Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2015) Volume 6, Issue 3
Species composition, population densities and size of jellyfish and ctenophores were recorded during 5 cruises in the heavily eutrophicated Limfjorden in 2014. No or very few ctenophores (Pleurobrachia pileus) and jellyfish (Aurelia aurita, Cyanea lamarckii) were recorded in April and June 2014, whereas in August and September numerous small individuals of the invasive ctenophore Mnemiopsis leidyi were found on all 4 locations studied, which were strongly reduced in population density during November. M. leidyi exerted a notable predation impact, most pronounced in Løgstør Bredning and Skive Fjord in August when the estimated half-lives of zooplankton were 4.8 and 7.3 d, respectively, and in late September, when the half-life in Skive Fjord was only 2.2 d. Severe oxygen depletion in Løgstør Bredning and Skive Fjord between June and September resulted in a release of nutrients. This was followed by a bloom of the dinoflagellate Noctiluca scintillans and a subsequent peak in the abundance of copepods which decreased rapidly after the introduction of M. leidyi into Limfjorden from the North Sea (between early April and mid- July) to become virtually absent during the rest of the season. This subsequently resulted in starvation and decay of the M. leidyi population. The small predatory ctenophore Beroe gracilis was recorded on most locations during August and September 2014 but although B. gracilis eats small M. leidyi, their low number suggested a negligible predation impact on the M. leidyi population. Our present understanding of the many biological and environmental factors that control the species composition, abundance and predation impact of jellyfish and ctenophore populations in Limfjorden are discussed. It is concluded that there are many unsolved questions, e.g. how gelatinous predation of zooplankton may reinforce anoxia and further habitat degradation in eutrophicated waters.
Keywords: Mnemiopsis leidyi; Beroe gracilis; Pleurobrachia pileus; Aurelia aurita; Predationimpact; Hydrography; Zooplankton; Limfjorden; Oxygen depletion; Hypoxia
Marine areas experiencing jellyfish and ctenophore blooms seem often to be those that reveal the greatest environmental degradation [1,2]. Limfjorden (Denmark) is heavily eutrophicated and large areas suffer from oxygen depletion each summer which has apparently caused a change “from fish to jellyfish” [3]. The abundance and predation impact of jellyfish and ctenophores in Limfjorden have been described in a number of studies conducted since 2003 [3-10] and recently reviewed by Riisgård and Goldstein [11] who also supplemented with observations from 2012 and 2013.
The common jellyfish Aurelia aurita is often very abundant in Limfjorden and may exert a considerable predatory impact on zooplankton and fish larvae. However, the population dynamics of A. aurita are not only strongly influenced by the high-salinity water brought into Limfjorden from the North Sea, but also by competition with the invasive ctenophore Mnemiopsis leidyi that occurred in Limfjorden for the first time in extremely high numbers in 2007 [12]. It has been suggested that Limfjorden may function as an incubator for M. leidyi with the potential to further spread M. leidyi into the Kattegat and adjacent Danish waters [9]. A recent study by Riisgård and Goldstein [11] reported on two bloom events of ctenophores, Pleurobrachia pileus and M. leidyi, along with their predators (Beroe spp.) in Limfjorden in the autumns of 2012 and 2013, when the previously dominating A. aurita was absent. Further it was observed that B. ovata, which is M. leidyi’s native predator, had now occurred as a new introduced species in Limfjorden.
The aim of the present study was to record the seasonal occurrence patterns of jellyfish and ctenophores in Limfjorden during 5 cruises with the Danish Marine Home Guard in 2014 in order to obtain a better understanding of the biological and environmental factors that control their species composition, population densities and predation impact. Our present knowledge on the complex interplay between biological and environmental factors that control the jellyfish and ctenophore populations in Limfjorden is briefly reviewed and discussed, including the importance of recruitment, life cycles, and hydrographic conditions.
Study area
Limfjorden is a Danish shallow-water system with a mean water depth of about 4.5 m that connects the North Sea via Thyborøn Kanal in the west to the Kattegat in the east (Figure 1). A dominating westeasterly wind creates an eastward current that brings highly saline North Sea water into Limfjorden, which also receives freshwater inputs from the surrounding land area, and this results in a salinity gradient from west to east [13-17]. Limfjorden is heavily eutrophicated and large areas suffer from oxygen depletion each summer [7,13,18], and this has apparently caused the bottom-dwelling fish to disappear and the number of jellyfish and ctenophores to increase [3]. Every year, strong westerly-winds induce about 60 intrusions of North Sea water through Thyborøn Kanal into Limfjorden succeeded by backflow of water into the North Sea on calm days. Annually, about 70 km3 of North Sea water enter Limfjorden, and approximately 65% flows eastwards into Løgstør Bredning which additionally receives 3 km3 freshwater so that about 8.7 km3 runs eastwards into Kattegat [10]. Inflowing water to Limfjorden mainly comes from the Jutland Coastal Current which flows northwards along the Danish west coast carrying mixed water masses from the English Channel and the southern North Sea [10,19-21].
