Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2015) Volume 3, Issue 3
Arabidopsis plasma membrane nanodomains have been visualized in vivo using Remorin proteins of the taxonomic group 1b. The physiological role of such domains is to facilitate protein-protein interactions near the plasma membrane and to act as signal transducing platforms, routing information from the extra- to the intracellular side of the cell. Beneath sterols and sphingolipids as lipid components, group 1b Remorin proteins showed a strong tendency to accumulate in nanodomains. In contrast to sterol- and sphingolipid enriched membrane domaincounterparts in animals, individual Arabidopsis nanodomains did not exhibit any kind of lateral movement in the plane of the membrane. Since group 1b Remorins are reported to form microfilaments in vitro, assemblies of Remorin microfilaments with stable structures could have an impact on domains´ lateral stability in the plane of the membrane. To test a possible influence of cell wall components on the mobility of individual Remorin proteins resident in plasma membrane nanodomains FRAP-experiments were performed in Arabidopsis mesophyll tissue and protoplasts, using fluorescently labeled group 1b Remorins. It was revealed that the cell wall has a stabilizing effect on the mobility of individual Remorins resident in nanodomains.
Keywords: Plasma membrane; Nanodomain; Remorin; FRAP; Protoplast
Plant plasma membrane domains have been visualized in vivo using sterol-dependent proteins of the Remorin family [1,2]. The Remorin gene family is unique to plants, found in angiosperms, gymnosperms and bryophytes [3]. To this date there is still minor knowledge concerning specific gene functions of individual family members [4]. The prototype Remorin has been isolated as cDNA from Solanum tuberosum and was initially assumed to be a PM-associated uronide binding protein that gets phosphorylated in the presence of oligogalacturonides [3,5] In vital tomato plants it has been shown that the orthologous protein, named Solanum tuberosum Remorin 1.3 (StRem1.3), is able to physically interact with a movement protein of the potato virus X and thereby to impair viral cell to cell movement [1]. StRem1.3 formed membrane patches when expressed in tobacco, BY-2 cells, and in Arabidopsis. In tobacco, the patch size was revealed to be about 80 nm in diameter using electron microscopy [1]. In Arabidopsis the patch size was measured to be around 100 nm in diameter, employing STED - superresolution microscopy. Recently two Remorin orthologues from Arabidopsis thaliana were isolated; these proteins - namely AtRem1.2 and AtRem1.3 - showed strong sterol dependencies in vitro and formed plasma membrane patches when expressed in plant cells [6] These patches were reminiscent of the patchy pattern of StRem1.3 in tobacco, exhibiting a similar size. Based on the tendency to preferably linger in sterol-rich lipid environments, AtRem1.2, AtRem1.3 and StRem1.3 accumulated in plasma membrane domains when expressed in viable Arabidopsis mesophyll cells where they showed a strong correlation. Therefore, group 1b Remorins were introduced as in vivo marker proteins for plasma membrane nanodomains in plants. Subsequently, it was demonstrated that almost all of the 16 remorin proteins known in Arabidopsis thaliana localize to (different) membrane microdomain platforms [2]. Concerning physiological roles of sterol- and sphingolipid-enriched compartments it was demonstrated in Arabidopsis that regulatory protein-protein interactions occur therein, as shown for the CPK21-mediated activation of the ion channel SLAH3 [7]. It is now established that sterols [8] and sterol-protein interactions play a role in the compartmentalization of plasma membrane proteins in plants and that these interactions play regulatory roles [9]. Findings suggested that Remorin-tagged domains would serve as protein-protein interaction sites in vital Arabidopsis plasma membranes.In animal cells sterol-enriched compartments in the plane of the plasma membrane, involved in mediating signaling processes, have been defined as membrane - or lipid rafts [10,11]. Animal raft domains are characterized by elevated sterol- and sphingolipid levels and further by the ability to include or exclude proteins to variable extents [12]. The majority of raft-affine proteins is involved in signal transduction events, like α-subunits of heterotrimeric G-proteins, glycosylphosphatidyl-inositol(GPI)-anchored proteins, Src-family kinases, particularly palmitoylated transmembrane proteins, and cholesterol-linked and palmitoylated proteins [13]. Beneath differences in their protein- and lipid composition membrane domains in plants differ from their counterpart in animals with respect towards the lateral mobility of these lipid-protein assemblies. In contrast to animal membrane rafts plant nanodomains were detected to be virtually immobile regarding a lateral diffusion in the plane of the membrane, exhibiting high temporal and spatial stability [2]. The immobility of plant membrane domains as a whole or the proteins therein could be explained either by attachment to intracellular structures like the cytoskeleton or microtubules or by fixation to components of the extracellular cell wall. While postulated that the actin cytoskeleton plays an important role in regulating dynamic movement of proteins between different membrane phases, microtubules were not considered important for microdomain formation.
