Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Review Article - (2013) Volume 2, Issue 3
A recombinant single chain fragment variable (scFv) antibody was constructed from hybridoma cell lines expressing anti-solamargine (As) monoclonal antibody (MAb). The characteristics of the As-scFv protein, expressed in both recombinant Escherichia coli and hairy root, were almost same with those of the original MAb. Up to 220 ng recombinant As-scFv was produced per milligram of soluble protein in transgenic hairy root cultures of S. khasianum. The solasodine glycosides concentration was 2.3 fold higher in the transgenic, than in the wild-type hairy root, reflecting from the soluble As-scFv level. The recombinant S. khasianum plant regenerated from recombinant hairy roots contained 2.3 fold concentration of solasodine glycoside, compared to the regenerated plant from wild-type hairy root. These results suggested that the scFv antibody expressed in transgenic plant controlled the antigen level. This methodology is the first success for molecular breeding of secondary metabolites, without understanding of biosynthetic enzyme.
Keywords: Solanum khasianum; Solasodine glycoside; scFv gene; Transgenic plant; Molecular breeding
In our former stage of medicinal plant breeding, we succeeded to clonally propagate Pinellia ternate (Araceae) via protocorm-like body [1], and by liquid phase culturing, which theoretically produced 4×1023 clonal plants from a single tuber in a year. We also investigated the clonal propagation of Aconitum carmichaeli (Ranunculaceae) [2,3], and Panax ginseng [4,5], by embryogenesis. Furthermore, we confirmed the relation between the virus infection and the component concentration, comparing with the virus free plant obtained by a tip tissue culture of Rhemannia species [6] (Scrophulariaceae). In the second stage of medicinal plant breeding, we did make evident that the medicinal plants clonally propagated promoted the homogeneity of bioactive component concentration, compared to the parent plant for aconitinetype alkaloid [7] (Aconitum spp.), atractylon [8] (Atractylodes spp.), and gentiopicroside [9] (Gentiana spp.).
Solanum species belonging to Solanaceae family contain steroidal alkaloid glycoside like solasonine, solamargine and khasianine (Figure 1).
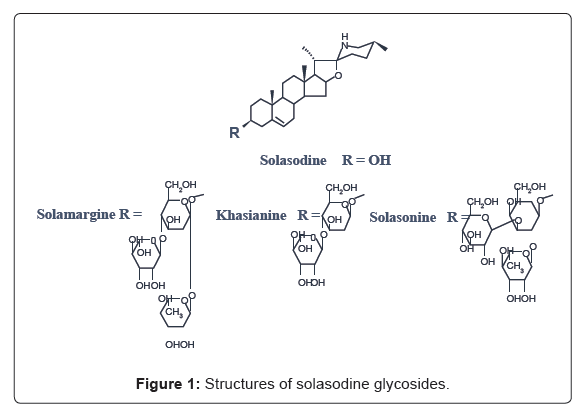
Figure 1: Structures of solasodine glycosides.
Solanum species have been widely used as herb medicines in China and Asian countries. In fact, solasodine glycosides have antiskin carcinoma properties in humans [10,11], and anti-neoplastic activity in mice [12]. Solamargine and solasonine (Figure 1) inhibited an acetyl-cholinesterase [13]. Solamargine is anti-herpes simplex virus type I [14], and a more powerful anti-fungal agent than ketoconazole [15]. Furthermore, since natural resources of adrenocortical and sex hormones, particularly from Dioscorea spp. have been insufficient, steroidal alkaloid glycosides like solamargine and solasonine having solasodine in a molecule as an aglycone (Figure 1) from Solanum spp., have become important as starting resources for the production of steroid hormones. Therefore, we have previously prepared antisolamargine monoclonal antibody (MAb), and confirmed the level of solasodine glycosides by an ELISA [16]. It became evident that the MAb can be used for the survey of all glycosides containing solasodine as an aglycone, because the MAb have wide cross-reactivity against all solasodine glycosides. This property is a big advantage for the survey of resource of solasodine glycosides. Furthermore, we set up a simple staining system named eastern blotting [17], and one step immunoaffinity separation [18].
Recently, it became evident that a single-chain fragment-variable (scFv) antibody has been expressed in plants to protect for viral attack [19-21]. Moreover, scFv can be used to study plant growth, and to interfere with the biological activity of antigens [22-24]. We therefore postulate that the recombinant scFv can target small antigens like solasodine glycosides, which can then lead to the accumulation of antigen-antibody complexes in plant organs.