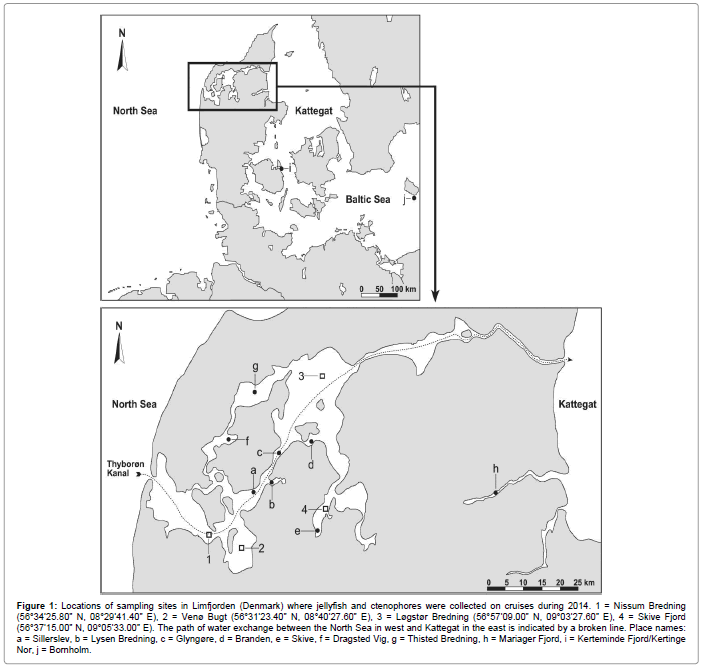
Figure 1: Locations of sampling sites in Limfjorden (Denmark) where jellyfish and ctenophores were collected on cruises during 2014. 1 = Nissum Bredning (56°34’25.80” N, 08°29’41.40” E), 2 = Venø Bugt (56°31’23.40” N, 08°40’27.60” E), 3 = Løgstør Bredning (56°57’09.00” N, 09°03’27.60” E), 4 = Skive Fjord (56°37’15.00” N, 09°05’33.00” E). The path of water exchange between the North Sea in west and Kattegat in the east is indicated by a broken line. Place names: a = Sillerslev, b = Lysen Bredning, c = Glyngøre, d = Branden, e = Skive, f = Dragsted Vig, g = Thisted Bredning, h = Mariager Fjord, i = Kerteminde Fjord/Kertinge Nor, j = Bornholm.
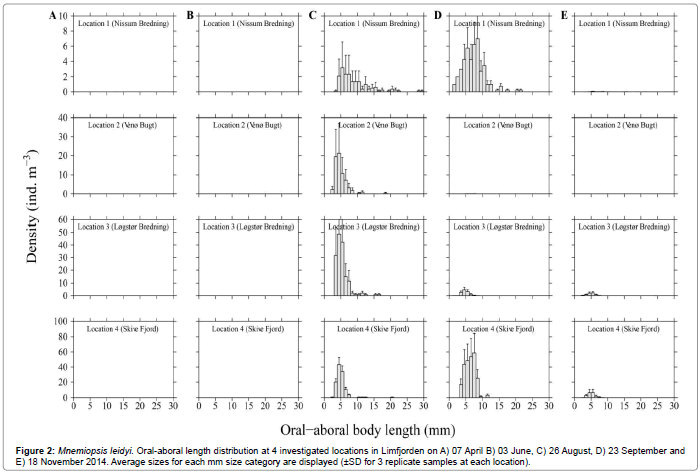
Figure 2: Mnemiopsis leidyi. Oral-aboral length distribution at 4 investigated locations in Limfjorden on A) 07 April B) 03 June, C) 26 August, D) 23 September and E) 18 November 2014. Average sizes for each mm size category are displayed (±SD for 3 replicate samples at each location).
