Pancreatic Disorders & Therapy
Open Access
ISSN: 2165-7092
ISSN: 2165-7092
Research Article - (2015) Volume 5, Issue 2
As an endogenous oscillator with a period of about 24 h, the circadian clock system enables us to optimize energy acquisition and homeostasis. In mammals, the clock system comprises of a central pacemaker and peripheral clocks. The pancreas has been shown to be a peripheral oscillator, which suggests a direct relationship between the circadian clock and pancreatic functions. Supported by evidences from animal models with molecular manipulation of clock genes and genetic studies in humans, the pivotal role that impaired clock system plays in the process of both endocrine pancreatic disorders and exocrine pancreatic disorders has been discovered. These findings provide novel insights into the pathogenesis of pancreatic disease, as well as possible new medical technologies.
<Keywords: Pancreatic disorders, Mammalian clock machinery, Circadian system
Since the modern era of clock genetics, or called “chronobiology” began in the 1970s, when the clock gene period (PER) was first described in Drosophila melanogaster [1], the emerging image of circadian clock system has triggered a great upsurge within research fields in the past few decades. As a fundamental characteristic of all living things and their organelles, the 24-h timing system closely correlates with nearly all aspects of biological processes and behaviors such as feeding-fasting cycles, sleep-wake cycles, metabolic cycles, and fluctuations of body temperature and blood pressure [2-5]. Accumulated experimental and epidemiological evidences suggest a strong relationship between circadian disruption and human health problems including disorders within the gastrointestinal tract and digestive glands [6,7]. Liver is a well-documented peripheral oscillator. It was reported that altered clock system plays a pivotal role in the pathogenesis of alcoholic liver disease [8], liver fibrosis [9,10], nonalcoholic fatty liver disease [11], and hepatic carcinoma [12]. Given that the liver and pancreas have similar embryological origins, it is also plausible for the participation of clock system in the course of pancreatic disorders [13]. Here, this review addresses the link currently known between the pancreas and the clock system, based on the conclusions of peer researchers and our latest findings, to provide novel insights into the pathogenesis and prognosis of pancreatic disorders.
The clock system of mammals is strictly hierarchical -the central oscillator synchronizes oscillations of peripheral clocks [14-16]. Located in the suprachiasmatic nuclei (SCN) of the anterior hypothalamus, the central pacemaker is oriented towards the external environment through the reception of light. The master clock is cell autonomous and molecularly comprises at least three interlocking feedback loops [17]. Of the most importance, the heterodimeric partnership of BMAL1 (also known as ARNTL) and CLOCK, that bind E-box sites and induce the expression of the repressors PER and cryptochrome (CRY). The latter two then translocate back into the nucleus and inhibit their own expression by interfering with the BMAL1: CLOCK complex, as shown in Figure 1. Other two loops consist of nuclear receptors (ROR and REV-ERB) or albumin D-box-binding protein (DBP and NFIL3). These transcription factors within each of these loops can regulate clock-controlled genes (CCGs), which constitute about 10% of the expressed genes in a given tissue (SCN or in peripheral tissues), and serve their different biological functions[3,18-20].
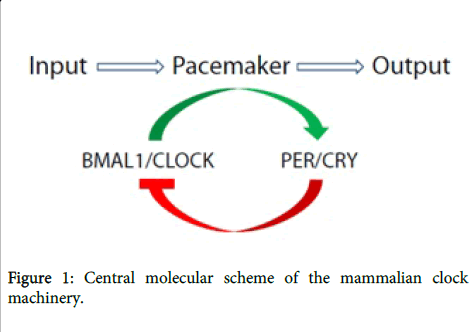
Figure 1: Central molecular scheme of the mammalian clock machinery.
In addition to the central clock in the brain, peripheral clocks with similar molecular components exist in several organs such as liver, heart, kidney, adipose tissue, and pancreas [21,22]. SCN maintains the phase alignment of the peripheral clocks, and optimizes metabolism and energy homeostasis throughout the day. Although not entirely understood, the hypothalamus pituitary adrenal axis and the autonomic nervous system, as well as glucocorticoids and catecholamines released by them respectively, serve as the key link through which the SCN clock maintains harmony with peripheral clocks [23].
Progress in understanding the relationship between the circadian system and pancreas, and the mechanisms by which the former influences the latter is lagged, even though a circadian oscillation throughout a 24 h period of clock genes including BMAL1, CLOCK, PER1, PER2, REV-ERBα and some others has been shown in rat pancreas, mouse pancreas, mouse islets, and human islets [24-26]. Recently, we obtained Period2::Luciferase (Per2::Luc) knockin mice on a C57Bl/6J background, which were housed in a 12 h light:12 h dark cycle, then pancreatic tissues were harvested and placed in a special dish, followed by continuous monitoring for >5 days. It was observed that pancreatic explants from Per2::Luc mouse showed a beautiful oscillation of the Per2 protein even in the ex vivo condition (Figure 2), which suggested that the pancreas possesses an intact clock.
