Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2014) Volume 3, Issue 2
Keywords: Streptomyces coelicolor, Tyrosine, Phosphorylation, LMW-PTP, Physiology, Antibiotic production
ACT: Actinorhodin; DCW: Dry Cell Weight; Exp: Exponential; LMW-PTP: Low Molecular Weight Protein Tyrosine Phosphatase; Oxp: Overexpression; PNPP: Para-Nitrophenyl Phosphate; RED: Undecylprodigiosin; WT: Wild Type; μmax: Maximum Specific Growth Rate; μexp: Maximum Specific Growth Rate in Exponential Growth Phase; μRED: Maximum Specific Growth Rate in the RED Producing Phase; rS: Specific Glucose Uptake Rate; q: Volumetric Productivity; YS/X: Yield of Biomass on Glucose
Protein phosphorylation is one of the most prevalent covalent posttranslational modifications and has tremendous regulatory and signaling potential [1]. Phosphorylation on proteins is well studied as a key regulatory mechanism in eukaryotes [2]. Eukaryotic cells rely extensively on phosphorylation of the hydroxyl group of the side chains of serine, threonine and tyrosine for their signal transduction cascades [3]. Bacterial genomes encode protein kinases and phosphatases, but their targets in prokaryotes have been less extensively studied until recently. Initial studies suggested that thr/ser/tyr phosphorylation is more common in eukaryotic signal transduction, while prokaryotes use his/asp phosphorylation in their two-component systems [4]. However, over the last two decades evidence for thr/ser/tyr phosphorylation in bacteria has emerged [5]. Through recent studies that have focused on deducing the entire phosphoproteomes of bacterial species the existence of proteins phosphorylated on thr/ser/tyr has been well established [6-8]. Bacteria possess kinases and phosphatases that structurally resemble their eukaryotic counterparts [9] but have also developed idiosyncratic kinases and phosphatases without known homologues in eukaryotes [7].
Some of these tyrosine kinases and phosphatases have been characterized in details [10]. There are three different classes of phosphotyrosine phosphatases found in bacteria namely; protein tyrosine phosphatases (PTPs), dual-specificity phosphatases (DSPs) and the low-molecular-weight phosphatases (LMW-PTPs) [9]. Grampositive bacteria also contain another class of PTPs in addition to LMW-PTPs, which resemble the phosphoesterase domain of DNA polymerase and histidinol phosphate (PHP) phosphoesterases. Bacillus subtilis encodes one such phosphatase named PtpZ [11]. Among the different classes, LMW-PTPs have been most often shown to control important physiological events. In Staphylococcus aureus and Klebsiella pneumonia these phosphatases are involved in production of a capsular polysaccharide, as a mechanism to overcome host defense mechanism during infection [6], while in B. subtilis LMW-PTPs are involved in the stress response [12] and production of teichuronic acid [13].
The soil dwelling bacterium Streptomyces coelicolor A3(2) has long been used as a model organism to study secondary metabolism in actinomycetes. The genome sequence of S. coelicolor revealed the presence of 44 ser/thr protein kinases [14]. Some of these ser/thr kinases have been studied in details and resemble eukaryotic kinases [15]. Later bioinformatic studies have been carried out to explore phosphatases in the S. coelicolor and S. avermitilis genomes. Each species was then shown to contain at least 55 eukaryotic type protein phosphatases that belong to four different families (those mentioned above) [16].
Existence of proteins phosphorylated on tyrosine in S. coelicolor has been documented by Waters et al. [17]. The phosphotyrosine protein phosphatase gene (ptpA) from S. coelicolor A3(2) was the first eukaryotic-type PP to be discovered in Streptomyces. PtpA belongs to the family of LMW-PTPs [18]. Its functional analysis demonstrated that the disruption of the ptpA gene had no observable effect on cell growth, formation of aerial mycelium and spores, or secondary metabolism in S. coelicolor A3(2). However, overexpression of the ptpA gene, which was carried out in Streptomyces lividans, increased the production of ACT and RED [19]. This led to a hypothesis that there might exist another tyrosine phosphatase that complements the function of ptpA in a ptpA disruption mutant. Shi and Zang [16] predicted gene sco3700 (annotated as a heavy metal reductase) to be a LMW-PTP by bioinformatic analysis of phosphatases in S. coelicolor A3(2), but no functional verification was performed.
In the current study we cloned sco3700 in E. coli and purified the protein. We confirmed the suggested tyrosine phosphatase activity of Sco3700 by biochemical studies. Furthermore, ptpA and sco3700 overexpression strains of S. coelicolor A3(2) were constructed in order to study the effects on physiology. Here we report a physiological characterization of these strains in microtiter plates and the effect of ptpA and sco3700 overexpression on antibiotic production in S. coelicolor A3(2).
Materials
Solvents used were HPLC grade and all other chemicals were analytical grade, unless otherwise stated, and purchased from Sigma– Aldrich (Steinheim, Germany).Water (MQ) used was purified from a Milli-Q-system (Millipore, Bradford, MA). Ni-NTA agarose was obtained from Qiagen (Hilden, Germany) and used for purification of 6X-His tagged Sco3700. All the primers used in this study were obtained from Sigma-Aldrich (Steinheim, Germany). Expand high fidelity Taq polymerase purchased from Roche diagnostics (Basel, Switzerland) was applied for PCR intended for cloning. Taq polymerase purchased from Sigma-Aldrich (Steinheim, Germany) was applied for analytical purposes. Restriction enzymes were obtained from New England Biolabs (Ipswich, Massachusetts).