Jellyfish and ctenophores
Jellyfish and ctenophores were collected with a 2 mm-meshed plankton net (mouth-ring diameter=1.5 m, mouth area=1.77 m2) at 4 locations in Limfjorden (Figure 1) during 5 one-day cruises in 2014 with the Danish Marine Home Guard MVH 902 ‘Manø’. Three hauls per location (haul length ~300 m) were performed at a depth of approximately 1 m at a speed of about 1.5 knots. After the three hauls, the number of medusae and ctenophores in the cod end of the plankton net were counted from (sub-)samples, and mean population densities were estimated from the densities in the 3 hauls. Further, mean (±SD) sizes were determined to the nearest mm after measuring ctenophore oral-aboral lengths (L, i.e. distance between mouth and opposite pole) and medusa inter-rhopalia umbrella diameters (d) of all, or at least 50 individuals per haul. Population clearance rates and predation impact of two species of ctenophores (Mnemiopsis leidyi, Pleurobrachia pileus) and the moon jellyfish (Aurelia aurita) on zooplankton (using copepods as reference) were estimated from recorded population densities using the equations below.
Mnemiopsis leidyi. The following relationship was used for converting oral-aboral length (L, mm) to individual body volume (V, ml ind.-1) (Riisgård et al.) [12]:
V = 0.0226L1.72 (1).
The following equation Riisgård et al., based on Decker et al. [10,23]. was used to estimate the individual clearance rate (Clind, l d-1) of M. leidyi feeding on copepods as a function of their body volume (V, ml):
Clind = 2.64V (2)
Pleurobrachia pileus. Clearance rates (Clind, l d-1) were estimated from the polar length (L, mm) by using the following equation for ctenophores feeding on copepods (Møller et al.) [23]:
Clind = 0.2L1.9 (3)
Aurelia aurita. The individual clearance rate (Clind, l d-1) of moon jellyfish feeding on copepods was estimated from the mean interrhopalia diameter (d, mm) by use of the equation [6]:
Clind = 0.0073d2.1 (4)
The volume-specific population clearance rates of jellyfish and ctenophores (Clpop, m3 water filtered by the ctenophore population in one m3 water per day=m3 m-3 d-1) was estimated as the product of the individual clearance rate (Clind, l d-1) and the population density (D, ind. m-3) for each location [10]:
Clpop = Clind × D/1000 (5)
The time (t1/2, d) it takes for a population of jellyfish or ctenophores with known Clpop to reduce the concentration of prey organisms (copepods) in V=1 m3 of water by 50% (i.e. the half-life of prey) was estimated as (Riisgård et al.) [10]:
t1/2 = ln 2 / Clpop (6)
Water chemistry and zooplankton
Water chemistry data (salinity, temperature, dissolved oxygen, chlorophyll a) and zooplankton biomass from Løgstør Bredning and Skive Fjord for 2014 were obtained from the national monitoring program of water and nature (NOVANA) conducted by the Danish Nature Agency, Ministry of the Environment. All data were collected and analyzed according to official monitoring guidelines [24]. Water chemistry data were obtained from CTD profiles from the surface to the bottom at the locations. Chlorophyll a concentrations at 1 m depth were obtained from the absorbance in spectrophotometric analysis of chlorophyll extracted with ethanol from the algae present in filtered water samples. Zooplankton was collected by using a submersible pump with a minimum capacity of 150 l min-1. The water was pumped up vertically by hauling the pump up at a speed of 0.5 m s-1. Collection of the zooplankton was done by using a 60 μm net at the outlet. The filtered water volume was calculated by using a mechanical flow meter. The filter was rinsed thoroughly and the sample preserved in either 2 to 3% formalin (pH 8.0 to 8.2) or neutral Lugol’s solution. The concentrations (ind. m-3) and biomasses (μg C l-1) of zooplankton species and taxonomic groups were subsequently determined.
