Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2015) Volume 4, Issue 5
One of the hallmarks of cancers is the silencing of tumor suppressor genes and certain signaling pathways. The Wnt pathway is activated in many types of cancers, leading to tumor progression and metastasis. Here we demonstrated that integrin-linked kinase (ILK) plays a critical role in the suppression of the Wnt pathway via decreasing Wnt3a-induced stabilization of β-catenin, which leaded to the block of the functional gap-junctional intercellular communications (GJIC). Inhibition of ILK in prostate cells resulted in the inactivation of the Wnt pathway components GSK3β and inactivation of Lef1 transcription with concomitant inducing of connexins, which forming channels between adjacent cells. In line with the above changes, over-expression of connexin 43 (Cx43) also inhibited the activity of Lef1 transcription factor induced by Wnt3a media and the transcription of target genes of Wnt signaling. Together, our data demonstrates a role for ILK as a regulator of Wnt pathway through Cx43 which is a negative feedback regulation of Wnt signaling and ILK may be a potential cancer therapeutic target.
<Keywords: ILK; Wnt signaling; Connexin 43; Prostate tumor cells
ILK: Integrin-Linked Kinase; Cx43: Connexin 43; Gjs: Gap Junctions; GJIC: Intercellular Communication via Gap Junctions; Lef-1: Lymphoid Enhancer-Binding Factor-1; TCF: T Cell Factors
Tumor progression, metastatic potential and response to therapy depend on complex genetic, epigenetic and tumor micro environmental interplay. Secreted signaling molecules of the Wnt family have been widely investigated and found to have a prominent role to induce human malignant diseases, such as breast and prostate cancer [1-3]. In the presence of Wnt stimulation, β- catenin at Thr41, Ser37 and Ser31 sites are dephosphorylated [4], resulting in accumulation of unphosphorylated β-catenin in the cytoplasm. The stabilized β-catenin enters the nucleus, which consequently activates transcription of downstream target genes via lymphoid enhancer-binding factor-1 (Lef-1) and T cell factors (TCF) [5,6]. Integrin-linked kinase (ILK) is an integrin associated actin and tublin cytoskeletal interacting effector, which is involved in the regulation of cell survival, proliferation, and migration [7,8]. ILK coordinates several signaling pathways, and it has been shown to activate Pi3Kinase/Akt, Wnt, TGF-β and epithelialmesenchymal transition signaling in various types of cancer cells [9-11]. Moreover, siRNA-mediated silencing of ILK in MDA-MB-231, PC3, and other cell lines examined result in inhibition of Ser473-Akt phosphorylation and induction of apoptosis [12,13], and the smallmolecule inhibitors of ILK, QLT0267 and T315 [14,15] , exhibit antitumor efficacy in vitro and/or in vivo in various types of cancer cells, in part, by targeting Akt activation. To form an organized multicellular structure, the cells within tissues and organs are connected via specialized junctions [16]. The four types of junctions between cells are the adherent junctions, tight junctions, desmosomes and gap junctions (GJs) [17]. Gap junctions are membrane-spanning channels that allow for the movement of small molecules across cell membranes [18,19]. Intercellular communication via gap junctions (GJIC) plays important roles in regulating cell growth and differentiation and in maintaining tissue homeostasis [20,21]. This type of cell communication is often impaired during cancer development, and several members of the connexin protein family have been shown to act as tumor suppressors [16,22,23]. A gap junction may consist of up to several thousand intercellular channels. One channel is formed by two hemi channels called connexins, with each composed of six connexin protein subunits.The connexin protein family comprises 21 members in humans, of which the best studied isoform is connexin 43 (Cx43) [17,21]. One potential regulator of gap junction expression and function that might have important implications for developmental processes as well as for normal function in adult stages is the Wnt family genes. However, the effect of ILK on GJIC remains elusive. Hence in the present study, we tried to determine whether ILK regulate Cx43 expression and function in prostate tumor cells and the mechanism.