To fathom the nature of this stability, a transgenic Arabidopsis line was created, in which an eGFP-tagged group 1b Remorin has been overexpressed. Real time FRAP-experiments in Arabidopsis mesophyll cells and protoplasts should bring clues concerning the mobility of single eGFP::StRem1.3 fusion proteins in nanodomains. FRAPexperiments might help to identify an involvement of Remorin proteins in the formation and maintenance of a filament-like nanostructure or even a scaffold involved in anchoring individual domains in the plane of the membrane by incurring interactions with stable structures, i.e. components of the cell wall.
Plant transformation
The transformation method based on the floral dipping method, employing an Agrobacterium-mediated gene transfer [14]. Transformed Arabidopsis thaliana Col 0 plants and seeds were selected on agar plates containing antibiotics, then grown in conventional soil.
Particle Inflow Gun
For transient plant transformations a home built gene gun was used employing tungsten as DNA-carrier, following the protocol of Raffaele, et al. [15]. Helium gas was used as kinetically incitement for a successful bombardment of intact mesophyll tissues with DNA-coated tungsten particles. Transient gene expression was detectable within 16- 24 h post bombardment; following the gene bombardment tissues were incubated in pure water in the dark at room temperature.
Protoplast Generation
Leaves of 8 to 10 week old transgenic eGFP::StRem1.3 A. thaliana col 0 plants were cut into small sections (1 x 1 cm) and were incubated for up to 3 h in digestion buffer at 28°C in an incubation shaker (30 rpm). The digestion buffer contained cellulase (0.8% w/v; Onozuka R10; Onozuka, Yakult Pharmaceutical Industry; Tokyo, Japan), pectolyase (0.1% w/v; Sigma Aldrich; Taufkirchen, Germany), bovine serum albumin (0.5% w/v) and polyvinylpyrolidone (0.5% w/v); finally calcium chloride was added (1 mM). The osmolarity was adjusted to 280 milliosmol/kg using sorbitol; the pH was adjusted to 5.6 (MESTris). Protoplasts were separated by centrifugation (80-100 rpm, 10 min, 4°C) from undigested leaf leftovers. After the centrifugation step protoplasts were carefully transferred into a suited vessel and stringently kept on ice.
Fluorescence Microscopy
Images were obtained using confocal microscopes (Zeiss LSM5 Pascal; Carl Zeiss Microimaging, Jena, Germany or TCS SP5; Leica, Mannheim, Germany).