Strains and plasmids
E. coli DH5α strain was used for gene cloning. A wild type S. coelicolor A3(2) strain (SCP1- and SCP2-) obtained from John Innes Center, UK was used as wild type strain. E. coli strain ET12567/pUZ8002 [20] was used for introducing the recombinant DNA into S. coelicolor A3(2) by conjugation. Integrative plasmid pIJ10257 containing the ermE* promoter was used for overexpression of tyrosine phosphatases in S. coelicolor A3(2) and was obtained from John Innes Center, UK. Commercial plasmid pQE30 available from Qiagen [21], encoding a C-terminal 6x-His tag and an IPTG inducible promoter was used to express sco3700 in E. coli.
Media and growth conditions during strain construction
E. coli cultures for molecular biology purposes and for protein purification were grown at 37°C in Luria Bertani broth [22]. Hygromycin (50 μg/ml) was used for E.coli DH5α with plasmid pIJ10257. Kanamycin (25 μg/ml) and choloramphenicol (25 μg/ml) were used for E.coli ET12567/pUZ8002. Nalidixic acids (20 μg/ml) were used for E. coli and hygromycin (50 μg/ml) were used for S. coelicolor ptpA and sco3700 overexpression strain selection.
Mannitol-Soya flour (MS) agar was used for plating of S. coelicolor. 2X YT medium was used for germination of spores and contained 16 g/L tryptone, 10 g/L yeast extract and 10 g/L sodium chloride. S. coelicolor strains were incubated at 28°C.
DNA techniques
DNA techniques involving E. coli were performed as described by Sambrook et al. [22]. E. coli transformations were done using electroporation. Chromosomal DNA from S. coelicolor was isolated as described by Kisser et al. [23]. S. coelicolor spores were streaked on MS agar plates to obtain dense masses of spores. These spores were isolated in 20% glycerol and used for conjugation.
Strain construction
Overexpression strains of ptpA and sco3700 of S. coelicolor were constructed by cloning the genes into vector pIJ10257 containing the ermE*promoter and XhoI and NdeI restriction sites. PtpA and sco3700 genes were amplified using primers as shown in Table 1. The forward primers in both cases possessed XhoI restriction site while the reverse primers possessed NdeI restriction site. The recombinant plasmids pIJ10257_ptpA and pIJ10257_sco3700 were then electroporated into E. coli ET12567/pUZ8002 and eventually conjugated into S. coelicolor. Hygromycin resistant colonies of S. coelicolor were selected on MS agar plates and propagated further.
| Name of primer | Sequence |
|---|---|
| Fw_ptpA_XhoI | TATTCTCGAGTCATGCCGCCCGTCCTTCCAC |
| Rv_ptpA_NdeI | CGTCGCATATGATGACCTACCGCGTCTGTTTC |
| Fw_sco3700_NdeI | TGCGACATATGATGCCCGACAAGCCTTCCGTGCTCTTCGTC |
| Rv_sco3700_XhoI | GATTCTCGAGTCATGACTGCGGCCGCGGCGCGATC |
| pQE30_sco3700_fw | TACGAGGATCCATGCCCGACAAGCCTTCCGTGCTC |
| pQE30_sco3700_rv | GATCTAAGCTTTCATGACTGCGGCCGCGGCGCGATC |
Table 1: Sequences of primers for over-expressed and delta ptpA and sco3700 strains of S. coelicolor.
Cloning of sco3700 in pQE30
The gene sco3700 was amplified using a forward primer containing BamHI restriction site and a reverse primer containing HindIII restriction site (Table 1) and then cloned into plasmid pQE30 using BamHI and HindIII restriction sites as described in The QIA expressonist manual to obtain pQE30_Hissco3700. This plasmid was then used to transform E. coli M15 cells [21]. Ampicillin resistant colonies were selected.
Purification of 6XHis-tagged Sco3700 from E. coli
Purification of 6X-His tagged Sco3700was performed from 1 liter culture. E. coli cells were grown and IPTG used for induction as described in the protocol 8 from The QIA expressionist manual. After induction, the cells were harvested by centrifugation at 4000 g for 20 minutes. The cell pellet was suspended in 5 ml Solution A (50mM Tris-Hcl pH 7.5, 100 mM NaCl and 10% glycerol) containing 1mg/ml lysozyme and 5 μg/ml DNase I and incubated at room temperature for 15 minutes. The cells were then lysed by sonication on ice. The lysate was centrifuged at 25000 g for 15 minutes to get rid of cellular debris and the crude protein extract was obtained. Ni-NTA agarose used for protein purification was obtained from Qiagen (Venlo, Netherlands) and purification was carried out following the manufacturer’s instructions. Fractions showing maximum protein were subjected to desalting using PD10 columns obtained from GE-Healthcare (Buckinghamshire, U.K.). Desalting was performed as per the manufacturer’s instructions. Qualitative determination of protein was performed using the Bradford reagent (Biorad, California, USA). The fractions containing maximum protein were pooled and used for assays.