Cell culture, transfection and infection
All the cell lines used in the present study were obtained from ATCC (Manassas, VA). Wnt3a-expressing and control L cells were used for the production of biologically active Wnt3a conditional media (Wnt3a) and control L media (L), respectively. PC3 cells were maintained in DMEM/F12 1:1 with 10% bovine serum (Life Technologies) according to instructions, DU145 cells were maintained in the DMEM. Wnt3aand L- conditioned media were used at 1:1 dilution in normal growth media. Cells were harvested by trypsinization, re-suspended in culture media, and transfected with various siRNA or plasmids using Lipofectamine Plus (Invitrogen) according to the manufacturer’s instructions. Lentivirus expressing shRNA-ILK or Cx43 was produced as below, and diluted 1:1 with growth media before use. For viral infections, cells were incubated with the virus for 12 hours before switched to growth media.
Lentivirus packaging
To generate shRNA lentivirus, shRNA vectors were co-transfected into HEK293T cells with the packaging plasmids pMDL, pREV and pVSVG using Lipofectamine. Supernatants were collected 48 hrs after transfection, and passed through 0.45 μm nitrocellulose filters. PC3 cells were infected with viral supernatants diluted 1:1 with growth media and supplemented with 5 mg/ml polybrene. Lentivirus of overexpression pLVX vectors were co-transfected into HEK293T cells with the packaging plasmids pMD2G and pSPAX and collected the supernatants using the same method.
Luciferase assay
For Lef1 transcriptional activity assays, the indicated cells were plated on 24-well dishes in triplicates and co-transfected with 500 ng of a 10:1 mixture of Lef1 reporter and Renilla. After 24 hrs, the cells were cultured in the media in presence of either L or Wnt3a media for 48 hrs. In some cases, the cells were co-transfected with siRNA oligonucleotides. The luciferase activity was assayed using the dual luciferase reporter assay system (Promega).
Quantitative RT-PCR
PC3 cells were seeded in 6-well plates. At confluence, cells were infected with lentivirus and stimulated with L or Wnt3a media for 48 hrs. Total RNA was extracted using TRIzol reagent (Takara Biotechnology). Messenger RNA levels of ILK, Cx43, Cx32, c-MYC and CyclinD1 were determined by quantitative RT-PCR according to the manufacturer’s instructions.
Western blot analysis
Cellular lysates were prepared and proteins were resolved by sodium dodecylsulfate- polyacrylamide gel electrophoresis. Proteins were transferred to PVDF membranes. The blots were blocked with 3% bovine serum albumin for 1 h at room temperature and probed with rabbit anti-human antibodies against p-glycogen synthase kinase (GSK) 3β, GSK3β, ILK, GAPDH.
Immunoprecipitation
PC3 cells were plated 1 × 104 cells/cm2 overnight, after a variety of treatments. Cytosolic and nuclear fractions of cells were prepared by using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Scientific, Waltham, MA, USA) as per the manufacturer’s instructions. β-Actin and Lamin-B were used as the internal standards for the cytosolic and nuclear fractions, respectively.
Statistical analysis
With the exception of western blot analysis, graphical data are presented as mean values ± standard error (SD). P-values were calculated using unpaired Student’s t-tests comparing control with treated cells. Statistically significant differences are indicated within the text and by asterisks. Data are representative of at least three independent experiments.
Inhibition of ILK activity decreased Wnt3a-induced activation of Lef1 transcriptional activity
The Wnt signaling pathway is required during embryonic development and for the maintenance of homeostasis in adult tissues. However, aberrant activation of the pathway is implicated in a number of human disorders, including cancer of the gastrointestinal tract, breast, liver, melanoma, and hematologic malignancies. To determine a potential role of ILK in canonical Wnt pathway, we initially examined activity of the Lef1 transcription factor in different human cancer cell lines. As shown in (Figure1A and 1B) Lef1-luciferase activity induced by Wnt3a media was suppressed significantly upon siRNA-mediated knockdown of ILK expression in PC3 and DU145 prostate tumor cells, indicating that inhibition of ILK leads to Lef1 inactivation in these cell lines. In contrast, over-expression of ILK significantly induced the Lef1-luciferase activity by 2 fold (Figure 1C and 1D). These data suggest that inhibition of ILK negates canonical Wnt signaling, in line with this observation, over-expression of ILK induced canonical Wnt signaling in either the presence or absence of Wnt3a.