Fluorescence recovery after photo bleaching (FRAP) acquisition and analysis
The FRAP-data were acquired with either the Leica LASAF FRAP Wizard or Zeiss LSM5 Pascal FRAP program, under the imaging conditions listed above. The bleach area on the surface of the plasma membrane was always a 1.5 μm diameter disc. The data could be acquired every 0.25 seconds but were reduced to 5 seconds between analysis point for the purposes of this manuscript. The images were analyzed with Image J (NIH, USA) and the data was exported to Origin (Origin Labs, USA). Average fluorescence intensities within ROIs in the bleached regions were analyzed to obtain the recovery data. Corrections were made for photobleaching during scanning by monitoring neighboring cells and analyzing their signals. Mobile fractions were calculated and the recovery was fit to the equation:
I=A1 - A2*exp(-k*t)
Where I is the average intensity, k is the rate of the exponential recovery, t is the time, A1 is the full recovery value and A2 is the value of the drop in intensity after the bleach. The lateral diffusion coefficients, D, were determined from

Where β is the bleach depth (determined to be 1.2), ω is the radius of the bleach area and k is the fitted rate constant.
In the transgenic Arabidopsis line eGFP:StRem1.3 was expressed in leaves (Figure 1A), guard cells (Figure 1B and 1C) and stems (Figure 1D and 1E), exhibiting the Remorin-typical patchy pattern throughout the plasma membrane. In roots there was no eGFP-specific signal detectable, which could be traced back to a weak expression level of the fusion protein. In native Arabidopsis plants group 1b Remorins are considerably expressed in roots [16] the lack of fluorescence in root tissues was a consequence of the place of insertion of Agrobacteriums´ tumor-DNA into the Arabidopsis genome and not necessarily to physiological means. The overexpression of StRem1.3 in Arabidopsis neither led to any obvious phenotype nor to any subcellular structural abnormalities. With fluorescently tagged group 1b Remorins as in vivo marker proteins for nanodomains there was no lateral movement of individual domains detectable in time periods of up to 60 minutes (not shown). To get some clues concerning the molecular organization of individual Remorin proteins within nanodomains fluorescence recovery after photobleaching (FRAP)-experiments were performed using transgenic Arabidopsis eGFP::StRem1.3 lines. FRAPexperiments in intact mesophyll tissue revealed that the fluorescence of the fusion protein did not recover over a time period of more than 120 seconds, which suggested that individual eGFP::StRem1.3 proteins are proper immobile within plasma membrane nanodomains. This situation dramatically changed upon removal of cell wall components by making use of proteolytic enzymes (see Material and Methods section), which enabled a complete digestion of cell wall components surrounding the plasma membrane to create protoplasts. Upon the creation of protoplast (Figure 2A), the same patchy pattern of green fluorescence could still be observed (Figure 2B) (and with red chlorophyll fluorescence (Figure 2C) as in the intact plants (Figure 1A-1D). However, fluorescence recovery of eGFP::StRem1.3 proteins in protoplasts began immediately after the bleaching (Figure 3B). The relative half maximal fluorescence recovered in 106 seconds. Within the documented time frame of 130 sec fluorescence recovered to a relative value of t130=0.63 (t0=0.15), which reflected a recovery rate of 69% (Figure 3B). The diffusion coefficient [D] was calculated to be about 5.14×10-4 μm2/s with a mobile fraction of 0.62 (immobile fraction: 0.38). These results indicated that the majority of fluorescent Remorin proteins were laterally mobile within plasma membrane nanodomains in protoplasts. At the same time individual Remorin-tagged domains remained their spatial stability in the plane of the membrane (Figure 3B, insets). FRAP-experiments revealed that cell wall components had a stabilizing influence on the mobility of individual Remorin proteins within a single nanodomain. Since Remorins of the taxonomic group 1b do not contain any known transmembrane motifs [17], these proteins must be attached to the plasma membrane from the cytosolic side [1]. The c-terminal anchoring of StRem1.3 has also been demonstrated by Perraki et al. [17], Therefore direct interactions with stable extracellular structures seemed very unlikely. Nevertheless it has been shown for StRem1.3 ex vivo that this protein is able to bind oligogalacturones as well as fractionated pectin - these compounds are protein components of cell walls [18].