Jellyfish and ctenophores
Species composition, population densities and size of jellyfish and ctenophores recorded during 5 cruises in Limfjorden in 2014 are listed in Table 1, along with the estimated half-life of zooplankton for the 3 most abundant species. It is seen that only Mnemiopsis leidyi exerted a notable predation impact, most pronounced in Løgstør Bredning and Skive Fjord in August, when the estimated half-life of zooplankton (copepods) were 4.8 and 7.3 d, respectively, and in Skive Fjord in late September when the half-life was 2.2 d. The size distribution of M. leidyi at the 4 investigated locations is shown in Figure 2. M. leidyi was not observed in Limfjorden during the first 2 cruises, 7 April and 3 June, respectively. But on the 3rd cruise (26 August) the ctenophore was found on all 4 locations, with largest body size (L=8.6 ± 4.8 mm) and lowest population density (D=18.5 ± 19.4 ind. m-3) in Nissum Bredning in the western part of Limfjorden. The highest density and smallest size were observed in Løgstør Bredning (D=158.8 ± 110.7 ind. m-3, L=4.9 ± 2.1 mm) and Skive Fjord (D=116.0 ± 25.6 ind. m-3, L=4.6 ± 1.9 mm). This pattern was somewhat changed one month later on the 4th cruise (23 September), and on the 5th cruise (18 November), when the population density of M. leidyi on all locations was strongly reduced. Along with the occurrence of M. leidyi, the small predatory ctenophore, Beroe gracilis was recorded on most sampling locations during the 3rd and 4th cruise, but not on the 5th cruise when only one large (L=64 mm) Beroe sp. individual was caught (Table 1). From Figure 3 it appears that B. gracilis eats small M. leidyi, but their low number suggests that the population-predation impact B. gracilis exerted on M. leidyi during 2014 was negligible.
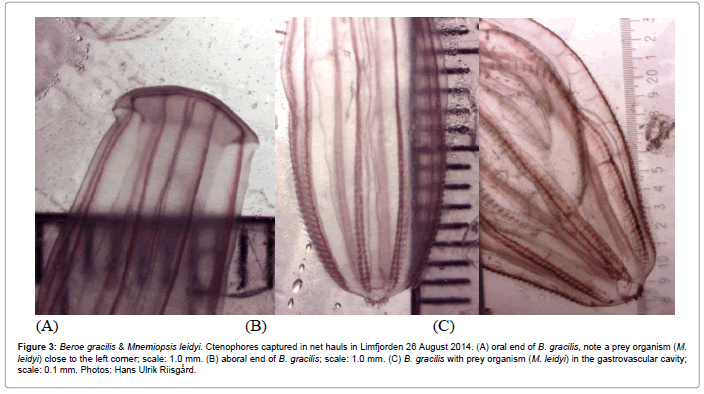
Figure 3: Beroe gracilis & Mnemiopsis leidyi. Ctenophores captured in net hauls in Limfjorden 26 August 2014. (A) oral end of B. gracilis, note a prey organism (M. leidyi) close to the left corner; scale: 1.0 mm. (B) aboral end of B. gracilis; scale: 1.0 mm. (C) B. gracilis with prey organism (M. leidyi) in the gastrovascular cavity; scale: 0.1 mm. Photos: Hans Ulrik Riisgård.
Water chemistry and zooplankton
Figures 4 and 5 show the temperature, salinity, dissolved oxygen and chlorophyll a concentrations in Løgstør Bredning and Skive Fjord during 2014. Figure 6 shows vertical profiles of temperature and salinity on selected dates, with full vertical mixing (Figure 6A) and bottom- near intrusion of high-saline North Sea water (Figure 6B) in Løgstør Bredning, and strong stratification due to combined thermo- and haloclines in Skive Fjord (Figure 6C), respectively. The dissolved oxygen concentration in the near-bottom water decreased to zero several times at both locations between June and September. As indicated by high chl a concentrations at the two locations (up to 45 μg l-1 in mid-August in Skive Fjord, Figure 5) the severe oxygen depletion had resulted in the release of nutrients (ammonium, phosphate; cf. Møller and Riisgård 2007c) [7] from the anoxic sediment causing an algal bloom which was subsequently followed by a tremendous bloom of the heterotrophic dinoflagellate Noctiluca scintillans (Figure 7A). This was again followed by a pronounced peak in the abundance of copepods and their offspring nauplii (Figure 7B). To judge from the population density of Mnemiopsis leidyi and the estimated half-life of copepods (t1/2=4.8 d) in Løgstør Bredning on 26 August 2014 (Table 1). It seems reasonable to suggest that the rapid decrease in the biomass of copepods during late July, and the following absence of zooplankton during the rest of the year was caused by a boom of the invasive ctenophore. As a consequence, it may therefore be suggested that the decrease in population density and the persistent small size of M. leidyi during September and November (Table 1) reflect a rapid depletion of prey organisms resulting in starvation and decay.
Species composition, population densities and predation impact
The abundance of ctenophores and jellyfish in Limfjorden are characterized by large fluctuations in species composition and population size, and while the yearly development of Aurelia aurita populations depends on local benthic polyps, incoming water from the North Sea seems to be responsible for a yearly reinvasion by Mnemiopsis leidyi and other ctenophore species from North Sea, and the present study supports the hypothesis that the North Sea serves as a refuge from where M. leidyi reinvades Limfjorden [10,11].