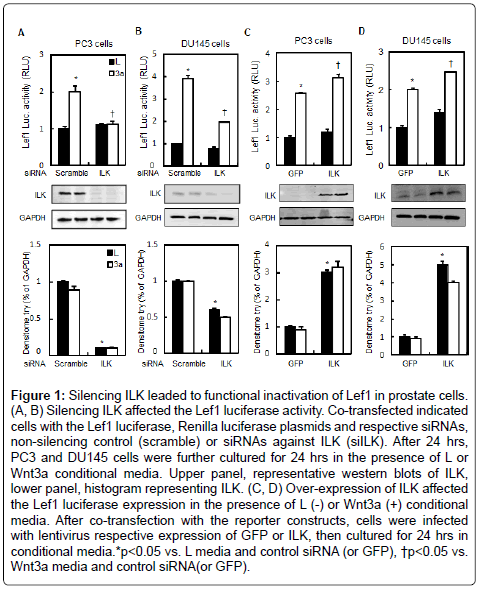
Figure 1: Silencing ILK leaded to functional inactivation of Lef1 in prostate cells. (A, B) Silencing ILK affected the Lef1 luciferase activity. Co-transfected indicated cells with the Lef1 luciferase, Renilla luciferase plasmids and respective siRNAs, non-silencing control (scramble) or siRNAs against ILK (siILK). After 24 hrs, PC3 and DU145 cells were further cultured for 24 hrs in the presence of L or Wnt3a conditional media. Upper panel, representative western blots of ILK, lower panel, histogram representing ILK. (C, D) Over-expression of ILK affected the Lef1 luciferase expression in the presence of L (-) or Wnt3a (+) conditional media. After co-transfection with the reporter constructs, cells were infected with lentivirus respective expression of GFP or ILK, then cultured for 24 hrs in conditional media.*p<0.05 vs. L media and control siRNA (or GFP), †p<0.05 vs. Wnt3a media and control siRNA(or GFP).
Inhibition of ILK activity decreased Wnt3a-induced stabilization of β- catenin
Given that β-catenin is the central signal transducer of the Wnt signaling pathway, we next examined the effect of the inhibition of ILK activity on β-catenin protein content, and Wnt3a-induced stabilization of β-catenin in prostate tumor cells. Wnt3a induced the β-catenin protein levels in cytosolic and nuclear fractions by 1.5 and 3.0 fold in PC3 cells, respectively. Whereas knockdown of ILK decreased nuclear fraction of β-catenin by 67% in the presence of Wnt3a and had no obvious effect on the basal levels of cytoplasmic β-catenin (Figure 2A). Silencing of ILK also reduced nuclear β-catenin protein levels in response to recombinant Wnt3a protein in DU145 cells (Figure 2B). β-catenin stabilization is considered as the key factor in the activation of the downstream components of Wnt signaling pathway. Phosphorylation of β-catenin at Ser45 by casein kinase-1 (CK1) and at Thr41, Ser37, and Ser33 by GSK3β regulates its stabilization in the cytosol, and the phosphorylated β-catenin is recognized by E3 ubiquitin ligase and undergoes proteolytic degradation. To determine the role of ILK in the stabilization of β-catenin, we assessed changes in β-catenin phosphorylation in PC3 cells. The levels of β-catenin phosphorylation at Ser33 sites in PC3 cells transfected with ILK or control (Scramble) siRNA were examined in either the presence or absence of Wnt3a media. Phosphorylation of β-catenin at Ser33 was reduced in response to Wnt3a (50%) but induced by siILK (Figure 3A). Parallel to the changes of phosphorylation levels of β- catenin, Wnt3a increased the GSK3β phosphorylation at Ser9 while siILK attenuated it.