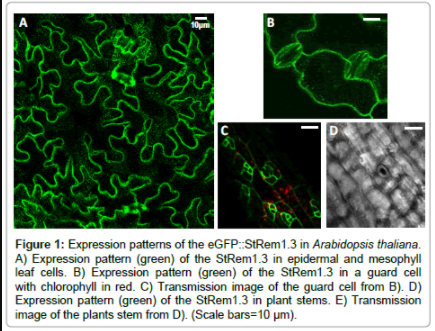
Figure 1: Expression patterns of the eGFP::StRem1.3 in Arabidopsis thaliana. A) Expression pattern (green) of the StRem1.3 in epidermal and mesophyll leaf cells. B) Expression pattern (green) of the StRem1.3 in a guard cell with chlorophyll in red. C) Transmission image of the guard cell from B). D) Expression pattern (green) of the StRem1.3 in plant stems. E) Transmission image of the plants stem from D). (Scale bars=10 μm).
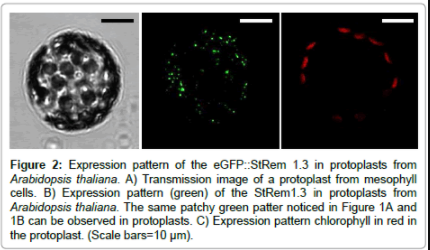
Figure 2: Expression pattern of the eGFP::StRem 1.3 in protoplasts from Arabidopsis thaliana. A) Transmission image of a protoplast from mesophyll cells. B) Expression pattern (green) of the StRem1.3 in protoplasts from Arabidopsis thaliana. The same patchy green patter noticed in Figure 1A and 1B can be observed in protoplasts. C) Expression pattern chlorophyll in red in the protoplast. (Scale bars=10 μm).
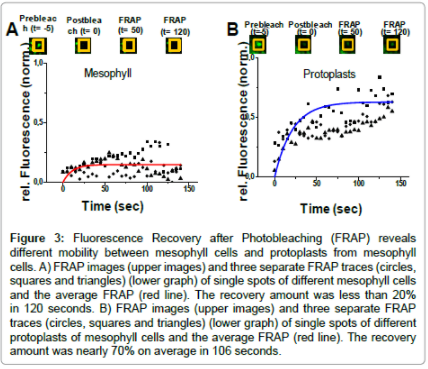
Figure 3: Fluorescence Recovery after Photobleaching (FRAP) reveals different mobility between mesophyll cells and protoplasts from mesophyll cells. A) FRAP images (upper images) and three separate FRAP traces (circles, squares and triangles) (lower graph) of single spots of different mesophyll cells and the average FRAP (red line). The recovery amount was less than 20% in 120 seconds. B) FRAP images (upper images) and three separate FRAP traces (circles, squares and triangles) (lower graph) of single spots of different protoplasts of mesophyll cells and the average FRAP (red line). The recovery amount was nearly 70% on average in 106 seconds.
Remorins of the taxonomic group 1b are invariably associated with membranes. Since Remorins are exhibiting hydrophilic profiles lacking known transmembrane motives, group 1b Remorins must be and have been demonstrated to be attached to the plasma membrane from the cytosolic side [15-17]. FRAP-experiments revealed that components of the cell wall have a stabilizing effect regarding the mobility of single Remorin proteins within individual nanodomains. Suggestions of this general concept have been made previously [19] but not for steroldependent plasma membrane proteins that compartmentalize. In intact mesophyll tissue single eGFP::StRem1.3 fusion proteins did not show a fluorescence recovery (Figure 3A). Referring to the data a majority of StRem1.3 proteins appeared to be proper immobile when the plasma membrane is surrounded by an intact cell wall structure. In protoplasts cell wall components were enzymatically removed. As a consequence the majority of eGFP::StRem1.3 proteins were mobile and showed a fluorescence recovery reaching half of the relative initial fluorescence intensity (half maximal fluorescence) after 106 seconds, with an diffusion coefficient [D] of about 5.14 × 10-4 μm2/s. The mobile fraction was calculated to 0.62 (Figure 3B). To our knowledge no similar datasets exists at this time concerning FRAP-experiments with sterol-dependent plasma membrane proteins in Arabidopsis. However, we note that others have had similar results of other plasma membrane proteins in plants [19]. In yeast FRAP, experiments were performed involving the sterol-dependent proteins Pma1 and Sso1, which are forming patchy-patterns in vital plasma membranes. Pma1 is a proton- ATPase from Saccharomyces that is accumulated in sterol-rich plasma membrane compartments. FRAP-experiments with a fluorescently labelled version of Pma1 in yeast revealed a mobile fraction of 0.77 in plasma membrane compartments. The half maximal fluorescence recovery was reached after 130 seconds with a bleach spot size of about 0.5 μm in diameter [20]. Sso1 in contrast