Biochemical characterization of Sco3700
Optimum pH for Sco3700 was determined by determining phosphatase activity of Sco3700 using incubation buffers of different pH. Sodium citrate buffers of pH 5.5, 6.0, 6.5 and 7.1, and phosphate buffers of pH 7.5, 8.1, 8.5 and 8.9 were tested. 100 mM PNPP (paranitrophenyl phosphate) was used as a substrate as described previously [24]. KM and Vmax for Sco3700 were determined using PNPP of different concentrations (5-30 mM with intervals of 5 mM) at 30°C. Enzyme activity was monitored over 60 minutes, by measuring the formation of p-nitro phenol at 405 nm.
Tyrosine phosphatase activity of Sco3700 was confirmed using B. subtilis PtkA (phosphotyrosine kinase) [25] that autophosphorylates on tyrosine in presence of ATP. In the assay, PtkA was allowed to autophosphorylate for 15 minutes at 30ºC by incubating with 10 mM ATP and 10 mM MgCl2. Sco3700 was then added to the above mixture and the release of phosphate was measured at 635 nm using Pi colorlock ALS kit from Innova Biosciences (Babraham, U.K.).
Frozen mycelia preparation of ptpA and sco3700 overexpression strains of S. coelicolor
Frozen mycelia were used as inoculum for all the cultivations in minimal medium. Frozen mycelia stocks were prepared as described by Sohoni et al. [26]. Spores for respective strains were inoculated into 50 ml 2X YT medium in shake flasks and incubated at 28ºC and 150 rpm. Mycelia were harvested in mid-exponential phase and centrifuged at 4ºC for 5 minutes, after which the supernatant was discarded. 2-3 ml of 20% pre-sterilized peptone was added to the pellet and mycelia were crushed using a glass homogenizer. 7-8 ml of 20% peptone was added to the crushed mycelia and distributed into cryo vials. Vials were stored at -20ºC and used for inoculation in minimal medium.
Physiological characterization of ptpA and sco3700 overexpression strains of S. coelicolor
Physiological characterization of the strains was performed in 24 deep square well microtiter plates with 3 ml working volume at 28°C and 150 rpm as described by Sohoni et al. [26]. Phosphate limited minimal medium modified from Evan’s medium was prepared as described by Borodina et al. [27]. The medium contained 30 g/L glucose, 3 mM phosphate and 100 mM ammonia. In addition, the medium also contained 100 mM MOPS buffer. Glucose was sterilized separately and mixed with the rest of the medium later. Medium pH value was adjusted to 6.85 after sterilization using 4 M NaOH. 80 ml of the medium was inoculated with 80 μL frozen mycelia and then 3 ml was distributed in each well in a microtiter plate. At each time point duplicate samples were taken from plates and OD450nm, dry cell weight (DCW), pH, glucose consumption and RED and ACT synthesis were monitored. All the analyses were carried out as described by Borodina et al. [27].
Analysis of phosphate in the broth
The Pi Color Lock TM ALS Kit (Innova Biosciences, Cambridge, UK) was used for the analysis of phosphate in the cultivation broth at different time points. The measurement of Pi in this kit is based on the change in absorbance of malachite green in the presence of molybdate and gives a sensitive detection of Pi.
The 0.8 mM Pi standard (provided by the kit) was diluted to the end concentrations of 0, 40, 80, 120, 160 and 200 μM. The samples were diluted to the phosphate concentration of 20-120 μM phosphate per assay. 200 μl ALS mix (provided by the kit) were added to 50 μl sample (diluted) and incubated for 5 minutes at 30°C. After adding 20 μl of stabilizer (provided by the kit) the mixture was incubated again for 30 minutes at 30°C. 1 ml of distilled water was added to each sample and the A635 was measured.
This study focuses on LMW-PTPs from S. coelicolor A3(2) and their role in regulating the secondary metabolism. One of the LMWPTPs, PtpA has previously been identified and shown to influence production of the antibiotics RED and ACT [18,19]. However, this is the first biochemical and physiological characterization of a second low molecular weight tyrosine phosphatase, Sco3700, present in S. coelicolor.
Biochemical characterization of Sco3700
In many bacteria the presence of two functionally active LMWPTPs has been reported [12,13,28]. In S. coelicolor so far only one such LMW-PTP, PtpA, has been isolated and characterized [18]. In this study we biochemically confirmed Sco3700 as being a LMW-PTP, which was previously suggested by bioinformatic studies by Shi (2004) but never functionally proven. 6xHis-tagged Sco3700 purified from E. coli was used for biochemical characterization. Sco3700 was able to hydrolyze the artificial substrate PNPP, with the KM value of 14.3 mM and pH optimum at 6.5 (Figure 1a and 1b). The KM value of PtpA for PNPP has earlier been reported to be 0.75 mM [18]. Mycobacterium tuberculosis, also belonging to the group of actinomycetes, encodes one LMW-PTP namely mptpA [29]. B. subtilis PTPs have the highest reported substrate affinity with KM of 0.156 mM and 0.25 mM for PtpA and PtpB, respectively [12], while an E. coli Ptp, Wzb, has been reported to have a KM of 1 mM [28]. S. aureus PtpA and PtpB have KM of 1.2 mM and 1.4 mM, respectively [30]. These values are comparable with the KM reported for S. coelicolor PtpA, but the KM of Sco3700 is 20 times higher, possibly reflecting that the two tyrosine phosphatases in S. coelicolor act at very different ranges of substrate concentration in the cell. PNPP is an artificial substrate and a small molecule, commonly used for detection of tyrosine phosphatase activity due to easy colorimetric determination of p-nitrophenol formed in the reaction after cleavage of phosphate from PNPP. To further verify the functionality of Sco3700 on a real protein substrate, we also tested the phosphotyrosine-protein phosphatase activity of Sco3700 using PtkA, a tyrosine phosphorylated protein from B. subtilis. The detected liberation of free phosphate from phosphorylated PtkA (data not shown) confirmed the capacity of Sco3700 to dephosphorylate tyrosine-phosphorylated proteins.