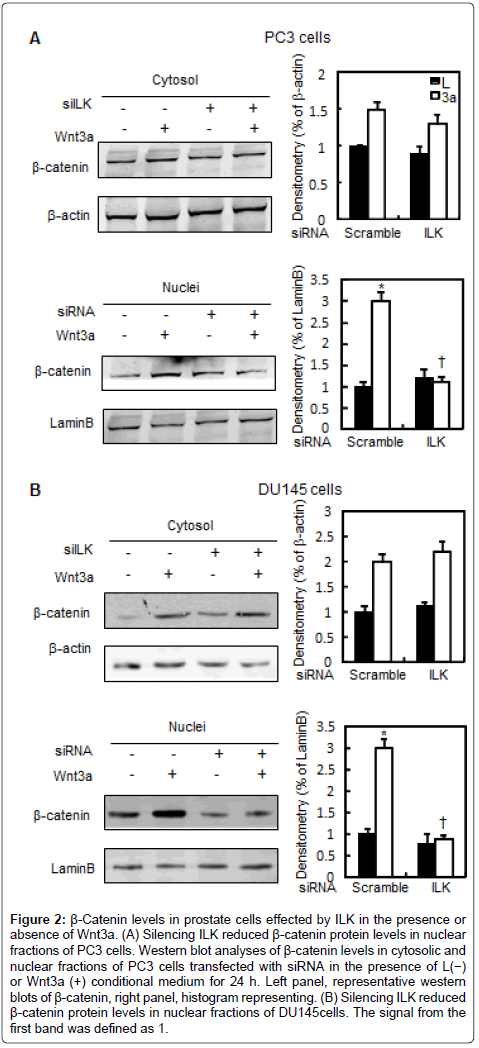
Figure 2: β-Catenin levels in prostate cells effected by ILK in the presence or absence of Wnt3a. (A) Silencing ILK reduced β-catenin protein levels in nuclear fractions of PC3 cells. Western blot analyses of β-catenin levels in cytosolic and nuclear fractions of PC3 cells transfected with siRNA in the presence of L(−) or Wnt3a (+) conditional medium for 24 h. Left panel, representative western blots of β-catenin, right panel, histogram representing. (B) Silencing ILK reduced β-catenin protein levels in nuclear fractions of DU145cells. The signal from the first band was defined as 1.
Then, we performed reporter assays and co-immunoprecipitation to confirm GSK3β signaling as a downstream event of ILK signals in the regulation of β-catenin stabilization. GSK3β inhibitor, SB216763, increased Lef1-luciferase activity but showed no significant effect in PC3 cells expressing ILK siRNA (Figure 3B). Furthermore, protein complexes precipitated with a ILK antibody contained an abundance of GSK-3β as well as ILK as expected, suggesting that physical interactions exist between endogenous ILK and GSK-3β (Figure 3C).
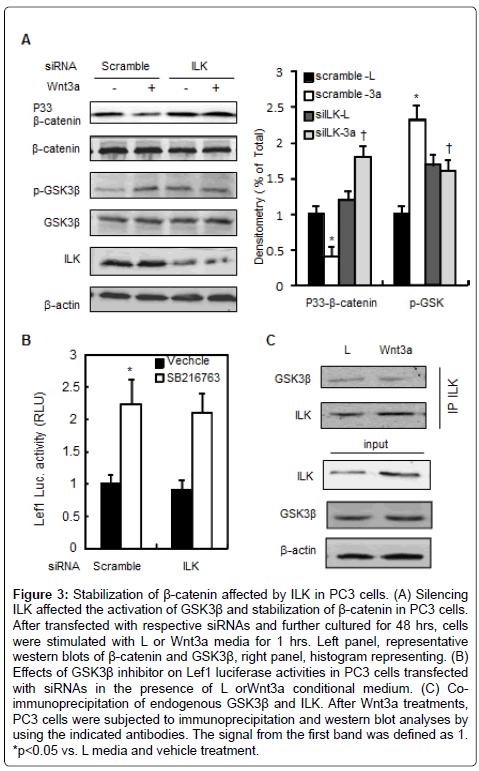
Figure 3: Stabilization of β-catenin affected by ILK in PC3 cells. (A) Silencing ILK affected the activation of GSK3β and stabilization of β-catenin in PC3 cells. After transfected with respective siRNAs and further cultured for 48 hrs, cells were stimulated with L or Wnt3a media for 1 hrs. Left panel, representative western blots of β-catenin and GSK3β, right panel, histogram representing. (B) Effects of GSK3β inhibitor on Lef1 luciferase activities in PC3 cells transfected with siRNAs in the presence of L orWnt3a conditional medium. (C) Coimmunoprecipitation of endogenous GSK3β and ILK. After Wnt3a treatments, PC3 cells were subjected to immunoprecipitation and western blot analyses by using the indicated antibodies. The signal from the first band was defined as 1. *p<0.05 vs. L media and vehicle treatment.