No or only very few ctenophores (Pleurobrachia pileus) and jellyfish (Aurelia aurita, Cyanea lamarckii) were recorded during the first two cruises in April and June 2014, and Mnemiopsis leidyi was first recorded on the 3rd cruise in late August (Table 1). However, the present first author (HUR) observed M. leidyi at a number of sites in Limfjorden (Sillerslev, Lysen Bredning, Glyngøre, Branden, Figure 1) between 25 and 28 June 2014. Therefore, M. leidyi must have entered Limfjorden with North Sea water through Thyborøn Kanal between 7 April and 25 July. In the period between the 2nd and 3rd cruise, salinity increased in the bottom-near water (from 25.5 to 27.9 psu) in Løgstør Bredning during the period 17 June to 1 July (Figures 5B and 7B). This is also coincident with the observation of the North Sea scyphomedusa Rhizostoma octopus which is frequently brought into Limfjorden in the autumn months [25]. The Jutland Coastal Current which carries mixed water masses from the English Channel and the southern North Sea gives rise to seawater inflows into Limfjorden, which also explains the occasional re-introduction of the hydromedusa Aequorea vitrina in Limfjorden where it is not able to establish a surviving population [6,7].
Differences in the life-history strategies of jellyfish and ctenophores are important for understanding the often strong variations in species composition and population dynamics. The scyphozoan Aurelia aurita has a life cycle which includes a pelagic medusa and a benthic polyp stage. Medusae reproduce sexually and larval development is followed by the disappearance of medusae in late autumn. After settlement, planula larvae metamorphose into polyps to pass the winter. In the early spring, the polyps produce ephyrae that are released into the water column to develop into a new generation of medusae [26,27]. In contrast to this, the ctenophore Mnemiopsis leidyi relies on a holoplanktonic life cycle. M. leidyi is a self-fertilizing hermaphrodite, releasing eggs and sperm in the ambient water where fertilization occurs [28-30]. Thus, high fecundity and rapid generation times during the whole season may explain the ability of M. leidyi (and Pleurobrachia pileus) to occur in large numbers in Limfjorden [11]. However, unless a major fraction of A. aurita ephyrae are washed out of Limfjorden in the early spring, this may also result in a mass occurrence [7] and interspecific competition with M. leidyi [9].The conspicuously few A. aurita observed in the present study as well as in the two preceding years [11] indicates that moon jellyfish populations in Limfjorden may currently experience the process of being outcompeted by the more numerous and faster reproducing M. leidyi, which could, on the longterm, also lead to a disappearance of local A. aurita polyps. However, alternative explanations are also possible, but especially improved understanding of polyp ecology seems necessary for understanding fluctuations in jellyfish numbers [26,31-34].
The abundance of ctenophores and jellyfish in Limfjorden are characterized by often large fluctuations in species composition and population size [11]. While the yearly development of Aurelia aurita populations in Limfjorden may largely depend on recruitment via asexual reproduction by local polyps, the incoming water from the North Sea is of decisive importance for the yearly reinvasion by Mnemiopsis leidyi and other ctenophore species from the warmer south western North Sea [35-37]. The present study supports the hypothesis that the North Sea in cold winters serves as a refuge from where M. leidyi reinvades Limfjorden, which may function as an incubator for M. leidyi, subsequently dispersing into the Kattegat and adjacent waters [10,11]. A number of observations support this hypothesis; thus, first observations of M. leidyi outside Limfjorden in 2014 were in Mariager Fjord (8 October, own observation), Kerteminde Fjord (3 October 2014, own observation), Kertinge Nor (7 November 2014, own observation) and the western Baltic Sea near Bornholm (21 November, Marie Stoor-Paulsen). The recent observations of B. ovata being a new species in Danish waters (Shiganova et al., Riisgård and Goldstein) has not yet caused measurable changes in the number and distribution of M. leidyi, and the biological and environmental factors controlling its sporadic occurrence are still poorly understood.