is a yeast SNARE-protein involved in mediating membrane fusions, for instance membrane fusions between secretory vesicles and the plasma membrane [21]. FRAP-experiments in living yeast cells revealed a diffusion coefficient [D]=2.5×10-4 using a GFP-tagged version of Sso1. The half maximal fluorescence recovery was reached after more than one minute. Using cells of the ergosterol synthesis mutant erg6 diffusion of Sso1 was twofold faster compared to wild type cells, which suggested that the physical properties of the lipid bilayer - especially accumulations of sterols - would have a direct influence on the diffusion coefficients of membrane proteins [22]. Interestingly plasma membrane compartments in yeast were laterally immobile in the plane of the membrane, as were plasma membrane nanodomains in Arabidopsis. In yeast it was revealed that components of both, the cytoskeleton and the cell wall, had no effects on the distribution pattern or the temporal stability of individual subcompartments in the plasma membrane, occupied by fluorescently tagged raft proteins. At the same time most of the Pma1::GFP proteins used to label raft-based membrane compartments were laterally mobile within their lipid subcompartment, evidenced by FRAP-experiments [20]. The immobility and maintenance of raft-based membrane subcompartments in yeast, which obviously do not require interactions with stable subcellular structures for their persistence, was explained by the self-organization of sterol-enriched, liquid ordered phases to form stable membrane domains in artificial membranes, where cholesterol containing membrane domains of several micrometers diameter can exist for up to hours. The yeast typical plasma membrane lipid ergosterol is in addition forming the most stable raft-based microdomain of all tested sterol derivates [23]. Based on the amount of identified raft proteins in yeast, an estimated 80% of the yeast plasma membrane could exist as liquid ordered phase and allow for the maintenance of raft-based plasma membrane subcompartments [20]. In Arabidopsis ergosterols are by far less abundant, being replaced by increased levels of sitosterol and stigmasterol as the dominating plasma membrane sterols [24]. In addition there is only an estimated one third of Arabidopsis plasma membrane proteins reported to exhibit strong sterol-dependencies [5] a basic requirement of proteins for possible raft-associations [11]. In plants associations of plasma membrane proteins or whole protein assemblies with stabilizing structures could therefore overcome decreased sterol levels by, for example, interactions or stabilization with the cytoskeleton [7]. There were less cytoskeletal proteins found in sterol-rich membrane domains in plants compared to those found in animal plasma membrane preparations; at the same time plant cell wall proteins made up the biggest functional group of proteins identified as responding to methyl-ß-cyclodextrin [5]. Conformably it is still under debate if plant cell wall proteins anchor the cell wall to the plasma membrane in a similar way as mammalian cells anchor their cytoskeleton [5,25,26].
We thank Fatih Demir for stimulating discussions on the topic and Claudia Horntrich for her assistance with protoplast preparations and maintenance of transgenic plant lines. This work was funded by the German Science Foundation (DFG) grants GK 1342 for the Graduate College of Lipid Signaling in Plants and for FZ-82 for the Rudolf Virchow Center of Excellence in Biomedine to GSH and JOB, by a Howard Hughes Medical Institute Grant to GSH and grants from the Ben Franklin and Northeast Pennsylvania Technology Institute to GSH.