Taken together, these observations suggest that inhibition of ILK may suppress canonical Wnt signaling through inducing GSK3β- mediated phosphorylation of β-catenin at Thr41, Ser37, and Ser33 sites.
Effects of ILK inhibition on growth and migration of prostate cancer cells
Our above results have indicated that ILK is involved in Wnt/β- catenin signaling, to examine the role of ILK in growth of prostate cancer cells, we tested the expression of canonical Wnt signaling target genes in the presence or absence of Wnt3a in PC3 cells. Wnt3a not only increased mRNA levels of CyclinD1 and c-Myc (Figure 4B and 4C), but also increased the expression of connexin 43 (Cx43) and Cx32, encoded gap junctions (Figure 4D and 4E). These observations suggest that ILK may be associated with intercellular communication via gap junctions. Proliferation rate was examined to detect the effect of inactivation of Wnt pathway in prostate cells. As shown in Figure 5A, the growth curves were shown that the proliferation of the cells treated with Wnt3a media was increased compared to the cells treated with L media after 24 hrs of culture, whereas the proliferation of PC3 cells infected by ILK-shRNA-expressing (shILK) lentiviruses slowdown. To assess the role of ILK in cell migration, we performed the wound-healing assay. Scrape wounds were introduced into confluent monolayers of PC3 cells and wound closure rates monitored at 24 hourly intervals in the PC3 cells infected with shILK lentiviruses, whereas significant increase in closure rates occurred in PC3 cells infected with scrambleshRNA- expressing (scramble) lentiviruses (Figure 5B). Thus, inhibition of ILK reduced cell proliferation and migration in prostate cells, these may be associated with the change of intercellular communication via gap junctions.
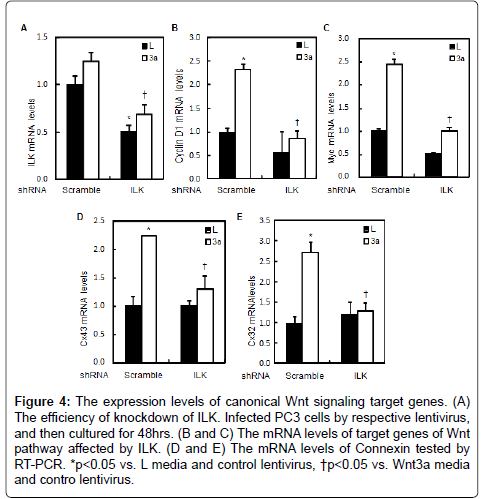
Figure 4: The expression levels of canonical Wnt signaling target genes. (A) The efficiency of knockdown of ILK. Infected PC3 cells by respective lentivirus, and then cultured for 48hrs. (B and C) The mRNA levels of target genes of Wnt pathway affected by ILK. (D and E) The mRNA levels of Connexin tested by RT-PCR. *p<0.05 vs. L media and control lentivirus, †p<0.05 vs. Wnt3a media and contro lentivirus.