Interplay between biological and environmental factors: our present understanding
Recently, Riisgård et al. [3] gave an overview of the environmental status of the heavily eutrophicated Limfjorden, including the historical development of nutrient overloading and subsequent oxygen depletion in near-bottom water, and further how the bottom dwelling fish species decreased while the number of jellyfish and ctenophores increased. In a case study of Skive Fjord, Møller and Riisgård [7] demonstrated the links between primary production, oxygen deficiency, nutrients and jellyfish. Each summer, Skive Fjord suffers from oxygen depletion in the near-bottom water and this causes large amounts of nutrients (phosphate and ammonia) to be released from the anoxic sediment, which subsequently stimulates a phytoplankton bloom, followed by an increase in zooplankton biomass [7] (Figure 2). The same phenomenon was observed in the present study, not only in Skive Fjord but also in Løgstør Bredning, where the chlorophyll a concentrations became very high during periods with oxygen depletion (Figures 4 and 5) followed by a bloom of Noctiluca scintillans succeeded by an increase in copepod biomass (Figure 7). Møller and Riisgård [7] combined available data on jellyfish with data on oxygen, nutrients, chlorophyll a, zooplankton, and other data from various studies conducted in Skive Fjord during the period 1996 to 2005 and found that especially severe cases of oxygen depletion apparently take place in years with mass occurrence of jellyfish. Due to their high predatory impact on the zooplankton community, blooming algae are not efficiently grazed on, but settle to the bottom and decay, thus leading to more severe hypoxia.
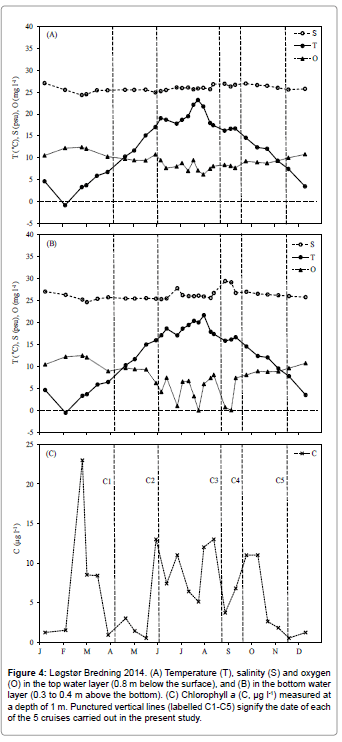
Figure 4: Løgstør Bredning 2014. (A) Temperature (T), salinity (S) and oxygen (O) in the top water layer (0.8 m below the surface), and (B) in the bottom water layer (0.3 to 0.4 m above the bottom). (C) Chlorophyll a (C, µg l-1) measured at a depth of 1 m. Punctured vertical lines (labelled C1-C5) signify the date of each of the 5 cruises carried out in the present study.
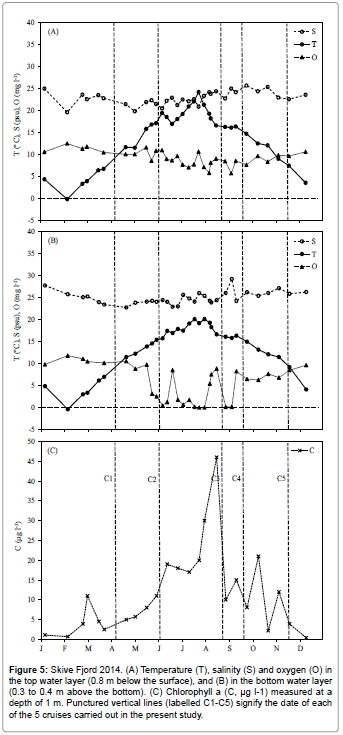
Figure 5: Skive Fjord 2014. (A) Temperature (T), salinity (S) and oxygen (O) in the top water layer (0.8 m below the surface), and (B) in the bottom water layer (0.3 to 0.4 m above the bottom). (C) Chlorophyll a (C, µg l-1) measured at a depth of 1 m. Punctured vertical lines (labelled C1-C5) signify the date of each of the 5 cruises carried out in the present study.
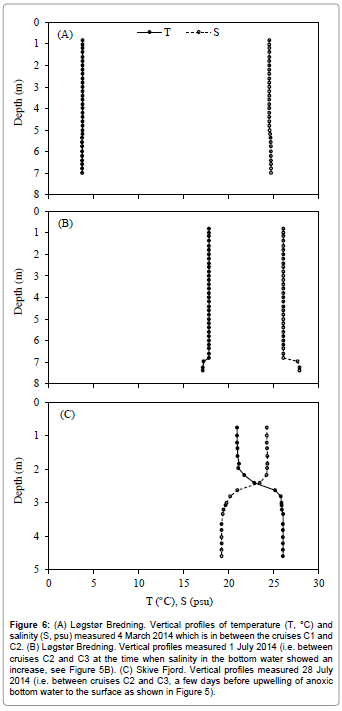
Figure 6: (A) Løgstør Bredning. Vertical profiles of temperature (T, °C) and salinity (S, psu) measured 4 March 2014 which is in between the cruises C1 and C2. (B) Løgstør Bredning. Vertical profiles measured 1 July 2014 (i.e. between cruises C2 and C3 at the time when salinity in the bottom water showed an increase, see Figure 5B). (C) Skive Fjord. Vertical profiles measured 28 July 2014 (i.e. between cruises C2 and C3, a few days before upwelling of anoxic bottom water to the surface as shown in Figure 5).