Figure 1: Biochemical characterization of Sco3700
In many bacterial genomes PTPs are located next to protein tyrosine kinases (TKs) [12,31,32]. In Actinomycetes (Mycobacterium spp and Streptomyces spp.) despite the presence of tyrosine phosphatases, no tyrosine kinases have been evidenced so far [33]. Classical bacterial tyrosine kinases of the BY-kinase family [10] are absent in Actinomycetes, and it is therefore possible that these bacteria harbor a presently unidentified family of tyrosine kinases or simply possess serine/threonine kinases with relaxed specificity, capable of phosphorylating tyrosine residues.
Physiological roles of tyrosine phosphatases in bacteria
LMW-PTPs have been shown to have important physiological roles in bacteria. Wzc in E. coli was found to be similar to proteins responsible for synthesis and export of exopolysaccharides that take part in capsule formation, which is an important mechanism in virulence [28]. Ptp of Acinetobacter johnsonii has also been predicted to be involved in exopolysaccharide formation [32]. In addition, as mentioned above, PTPs of S. aureus and K. penumoniae take part in capsule formation and hence in pathogenicity. PtpA from M. tuberculosis has also been shown to serve as a virulence factor while infecting macrophages [29,33]. Furthermore, deletion and over-expression of tyrosine phosphatases resulted in altered polysaccharide biosynthesis in a range of bacteria and hence, these phosphatases might be attractive targets for the development of novel anti-microbials [34]. The role of protein phosphorylation in regulation of secondary metabolism is also well known. The two component systems AbsA1- AbsA2 and AfsK-AfsR that regulate antibiotic biosynthesis in S. coelicolor have been shown to mediate control by reversible Ser/Thr phosphorylation [35,36]. Waters et al. [17] demonstrated changes in the pattern of tyrosine phosphorylation in different Streptomyces species in different phases of growth and differentiation. In S. lividans cultures grown in minimal medium, they reported increased protein phosphorylation in early stationary phase and a specific increase in tyrosine phosphorylation in S. lavendulae in early stationary phase. They also suggested the possibility of tyrosine phosphorylation playing an important role in a switch from primary to secondary metabolism. In this study we examined the impact of the tyrosine phosphatases PtpA and Sco3700 on growth and antibiotic production in S. coelicolor on liquid, minimal medium.
Physiological characterization of ptpA and sco3700 S. coelicolor over expression strains was carried out in microtiter plates. Growth trajectory was followed for 120 hrs and samples were taken from microtiter plates at various time points. The wild type S. coelicolor A3(2) strain served as a control and all the physiological parameters for the two recombinant strains have been compared to the wild type strain.
Physiological effect of ptpA over expression
Growth behavior of S. coelicolor oxp-ptpA was very similar to the WT strain (Figures 2b and 2a), with two growth phases. The first was the exponential phase with a μmax of 0.11 h-1 (same as the wild type), while the specific growth rate in the second growth phase, the REDproduction phase, was slightly higher than for the WT strain (Table 2).
