations have started [39]. Concerns have been expressed about the potential risk that WSSV poses to both cultivated and wild populations of European native crustaceans because of its unusually large host range. Corbel et al. [45] observed mortality of European crustaceans due to WSSV infection. McColl et al. [8] detected the presence of WSSV and YHV in frozen shrimps imported from Australia. The authors warn on the high potential risk of spreading of WSSV in EU waters and emphasised the justification for a rigorous monitoring of imported shrimp, including frozen consignments, to prevent introduction of the virus and subsequent losses of cultivated and natural populations of crustaceans. East et al. [104] examined the populations of Australian crustaceans for evidence of WSSV and found no mortalities or clinical signs of disease or evidence of WSSV during the survey.
It can be inferred that WSSV DNA is not destroyed by icing, freezing, cooking, and cooking followed by slow freezing, canning, and cold storage processes. The nested PCR is the most reliable technique to detect WSSV DNA from shrimp products. It is clear that WSSV DNA could be destroyed by cooking followed by quick freezing process. Hence, the transmission risk of WSSV from infected shrimp products to the native aquaculture systems could be reduced by coupling the processes of cooking with quick freezing and thereby reducing the risk of economic losses to the aquaculture industries throughout the world.
Authors wish to thank the Dean, Fisheries College and Research Institute, Tuticorin, India for providing all facilities and support to carry out this study.