Mass occurrence of Mnemiopsis leidyi was first observed in Limfjorden in 2007, but it probably arrived the previous year [38]. This invasive holoplanktonic ctenophore exerted further predation pressure in addition to the indigenous scyphomedusa Aurelia aurita, which frequently caused the zooplankton organisms to become virtually absent, as also observed with some time-lag after the arrival of M. leidyi (Figure 7 and Table 1) in the present study. Figure 8 shows the oxygen concentration in Skive Fjord in the period 1982 to 2014. It is notable that the tendency to severe oxygen depletion in the bottomnear water during the summer months has apparently increased since 2006, and this may perhaps be linked with the arrival of M. leidyi in 2006 (in the absence of a plausible alternative explanation). Tomczak et al. [39] made an integrated ecosystem assessment for Limfjorden over the period from 1984 to 2008 and showed that from 1990 to 1995, the structure shifted from dominance by demersal fish species to that of pelagic fish species (sprat, herring, sticklebacks) and jellyfish. Nutrients (N, P) have been and are still considered a major bottom-up forcing factor in the system. Since the early 1900s and up to the mid-1980s there was a six-fold increase in nutrients which caused recurrent events of oxygen depletion, but although the loadings have decreased by 33% (N) and 67% (P) due to a water action plan decided by the Danish Parliament in 1987 this has so far not resulted in any noticeable improvement in ecosystem state [39,40]. In Skive Fjord, the tendency towards severely reduced oxygen concentrations during the summer has even increased since 2006 (Figure 8), possibly caused by the new invasive M. leidyi that eliminates the zooplankton so that the abundant microalgae are not grazed but settle and decay at the bottom, thus reinforcing severe hypoxia or anoxia and further habitat degradation. However, this hypothesis needs to be further substantiated.
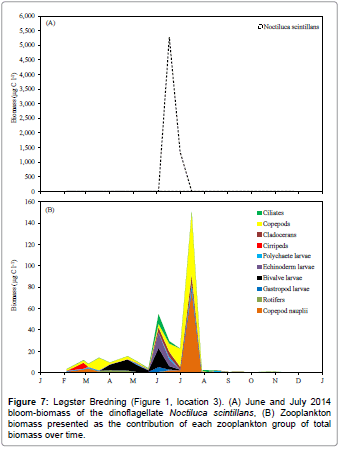
Figure 7: Løgstør Bredning (Figure 1, location 3). (A) June and July 2014 bloom-biomass of the dinoflagellate Noctiluca scintillans, (B) Zooplankton biomass presented as the contribution of each zooplankton group of total biomass over time.
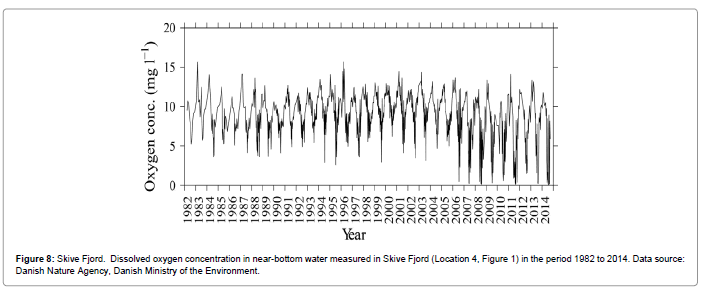
Figure 8: Skive Fjord. Dissolved oxygen concentration in near-bottom water measured in Skive Fjord (Location 4, Figure 1) in the period 1982 to 2014. Data source: Danish Nature Agency, Danish Ministry of the Environment.