In order to open a positive molecular breeding for the increase of bioactive component like solasodine glycoside, we constructed a functional anti-solamargine (As)-scFv antibody gene and introduced into S. khasianum plant. In this review, we will discuss whether such the transformation of scFv gene into plants can be used as a new molecular breeding methodology, for the increase of secondary metabolite concentration.
Construction of scFv and Induction into E. coli and Agrobacterium rhizogenes, and Expression of scFv
We constructed scFv using an Escherichia coli expression vector, essentially according to the protocols of the recombinant phage antibody system purification and expression modules [25].
We extracted mRNA from hybridoma cells secreting the As-MAb, SM-BD9 [16], and synthesized first-strand cDNA primed by random hexamers, using murine reverse transcriptase. PCR was performed usingVH and VL primers from the scFv module of the RPAS kit and KOD DNA polymerase. The cDNAs of theVH and VL chain were assembled by seven incubation cycles, using a DNA linker fragment encoding the amino acid sequence (Gly4Ser) 3 supplied with the RPAS kit (Pharmacia). The resulting fragment was ligated into the pCANTAB5E phagemid (Pharmacia), from which high affinity scFv protein was selected. The DNA encoding scFv fused to an E-tag, and an amberstop codon was amplified by PCR from the template pCANTAB5E. The gene was subcloned into the pET-28a(+) expression vector to contain His- and T7-tags at the N-terminus, and an E-tag at the C-terminus.
Figure 2 shows the nucleotide and deduced amino-acid sequences of As-scFv. The hypervariable regions or complementarity determining regions (CDRs) were identified as indicated in figure 2. The nucleotide sequence data appear in the GenBank nucleotide sequence database under the accession number AF332008. The sequence was compared with those compiled in the Kabat antibody sequence database. The VH region of MAb (SM-BD9) belonged to the mouse heavy chain subgroup II(A), whereas the VL region belonged to the mouse kappa light chain V group.

Figure 2: Nucleotide and deduced amino acid sequences of As-scFv antibody. Linker fragment is shown in italics. Nucleotide sequences encoding respective complementarity determining regions (CDRs) are underlined. inhibition of purified scFv protein were determined by direct and competitive ELISA, respectively.
Expression of the recombinant scFv protein in E. coli was induced by incubation with isopropyl-thio-β-d-galactopyranoside (IPTG). After harvesting induced cells, renaturation was initiated by rapid 100- fold dilution of the denatured protein into refolding buffer, consisting of Tris-HCl, arginine and EDTA. Samples were incubated, dialyzed against Tris-HCl, and then purified by metal chelate affinity chromatography. Bound protein was eluted using Tris-HCl, containing imidazole and NaCl. The binding activity and binding inhibition of purified scFv protein were determined by direct and competitive ELISA. Plant expression vectors were transferred with the pRK 2013 helper plasmid into Agrobacterium rhizogenes strain 15834 by triparental mating, and were used to transform S. khasianum using standard procedures [26].
Transgenic hairy roots were cultured on hormone-free halfstrength MS medium containing 60 mg/l kanamycin. Total soluble protein was prepared by homogenizing hairy roots in extraction buffer. Soluble protein was quantified using a Bio-Rad protein assay kit, with bovine serum albumin as standard.
The molecular weight of As-scFv protein produced by transgenic hairy roots was the same as that of the protein expressed in E. coli, having 31 kD as indicated in Figure 3, whereas wild type hairy roots did not produce As-scFv [25].
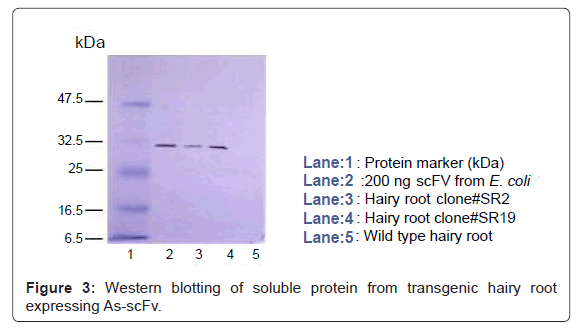
Figure 3: Western blotting of soluble protein from transgenic hairy root expressing As-scFv.