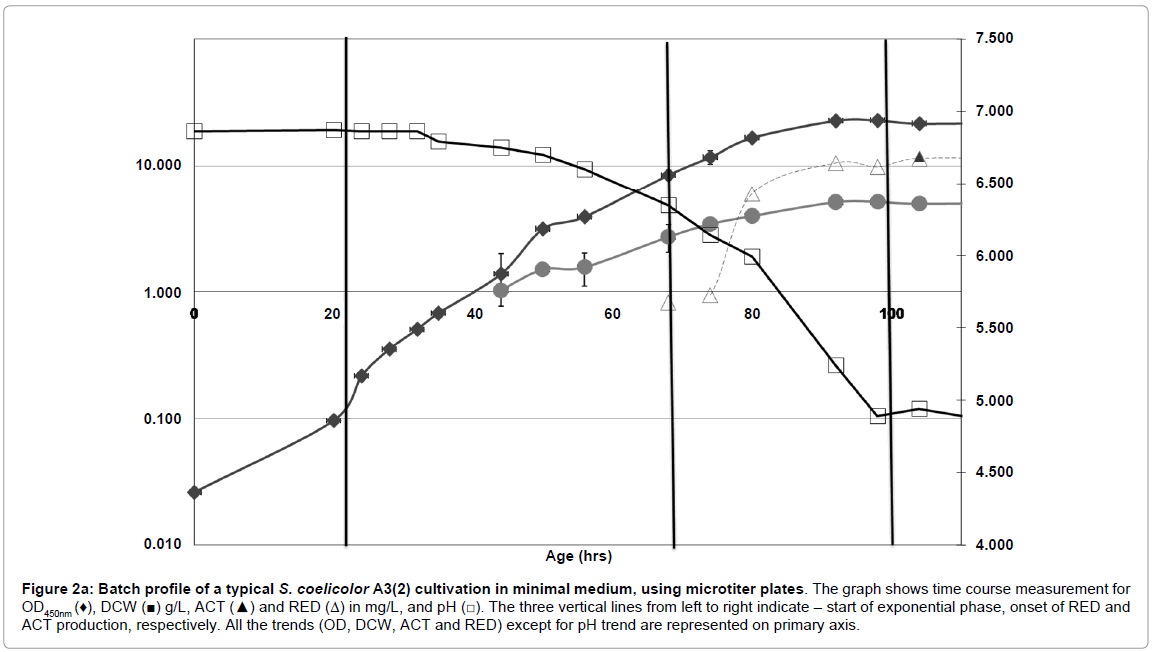
Figure 2a: Batch profile of a typical S. coelicolor A3(2) cultivation in minimal medium, using microtiter plates. The graph shows time course measurement for OD450nm (♦), DCW (■) g/L, ACT (▲) and RED (Δ) in mg/L, and pH (□). The three vertical lines from left to right indicate – start of exponential phase, onset of RED and ACT production, respectively. All the trends (OD, DCW, ACT and RED) except for pH trend are represented on primary axis.
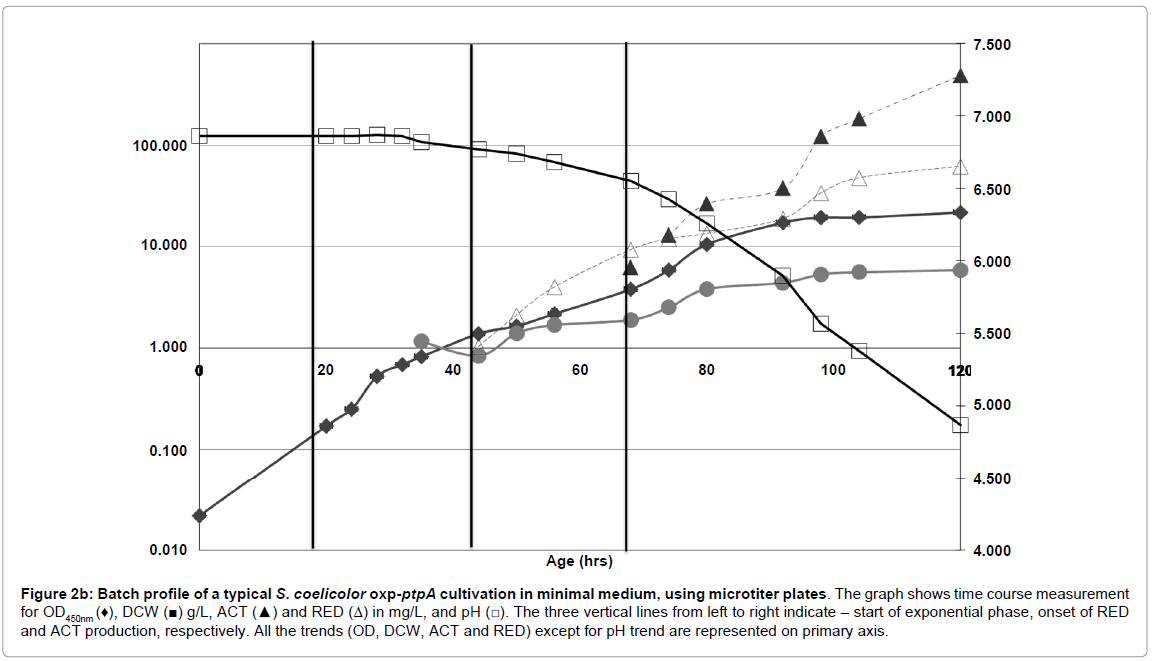
Figure 2b: Batch profile of a typical S. coelicolor oxp-ptpA cultivation in minimal medium, using microtiter plates. The graph shows time course measurement for OD450nm (♦), DCW (■) g/L, ACT (▲) and RED (Δ) in mg/L, and pH (□). The three vertical lines from left to right indicate – start of exponential phase, onset of RED and ACT production, respectively. All the trends (OD, DCW, ACT and RED) except for pH trend are represented on primary axis.