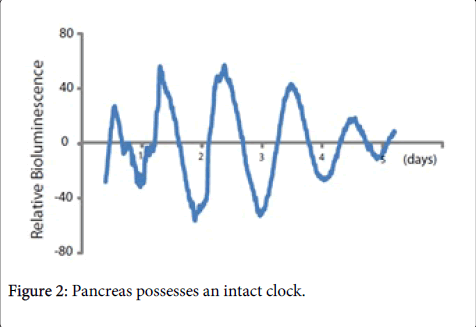
Figure 2: Pancreas possesses an intact clock.
By releasing hormones such as insulin and glucagon, the pancreas regulates the metabolism and energy homeostasis of human body, which, at the same time, has been demonstrated to be governed by the circadian system as well. Parameters related to glucose metabolism such as glucose tolerance, insulin sensitivity, as well as glucose, glucagon, and insulin plasma levels are known to exhibit 24 h circadian variations independent of food intake along the day [27-30]. Mounting reports have identified certain clock genes, for example, BMAL1, CLOCK, and REV-ERB, are involved in the growth, survival, and proliferation of insulin-secreting β cells and the glucagon-secreting α cells, so that play a pivotal role in the regulation of glucose levels [31-33]. These conclusions are supported by both animal models with manipulated clock genes and clinical investigation on large samples. In addition, the nearest finding by our group suggested that there is a possible link between clock genes and pancreatic stellate cell activation (results will be shown in our article ready for publication).
The relationship between alterations in circadian rhythm derived from current life style or social habits and metabolic diseases has been proved by several clinical investigations [34]. Disruption of the circadian rhythm correlates with pancreatic diseases including endocrine dysfunctions and exocrine dysfunctions. As a paradigmatic model of circadian misalignment, shift workers have alterations in pancreatic β-cell responses and in glucose and lipid metabolism, which places them at high risk of experiencing diabetes, obesity, cardiovascular disease, and other metabolic diseases [35-38]. Genetic studies in human have demonstrated that single nucleotide polymorphisms of CLOCK, BMAL1, CRY2, or PER2 closely correlate with total energy intake, obesity, impaired fasting glucose, and type 2 diabetes [39-42]. Using animal models with molecular manipulation of clock genes, more detailed interaction between clock genes and endocrine pancreas has been observed. Both CLOCK19/19mutant mice and pancreas-specific BMAL1knockout (KO) mice exhibit hyperglycemia, hypoinsulinemia, and glucose intolerance [43]. Aberrant insulin secretion was detected in isolated islets of whole-body BMAL1 KO and pancreas-specific BMAL1KO mice [44], while a similar diabetic phenotype and secretion defects were reported in β-cell-specific BMAL1 KO mice [33]. Other clock genes also participate in the pathophysiological process of pancreatic disorders. The findings by Vieira et al. identified REV-ERBα as an intracellular regulator of glucagon secretion via AMPK/Nampt/Sirt1pathway [31]. While Nakabayashi et al. found that downregulation of DBP contributed to direct suppression of the expression of BMAL1in diabetes [45]. These results unravel that the endocrine pancreas has an intrinsic self-sustained clock, and its disruption contributes to the endocrine disorders of the pancreas.
Recent findings have shown that the impaired circadian system is also involved in exocrine pancreatic disease. An important aspect to consider in pancreatic carcinogenesisis the role of the circadian clock. Tavano et al. [46] and Relles et al. [47] found a significant downregulation of BMAL1, CRY1/2, PER1/2/3, SIRT1, and some other key clock genes in pancreatic ductal adenocarcinoma specimens, respectively. So far, considerable alterations in sleep quality and other circadian abnormalities among patients with chronic pancreatitis have been witnessed by clinical workers. More than one-third of patients with chronic pancreatitis showed signs of desynchronosis, which was recorded by the investigation of Fedorov et al. on 417 patients with chronic pancreatitis [48]. They also reported that the use of given medical technologies in patients’ most favorable period makes it possible to increase the results of treatment from 11.4 to 18.3%. For further evidence, our group compared the expression of pancreas clock genes in control and fibrosis mice, which was established by 4-week cerulein induction. The findings of the present study provided direct evidence that the molecular clockwork is completely disrupted in fibrotic pancreas (figures and data will be shown in our article ready for publication). In addition, we found that such a disruption of circadian system correlates with an altered profile of cytokines secretion in the experimental mice, which suggested their lies a possible link between chronic pancreatitis and clocks. Jaworek et al. [49] found that experimental pancreatitis induced in night was significantly less severe than that was induced during the light phase, and they proposed that the way circadian network modulates severity of pancreatitis was through the rhythmic secretion of melatonin.
Tremendous progress in the past decades has been made in our understanding of what constitutes a circadian system and how it affects our health and disease. As an organ responsible for the secretion of various hormones and digestive enzymes, the pancreas possesses an intrinsic clock, and thus ties its physiology and pathophysiology to the regulation of clock system. Evidences from both bench and bedside have shown the role played by circadian rhythm and clock genes in the course of pancreatic disorders and diseases, which, to some extent, enhances the understanding about these pancreatic diseases that puzzled researchers for years. On the other hand, application of circadian rhythm-based intervention strategies to these disease targets is of great potential and will likely lead to inspiring advances in the future.