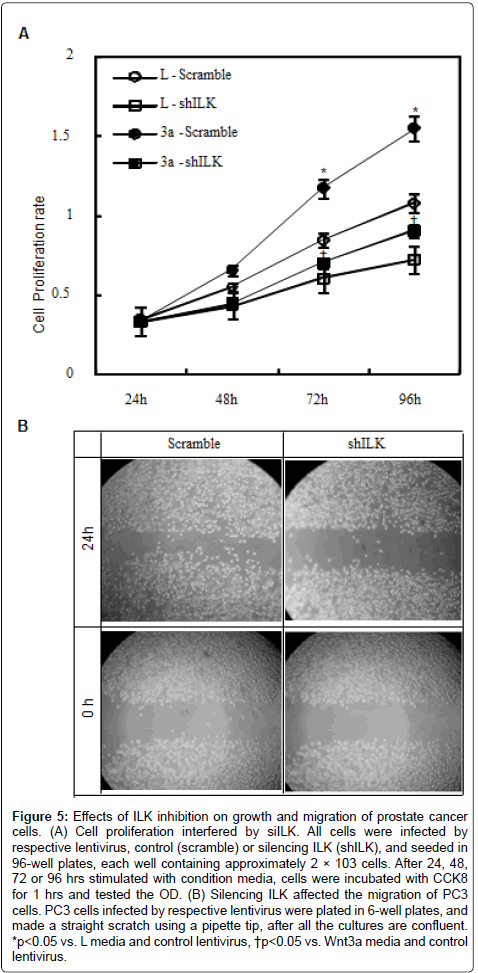
Figure 5: Effects of ILK inhibition on growth and migration of prostate cancer cells. (A) Cell proliferation interfered by siILK. All cells were infected by respective lentivirus, control (scramble) or silencing ILK (shILK), and seeded in 96-well plates, each well containing approximately 2 × 103 cells. After 24, 48, 72 or 96 hrs stimulated with condition media, cells were incubated with CCK8 for 1 hrs and tested the OD. (B) Silencing ILK affected the migration of PC3 cells. PC3 cells infected by respective lentivirus were plated in 6-well plates, and made a straight scratch using a pipette tip, after all the cultures are confluent. *p<0.05 vs. L media and control lentivirus, †p<0.05 vs. Wnt3a media and control lentivirus.
Cx43 constrain growth of prostate cancer cells by interfering with the Wnt/β-catenin pathway
Impairment of gap junctional intercellular communication, caused by mutations or loss of function of connexins, especially Cx43 which is expressed in several tissues and organs, like heart, gonads, lens and skin, is involved in a number of diseases. Our above results have indicated that ILK is involved in Wnt/β-catenin signaling and affects the mRNA levels of Cx43 in prostate cells. In order to examine the role of connexin in tumor progression, we tested the effect of Cx43 on proliferation. Wnt3a promoted PC3 cells proliferation slightly, whereas cells ectopically expression of Cx43 slowed down the proliferation compared with control (Figure 6A). The mRNA levels of CyclinD1, the target genes of Wnt/β-catenin signaling were induced by Wnt3a media but attenuated by over-expression of Cx43 (Figure 6B). Then we examined the apoptosis affected by Cx43 in PC3 cells. Western blotting results showed that over-expression of Cx43 induced the levels of caspase3 and caspase9 (Figure 6C). These observations raise the possibility that ILK affected the PC3 cells proliferation and vitality due to the abnormal expression of Cx43.