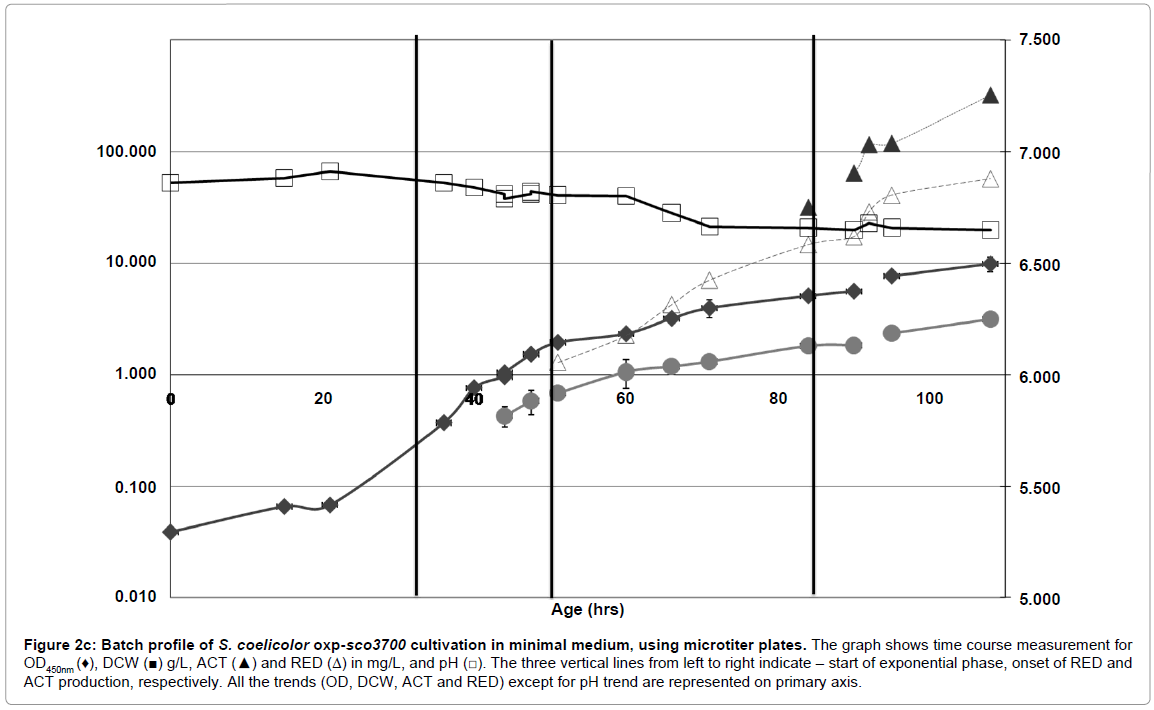
Figure 2c: Batch profile of S. coelicolor oxp-sco3700 cultivation in minimal medium, using microtiter plates. The graph shows time course measurement for OD450nm (♦), DCW (■) g/L, ACT (▲) and RED (Δ) in mg/L, and pH (□). The three vertical lines from left to right indicate – start of exponential phase, onset of RED and ACT production, respectively. All the trends (OD, DCW, ACT and RED) except for pH trend are represented on primary axis.
| Strain | µexp(h-1) | µRED(h-1) | YS/X (g/g) | -rS (g glu/g DCW.h-1) | qRED(g/L/hr) | qAct (g/L/hr) |
|---|---|---|---|---|---|---|
| S. coelicolor A3(2) [WT] | 0.11 ± 0.01 | 0.04 ±0.02 | 0.31 ± 0.02 | 1.8 ± 0.04 | 1.7 ± 0.28 | 12.4 ± 2.4 |
| S. coelicolor oxp-ptpA | 0.11± 0.01 | 0.04 ±0.02 | 0.31± 0.01 | 1.9 ± 0.004 | 1.6 ± 0.01 | 16.1 ± 0.4 |
| S. coelicolor oxp-sco3700 | 0.11± 0.005 | - | 0.29 ± 0.007 | 2.2 ± 0.04 | 0.33 ± 0.03 | - |
Table 2: Comparative table representing physiological parameters for the different strains in this study.
Dry cell weight (DCW) measurements showed a significantly higher biomass concentration for oxp-ptpA (5.6 g/L) than for the WT strain (3.1 g/L) at the end of fermentation (Figures 2b and 2a).
However, the yield of biomass on glucose for oxp-ptpA was found to be comparable to that of the WT strain (Table 2), as was the substrate uptake rate (-rs) (1.8 g glu/g DCW. h-1 for WT, and 1.9 g glu/g DCW. h-1for the oxp-ptpA strain).
In comparison with the S. coelicolor WT strain, there was an early onset of both RED and ACT production in the oxp-ptpA strain (Figure 3). The volumetric production rate for RED in oxp-ptpA (1.6 mg/L/hr) was slightly lower than in S. coelicolor WT (1.7 mg/L/hr), whereas the volumetric production rate for ACT was higher in the oxp-ptpA strain (16.1 mg/L/hr) than for S. coelicolor WT (12.4 mg/L/hr).
