Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Editorial - (2012) Volume 1, Issue 1
To date, the luciferase in reporter assays and the gateway toolkit are among our routinely- used reagents [1,2], and the latter also facilitates the large-scale gene cloning so that we can switch to pick up ones of interest from the genome-wide ready- to- use collections [3]. However, it was particularity a small number of enzymes that probably have advanced huge successful stories in research of biology and medicine. They actually help us perform the special- pediment DNA cloning, genome editing and transgenics. This has greatly enlightened us. Interestingly, the toolkit equipped with one of them, e.g. Cre/lox (Flp /FRT) recombinase system for gene targeting has already been awarded the Nobel Prize [4]. We would hence like to expect more cases among them in the future.
We now enter the era of post-genome. The Cre/lox and Flp /FRT recombinase systems are among the heroes to help us understand our genome. Nobel laureate Mario Capecchi [4] first conveyed his creative ideas on gene targeting with Cre/lox recombinase system first found in phage to NIH for funding but it was given a rejection.It becomes today well-known that this system can harness the machinery of mammalian cells to precisely mutate any gene we wish. Furthermore, its derivatives such as tissue-specific inducible conditional knock-outs accelerate our understanding of gene functions. For instance, now expression of Cre and hence targeting of the floxed gene can be restricted to e.g. chondrocytes (collagen type II alpha1 promoter, Figure 1), T cells (lck promoter), cardiac muscle (cardiac myosin promoter), neurons (enolase promoter). This technique not only promotes generation of mice with human diseases for study, but it may hold a great promise in-future gene therapy to correct disease-causing genes.
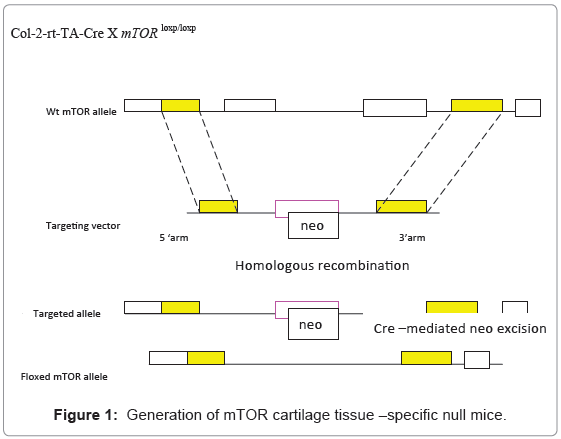
Figure 1: Generation of mTOR cartilage tissue –specific null mice.
Targeting of mTOR
The targeting vector was constructed to replace the endogenous mTOR exon locus with a neo cassette by homologous recombination.
The generality of application of Flp /FRT recombinase system can definitively expand to fruit fly (Drosophila melanogaster) in vitro gene targeting [5] and of the Cre/lox recombinase system extends to in vivo integration of low -copy transgenic Caenorhabditis elegans ( C. elegans ) [6-8].
It is an amazing breakthrough to identify a similarly flexible homologous recombination reaction in Escherichia coli (E. coli) as in the yeast, an assay on the basis of recombination between linear and circular DNA. Only sbcA E. coli strains, which express RecE and RecT, presented the intended recombination products [9,10]. This started a new logic for DNA engineering using recombination in E. coli, unlimited by the disposition of restriction endonuclease cleavage sites or the size of the target DNA and without the need for DNA ligases. This new form of chromosome engineering, termed recombinogenic engineering or recombineering, is efficient and greatly decreases the time it takes to create transgenic mouse models by traditional means [9]. Hence, highly efficient phage-based Escherichia coli homologous recombination systems enable genomic DNA in bacterial artificial chromosomes (BACs) or Fosmids to be modified and subcloned with precision and ease, and to use them for transgenesis in higher model organisms, like mouse [4] and C. elegans [6,7]. Importantly, many reagents are currently commercially available. Recently Poser I et al. [11] have developed the localization, protein-protein and/or protein- DNA interactions of the tagged protein 3-in-1 mammalian system via rapid and reliable generated BAC transgenes using 96-well-format recombineering [12]. Such systems have already been applied on C. elegans and contributed a great to Modencode [13,14]. One of its latest applications includes a conditional knockout resource for the genomewide study of mouse gene function [15]. Similarly, we described the use of galK gene as a selection marker for both the positive and negative selection steps in recombineering, realized placement of homology arms to commonly used GFP and TAP fusion genes flanking the galK cassette [6,7,16].
Finally, recombineering can facilitate many kinds of genomic experiment that have otherwise been difficult to carry out, and should enhance functional genomics studies by providing better mouse and other models and a more refined genetic analysis of the mouse and other genome, from individual(s) to system(s) screen with high throughput [16].