Recombinant protein was reacted with T7-tag antibody, followed by peroxidase-labeled anti-mouse IgG, and then stained with 4-chloro-1- naphthol. Lines 1: Protein marker, 2: 200 ngscFv from E. coli, 3: affinitypurified scFv from hairy root clone SR2, 4: affinity-purified scFv from hairy root clone SR19, 5: soluble protein from wild-type hairy root.
MALDI mass spectrometry confirmed the exact molecular weight of scFv protein produced by hairy root. A molecular peak [M+H]+ appeared at 31,529 m/z, in good agreement with the molecular weight of 31,504 calculated from amino acid sequence [25], as shown in figure 4. From this result, it becomes clear that the constructed gene functioned normally without any mutation.
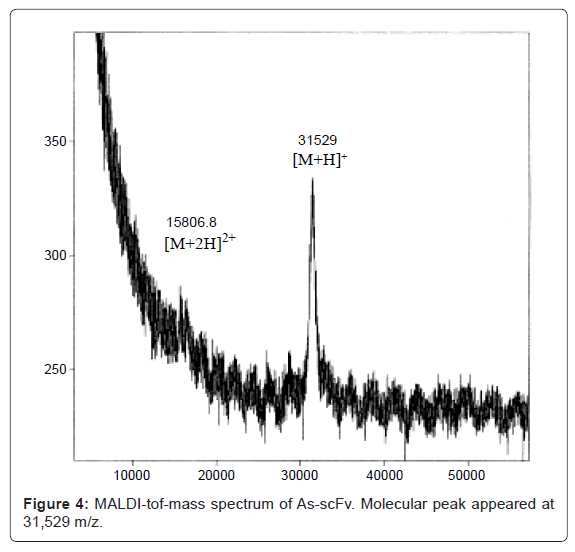
Figure 4: MALDI-tof-mass spectrum of As-scFv. Molecular peak appeared at 31,529 m/z.
In order to confirm the equivalence between the As-scFv produced by hairy root and the original As-MAb, we investigated the cross reactivity of As-scFv and MAb (SM-BD9) [16], against related steroidal alkaloid glycosides. Cross-reactivities of As-scFv fragments against solasodine glycosides having different sugar moieties, such as khasianine (80.7%), solasonine (92.1%), 3-O-β-d-glucopyranosylsolasodine (112.9%), O-α- L -rhamnosyl-(1→2)-3-O-β-d-glucopyranosylsolasodine (98.5%), 3-O-β-d-gala-copyranosylsolasodine (115.4%), O-β-d-glucopyranosyl -(1→3)-3-O-β-d-galactopyranosylsolasodine (118.2%), and isoanquivine (150.4%), were almost same to those for the MAb [18]. The property having wide cross-reactivity against all solasodine glycosides is an advantage, because it is important to accumulate all solasodine glycosides, which are used as the resource of steroidal hormone. Moreover, we analyzed the binding kinetics of Ac-scFV from E. coli, hairy root and original MAb, using competitive ELISA, resulting in almost identical [16]. From these results, it is easily suggested that As-scFv can function for the antibody-antigen reaction in a plant.
The concentration of solasodine glycosides in all clones of As-scFv transgenic hairy roots ranged from 62 to 100 μg/g dry wt., whereas that of the wild type was 43 to 45 μg/g dry wt. It is 2.3-fold higher than that of the wild type hairy root. Of 37 cloned transgenic hairy roots, 7 produced double the amount of As-scFv. The solasodine glycoside concentration increased in proportion to the soluble scFv level [25]. Therefore, the expression level of AsscFv seems to be important for the increase of solasodine glycoside concentration. The solasodine glycoside concentration was two times or more in the transgenic hairy root, indicating that As-scFv protein combines directly to solasodine glycosides, and enhances their biosynthetic pathways in transgenic hairy roots. This evidence suggests that transgenic hairy roots expressing As-scFv produce an antigen-antibody complex that accumulates in plant cells.
In order to confirm the above hypothesis, we regenerated plantlets from hairy roots. Figure 5 shows the regeneration step of S. khasianum plant from transgenic hairy root.