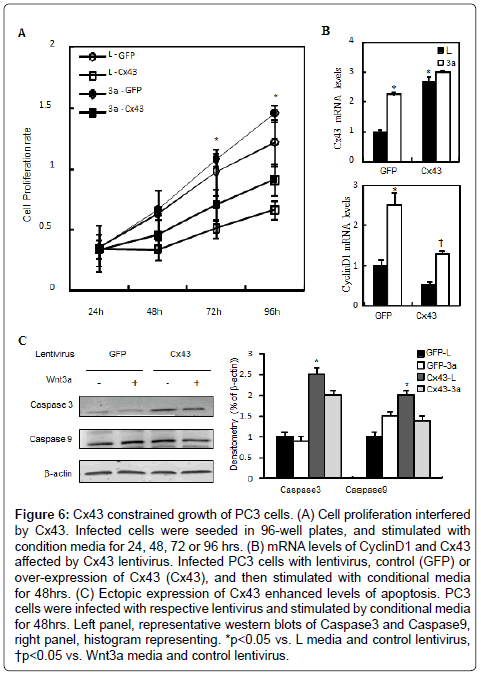
Figure 6: Cx43 constrained growth of PC3 cells. (A) Cell proliferation interfered by Cx43. Infected cells were seeded in 96-well plates, and stimulated with condition media for 24, 48, 72 or 96 hrs. (B) mRNA levels of CyclinD1 and Cx43 affected by Cx43 lentivirus. Infected PC3 cells with lentivirus, control (GFP) or over-expression of Cx43 (Cx43), and then stimulated with conditional media for 48hrs. (C) Ectopic expression of Cx43 enhanced levels of apoptosis. PC3 cells were infected with respective lentivirus and stimulated by conditional media for 48hrs. Left panel, representative western blots of Caspase3 and Caspase9, right panel, histogram representing. *p<0.05 vs. L media and control lentivirus, †p<0.05 vs. Wnt3a media and control lentivirus.
ILK has received much attention in light of the mechanistic link between aberrant ILK up-regulation and tumor progression in many types of human malignancies including breast, colon, liver, ovary, pancreas, prostate, stomach, and thyroid [24-26]. ILK is well known as a critical regulator in the surviving of cancer cell through the Akt pathway [27]. In addition, the Wnt pathway plays an important role in the maintenance of the balance of the cell number and in the promotion of tumor genesis [28-30]. Recently accumulative data suggest two different roles of ILK in the regulation of the Wnt signaling pathway in mammalian cells [10,28]. Firstly, as an intermediate regulator in the acute stabilization of β-catenin by Wnt3a, ILK may play role in the multi-protein β-catenin destruction complex [31,32]. Secondly, there is a more pronounced effect of ILK inhibition on the nuclear accumulation of β-catenin [33]. Thus, ILK could also play a role in nuclear localization of β-catenin. Our research indicated ILK was able to induce inhibitory phosphorylation of GSK-3β, which modulates activities of transcription factors Lef1/Tcf, including activator protein β-catenin and supported this notion in prostate cells such as PC3 and DU145.
The communication between cells and their microenvironment induces activation or inactivation of intracellular signaling pathways and the survival of tumor cells depend on their ability to adapt to their environment [34]. Impaired gap junction intercellular communication was observed in cancer cells nearly 50 years ago [35]. In the normal colonic epithelial tissue, three connexin isoforms, connexin 26 (Cx26), Cx32 and Cx43, have been shown to be expressed at the protein level [36]. Colorectal cancer development is associated with loss of connexins expression or re-localization of connexins from the plasma membrane to intracellular compartments. It has been reported that ILK activates the expression of YB-1, which facilitates EMT through the translational activation of Snail and other EMT- inducing transcription factors [37,38]. This premise is supported by the finding that genetic knockdown or pharmacological inhibition of ILK could inhibit EMT, in part, through the suppression of Snail expression in PC3 and MDA-MB-468 cells [38,39]. However, to date the role of ILK as an intermediate regulator of GJIC is not clear. Our data suggest ILK inhibits the level of connexins in PC3 cancer cells. These results raise the possibility that ILK promotes tumor progression through blocking the GJIC. We have observed that there is a more pronounced effect of ILK promoting tumor progression. Some data pointed that Wnt3a induced the expression of Cx43 [40,41]. In this paper we found ectopically expression of Cx43, the target gene of Wnt pathway in PC3 cells inhibit the proliferation which is consistent with the results down regulation of ILK. Our hypothesis is that inhibition or genetic depletion of ILK down-regulate the expression of Cx43, and reducing the gap junction communication, resulting the change of tumor progression. Our data suggest a novel mechanism of regulation of the Wnt pathway by ILK in tumor cells. With this work, we have provided new insights of Wnt pathway and provided data suggesting that ILK is a potential therapeutic target of inhibiting tumor cell growth through Cx43.
The authors declare that they have no competing interests.
This work was supported by the National Natural Science Foundation of Zhejiang Province (LY12H16005 to YPY).