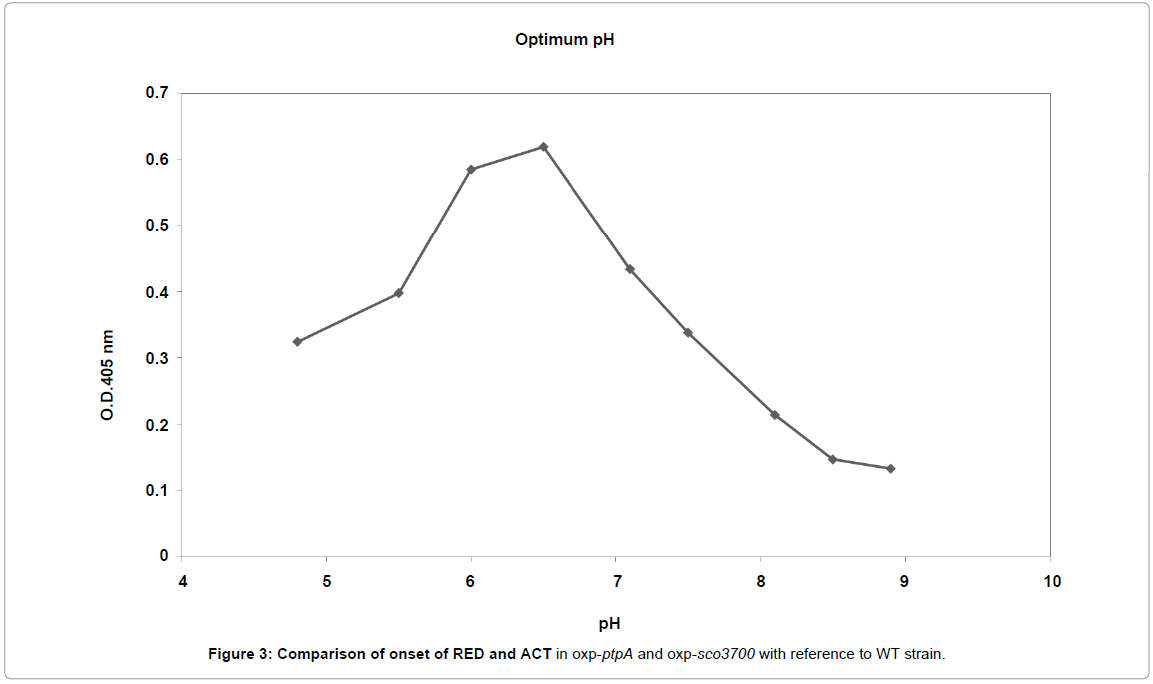
Figure 3: Comparison of onset of RED and ACT in oxp-ptpA and oxp-sco3700 with reference to WT strain.
Physiological effects of sco3700 overexpression
The growth curve of S. coelicolor oxp-sco3700, unlike that of the WT strain, exhibited a sigmoidal shape (Figure 2c). There was only a single growth phase with μmax of 0.11 h-1. As for the ptpA over expression strain, dry cell weight measurements showed significantly higher values for oxp-sco3700 (5.0 g/L) than wild type strain (3.1 g/L) at the end of fermentation. There was also a significant drop in pH from 6.86 to 4.87 for the oxp-sco3700 strain. Despite higher final biomass concentration, the yield of biomass on glucose (YSX) was lower for oxpsco3700 (0.29 g/g) compared to S. coelicolor WT (0.31 g/g) and oxpptpA (Table 2). The lower biomass yield was as a consequence of acid production. HPLC analyses showed presence of acetate, which is likely to be produced by overflow metabolism due to the increased substrate uptake rate. Also differing from what was observed when ptpA was overexpressed, the glucose uptake rate (-rs) was significantly higher in the oxp-sco3700 strain (2.2 g glu/g DCW. h-1) than in the S. coelicolor WT strain (1.8 g glu/g DCW. h-1).
With regards to antibiotic production, an opposite behavior compared to overexpression of ptpA was seen. Onset of antibiotics production (RED and ACT) was significantly delayed in the oxpsco3700 strain. Also the volumetric production rate for RED was pronouncedly lower (0.3 mg/L/hr for the oxp-sco3700 strain) than in S. coelicolor WT (1.9 mg/L/hr) (Table 2). The volumetric production rate for ACT could not be calculated in the oxp-sco3700 cultivations due to the late onset of ACT production (around 100 hrs). The cultivation was terminated at 120h since wall growth started to appear in oxp-sco3700 cultivation.
Comparison of physiological behavior of the ptpA and sco3700 over expression strains and possible regulatory effects
Dephosphorylation of proteins can mediate both activation and inactivation of target proteins, and by consequence physiological functions [37]. Overexpression of ptpA and sco3700 affected antibiotic production in S. coelicolor differently. Even though the oxp-ptpA strain behaved similarly to the WT strain, it exhibited earlier onset of antibiotics production and increased volumetric production of ACT. Interestingly, oxp-sco3700 exhibited a very atypical growth behavior. Antibiotic production was significantly affected in the oxpsco3700strain, with volumetric production rates for RED and ACT severely reduced.
Eukaryotic LMW PTPs play an important role in signal transduction mainly by specifically dephosphorylating and downregulating tyrosine kinase receptors such as the PDGF receptor or insulin receptor [37]. In S. cerevisiae heterologous overexpression of a LMW-PTP, Stp1, (from Schizosaccharomyces pombe) resulted in a large number of phenotypes that indicated down-regulation of the Ras pathway [38]. These phenotypes included reduction in both cAMP signaling and GTP loading of Ras2, impaired growth on nonfermentable carbon source, alteration of cell cycle parameters, delayed recovery from nitrogen starvation, increased heat shock resistance and attenuation of invasive growth. This suggests that dephosphorylation of proteins by LMW-PTPs may lead to global changes in the cell. LMWPTPs from different bacteria have been characterized at biochemical level, but studies on their overexpression are scarce.