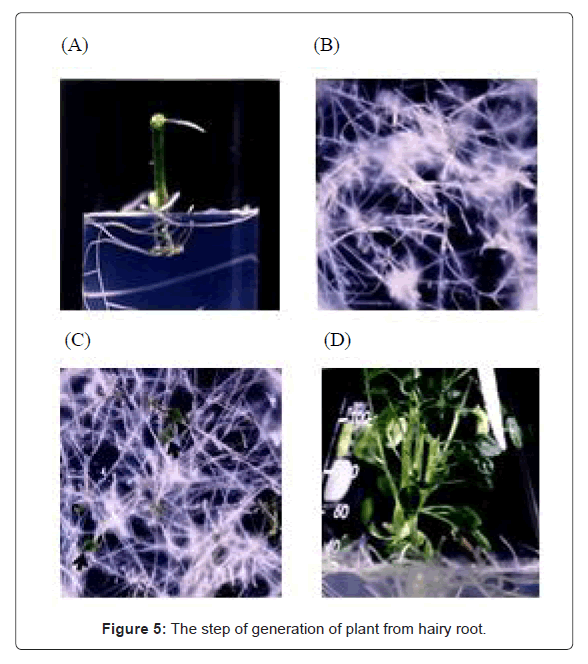
Figure 5: The step of generation of plant from hairy root.
(A) : Infected shoot segment with Agrobacterium rhizogenes.
(B) : Growth of hairy root.
(C) : Appearance of green spot.
(D) : Regenerated S. khasianum plant.
The fruits of transgenic S. khasianum contained 126.95 ± 6.03 μg/ mg dry wt. solasodine glycosides (Figure 6), which was over twice that produced by S. khasianum transformed by wild A.rhizogenes (53.43 ± 3.90 μg/mg dry wt.).

Figure 6: Photograph of regenerated S. khasianum.Leaf and fruit of control (A) and transgenic leaf and fruit (B).
Since the ELISA survey using As-MAb analyzed the total concentration of solasodine glycosides, we determined the qualitative analysis of solasodine glycosides contained in transgenic S. khasianum plant. We indicated the fingerprinting of solasodine glycoside contained in transgenic S. khasianum by Eastern blotting. Since anti-solamargine MAb has a wide cross-reactivity [16], all solasodine glycosides can be stained by Eastern blotting [17]; indicating that three major steroidal alkaloid glycosides; khasianine, solamargine and solasonine, can be clearly detected compared to that of fingerprinting by TLC, although the ratio of individual solasodine glycoside concentration was not clear.
Recombinant antibodies, such as scFv antibodies, have opened biomedical and biotechnological methodology [27], since they possess the same affinity and monovalent binding specificity compared to their parent antibodies [28]. The application of scFv engineering as a diagnostic tool in plant pathology has been demonstrated [29-31]. Other advances in antibody construction owe various means of conferring novel properties upon plants. Resistance to plant viruses can be enhanced using scFv antibody engineering against virus coat protein in plants [19-21], as previously described. Although molecular breeding of secondary metabolites has been owned to biosynthetic enzyme, we discovered that the transformation with the scFv fragment improves the yield of a pharmacologically active compound in this review. We, therefore, postulate that the recombinant scFv can target small antigens like solasodine glycosides, which can then lead to the accumulation of antigen-antibody complexes in plant organs. This system also allows the recovery of both antigen and scFv protein, which can be used for further analytical studies. Our system established in this review may open up a new plant molecular breeding methodology for pharmacologically active glycosides, such as ginsenosides (Panax species), glycyrrhizin (Glycyrrhiza species), saikosaponins (Bupleurum falcutum), sennosides (Rheum species) and crocin (Crocus sativus), that we prepared MAbs against those compounds [32], without understanding of their biosynthetic pathway like a neutralized antibody. Therefore, we named it a missile type molecular breeding. Recently, we succeeded the production of plumbagin in Plumbago zeylanica hairy root, using a scFv against plumbagin as evidence exhibiting 2.2 times higher than those obtained from wild-type plant [33]. The high correlation between the plumbagin-scFv expression level and plumbagin concentration of the recombinant plants suggested that the plumbagin biosynthesis pathway had been modulated by the expression of plumbagin-scFv protein in the hairy roots of Plumbago zeylanica.This result strongly support our postulation discussed in this review. Therefore, we have been further investigating the molecular breeding, by using scFv gene in our ongoing study on plant breeding.