The Cold Fusion technology is a “cool”, revolutionary cloning approach for rapid, efficient and precise direct cloning of any PCR fragments to any linearized vector in one single step, at any site. The PCR fragments can be generated with primers that are designed to have at least 15 bases of homology at their linear ends to where you want the product to “fuse”. You generate the linearized vector by PCR or restriction enzymes (single or double cut). With the mixture of enzymes that prepare the ends of DNA fragments for sequence-directed alignment for “cold” cloning in one tube simple reaction format, you incubate them for 5 minute at room temperature then put them for 10 minutes on ice. Subsequently, your PCR product(s) will rapidly fuse into the linearized vector in the desired orientation because of compatible DNA ends with great accuracy. It is so robust that multiple DNA fragments can be assembled simultaneously and cloned into one construct in a single step. The system is highly efficient along with a rate of more than 95% positive cloning. Obviously, this cloning is also adaptable for rapid high throughput application. Experimentally, this method can have accomplished 4-in-1 induced pluripotent stem cells (iPSC) construct [17] in 3 days.
Zinc finger nucleases (ZFNs) are a class of engineered DNAbinding proteins that facilitate targeted editing of the genome by creating double-strand breaks in DNA at user-specified locations [18]. Simply speaking, each Zinc Finger Nuclease (ZFN) consists of two functional domains: the DNA-binding domain for zinc-fingers, which is engineered to bind to a specific DNA sequence, fused to the DNA endonuclease domain from the FokI [19]. Such fusion makes a highly-specific pair of ‘genomic scissors’ created. When two ZFNs bind cognate target sequences in the proper orientation, the FokI domains can dimerize and create a nuclease that makes a DNA double-stranded break (DSB) between the two cognate sequences. DSBS are important for site-specific mutagenesis to stimulate the cell’s natural DNA-repair processes, namely homologous recombination and Non-Homologous End Joining (NHEJ) [20].
These processes are harnessed to generate precisely targeted genomic edits, resulting in cell lines with targeted gene deletions, integrations, or modifications. ZFN-mediated genome editing takes place in the nucleus when a ZFN pair targeting the user’s gene of interest is delivered into a parental cell line, either by transfection, electroporation or viral delivery. It can make rapid disruption of, or integration into, any genomic loci [20]; Mutations made are generally permanent and heritable in a variety of mammalian somatic cell types. Researchers could obtain knockout or knock-in cell lines. Importantly, you can make knockout of genes not amenable to RNAi.
Human ES cells and iPS cells grow very poorly when plated as single cells. However, now several studies have also reported successful gene targeting by HR-mediated gene insertion at a few selective loci in normal or disease-specific human iPS cells [21,22]. For sure, facile correction of genomic mutations in iPS cells still has a long way to go. As aforementioned, the use of zinc-finger nuclease (ZFN) generated site-specific DSBs to stimulate HR efficiencies. The use of a ZFN pair also increases the overall sequence specificity, enabling them to precisely target a single unique locus in the genome [20]. Stimulated by ZFN-induced DSBs, endogenous loci can be targeted with high efficiency not only by either HR (in the presence of an exogenous donor DNA fragment serving as a repair template) or but also through error-prone nonhomologous end-joining (especially in the absence of a DNA template) [20].
While the use of ZFNs can stimulate HR rates significantly, the limitation of ZFN efficiency had stymied its clinic application, namely, it is still not high enough that the step of gene targeting or correcting can be assumed to be either facile (especially for transcriptionally silent loci) or of short enough duration to be adaptable to a clinical setting[20]. The somatic hypermutation (SHM) could possibly mutate target ZFN to evolve its proteins directly in living mammalian cells [23]. Mutant ZFNs are expressed and selected or screened for desired more modular, higher efficient at targeting human somatic cells, high-level gene transfer for increasing HR and other active merits for avenues of clinic application.
Enzyme Fragment Complementation (EFC) Assay Technology is a homogeneous, non-radioactive detection technology based on two genetically engineered β-galactosidase fragments-a large protein fragment (Enzyme Acceptor, EA) and a small peptide fragment (Enzyme Donor, ED). Separately, the β-gal fragments are inactive, but in solution, they rapidly recombine to form active β-galactosidase enzyme that hydrolyzes substrate; producing an easily detectable chemiluminescent or fluorescent signal. EFC technology offers drug discovery the means to interrogate biomolecular reactions for advancing therapeutic drug and screening programs. Robust, reliable assay technology can be used for both biochemical and cell based formats [24].
For the matrix metalloproteinase MMP for tumor surgery , Nobel laureate Roger Tsien et al. [23] have modified their activatable cell penetrating peptides(ACPPs), which was previously described for in vivo fluorescence localization of active MMP-2 and-9 in xenograft and transgenic tumor models to in vivo visualize the matrix metalloproteinase activities by MRI and fluorescence of dendrimeric nanoparticles coated with activatable cell penetrating peptides (ACPPs), labeled with Cy5, gadolinium, or both. Uptake of such nanoparticles in tumors is 4 to15-folds higher than for unconjugated ACPPs. They proposed that that work could improve MRI-guided clinical staging, presurgical planning, and intraoperative fluorescence-guided surgery. The approach may be generalizable to deliver radiation-sensitizing and chemotherapeutic agents [25]. In addition, MMP-2 and MMP-9 is also critical for osteoarthritis [26] and stemness.
Finally, these enzymes could hence make our research in biology and biomedicine fruitful and full of exciting surprises now and in the future. However, we believe that development of such enzymes and their toolkits on many fronts will be an intense field of research in the near future, and their progress will have a long -term impact not just in basic biological research, but also cancer research, regenerative medicine, drug screening and disease modeling.