The observed effects of ptpA and sco3700 on S. coelicolor cell are likely to be at the regulatory level, but one cannot exclude direct metabolic effects. Antibiotic production in actinomycetes is believed to be a result of stress or nutrient limitation. In this study we used phosphate limitation and hence it was interesting to monitor the phosphate consumption over the period of fermentation. Figure 4 represents time course measurement of phosphate consumption in the different strains with respect to WT strain. The oxp-ptpA and oxp-sco3700 strains consumed phosphate faster than the WT strain (Figure 4). This could be one reason for an early onset of antibiotic production in the oxp-ptpA strain. A plausible hypothesis for the regulatory interaction between LMW-PTPs and antibiotic production would be that PtpA and Sco3700 interact with one or several tyrosinephosphorylated proteins, some of which are implicated in regulation of antibiotic biosynthesis. Accordingly, overexpression of ptpA triggered production of antibiotics early. This type of regulation has been seen for colonic acid production in E. coli. The ca gene cluster in E. coli contains one tyrosine autokinase (Wzc) and one LMW-PTP (Wzb) that function as a pair of kinase/phosphatase in the regulation of colanic acid production. As a consequence, colanic acid is only produced after the dephosphorylation of the phosphorylated Wzc by Wzb [28].
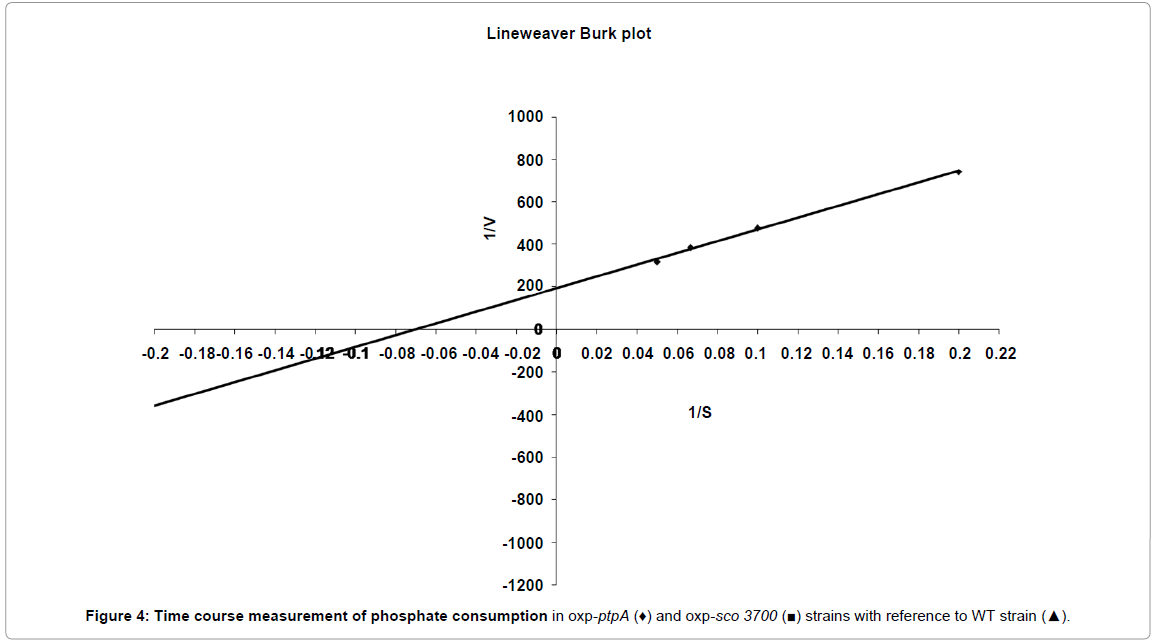
Figure 4: Time course measurement of phosphate consumption in oxp-ptpA (♦) and oxp-sco 3700 (■) strains with reference to WT strain (▲).
Behavior of the oxp-sco3700 strain was unexpected. Even though phosphate in the medium was depleted earlier, onset of both the antibiotics, RED and ACT was delayed. Sco3700 has twenty times higher KM for PNPP than PtpA. Hence, Sco3700 is likely to act on a different set of physiological substrates and be involved in regulatory loops different from those of PtpA. Final responses to these questions will come only with phosphoproteomic studies in S. coelicolor that will assign physiological substrates to all protein kinases and phosphatases.
Sco3700 was biochemically characterized and confirmed to be a tyrosine phosphatase. The enzyme seems to present significant differences to the second earlier described tyrosine phosphatase in S coelicolor PtpA, both in terms of biochemical and physiological characteristics. The oxp-sco3700 recombinant strain turned out to behave very differently compared to the oxp-ptpA and WT strain and affected the production of antibiotics negatively. The significant changes in growth behavior observed in oxp-sco3700 led to the conclusion that overexpression of sco3700 have a pleiotropic effect on the cell. In agreement to the results of Umeyama et al. [19], significant increase in volumetric productivity of ACT was observed but no significant changes in yield was observed when looking at the yield of product per amount of biomass. An early onset of the ACT production could be confirmed and furthermore a higher volumetric productivity was seen. Despite the fact that in the overexpression strain of sco3700 antibiotic production was affected, the growth was not severely affected.
Authors would like to thank the Technical University of Denmark, the Danish Biotechnology Research School and the European Commission (IP005224 ActinoGEN) for financial support during this work. We would also like to acknowledge Dr. Bertolt Gust from University of Tubingen for fruitful discussions on the work.