According to Hansen et al. [41], the first severe case of oxygen depletion in Limfjorden in 2014 took place in mid-June, including the stratified Skive Fjord (Figure 6C) and other inner and central parts and central north-western parts (Thisted Bredning, Dragstrup Vig, Figure 1) where dead fish were observed on the beaches. By the end of June strong westerly winds resulted in mixing of the water column and improved oxygen conditions, but from the beginning of July the water column became stratified resulting in further oxygen depletion soon after. Thus, by the end of July 2014, about 30% of the total bottom waters in Limfjorden were depleted of oxygen with subsequent release of toxic hydrogen sulphide from the anoxic sediment (cf. Jørgensen) [13]. In Skive Fjord, off-shore winds in late July and early August pushed the surface water away from the shore, which caused upwelling of nearbottom water containing high concentrations of hydrogen sulphide. Subsequent oxidation made the new surface water white-colored due to precipitation of free sulphur [41]. No systematic studies of the effects of oxygen-depletion on fish, phyto- and zoobenthos have been conducted by the environmental authorities in 2014, and likewise possible effects on jellyfish and ctenophores remain unknown. However, due to the presence of high population densities of Mnemiopsis leidyi in Skive Fjord in late August and September (Table 1), it may be suggested that this ctenophore is rather resistant to poor environmental conditions as also emphasized by Decker et al. [22] and Kolesar et al. [23] who observed that M. leidyi is more tolerant to hypoxia than their prey and competing zooplanktivorous fishes.
From both the present study and earlier accounts Riisgård and Goldstein [11] it appears that the species composition of jellyfish and ctenophores, their population dynamics and predation impact in Limfjorden is highly unpredictable, depending on the complex interplay between recruitment, life cycles, and hydrographic conditions. Every year since 2007, M. leidyi has re-invaded Limfjorden from the North Sea via Thyborøn Kanal with water partly originating from the English Channel Antajan et al. [25], the Belgian part of the North Sea [42], and the western Dutch Wadden Sea where it is present the whole year round (van Walraven et al.) [36], and recent modelling by van der Molen et al. [43] and David et al. [37] supports this interpretation. M. leidyi has frequently exerted a significant predation impact on the zooplankton biomass in Limfjorden during late summer, but in October and November 2012 it was another ctenophore, Pleurobrachia pileus, that controlled the zooplankton in Løgstør Bredning, with t1/2=1.5 and 2.8 d in October and November, respectively, while only very few M. leidyi were observed [11]. However in most years since 2007, M. leidyi has thrived extremely well during late summers in Limfjorden, from where the ctenophore with its rapid reproduction may further spread into Kattegat and the inner Danish waters, perhaps even to the western Baltic Sea [44].
In the present study, it was observed that Beroe gracilis feed on small (<20 mm) Mnemiopsis leidyi (Figure 3) which confirms recent observations by Hosia et al. [27] and van Walraven et al. [36]. Apparently, B. gracilis follows M. leidyi from the southern North Sea from where also Pleurobrachia pileus comes, but hitherto, B. gracilis has been considered a specialized predator on P. pileus and new to the inner Danish waters [45]. This phenomenon may be another consequence of the now established self-sustaining populations of M. leidyi in the English Channel and southern North Sea coasts.
Herring (Clupea harengus) and sprat (Sprattus sprattus) are pelagic fish species in Limfjorden and have to date been used for both human consumption and processing for fish meal and oil, but because these fish feed on zooplankton, they may, in certain areas and periods, compete with large numbers of ctenophores and/or jellyfish [11]. The herrings enter Limfjorden from both the North Sea and the Kattegat in late autumn and early winter to spawn in the spring, and the adults leave again in early summer while the juveniles stay (Erik Hoffmann, pers. comm.), but no studies have so far attempted to determine the degree of interspecific competition for zooplankton [46]. The short estimated half-lives of zooplankton caused by M. leidyi in August 2014 in Løgstør Bredning and Skive Fjord (Table 1) and the conspicuous reduction of the zooplankton biomass (Figure 7) suggests very poor feeding conditions for juvenile herrings in Limfjorden [47,48].
The present study has been focused on the complex interplay between biological and environmental factors that control the jellyfish and ctenophore populations in Limfjorden, and hopefully, the many still unsolved questions may stimulate future research with focus on e.g. how gelatinous predation of zooplankton may reinforce anoxia and further habitat degradation in eutrophicated waters.
Thanks to the Danish Nature Agency, Ministry of the Environment, for providing water chemical data, to skippers Mogens Grimstrup and Henning Bach and their crew aboard MHV 902 MANØ (Danish Marine Home Guard), to Jan Brandt Wiersma for technical assistance, to Bernd Lüskow for help with the map, and to Jens Würgler Hansen and Ole Secher Tendal for comments on the manuscript. Two researchers, Sabine Holst and Cornelia Jaspers took part in the 3rd and 4th cruise, respectively, and they assisted in counting and measurement of collected jellies.