Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Review Article - (2016) Volume 5, Issue 3
Agriculture is an indispensable part of every person’s life, ensuring that nutritious and inexpensive food is readily available. As any other organisms, plants are subject to numerous parasitic infections. Biological evolution has allowed plants to produce a variety of toxic compounds to deal with their pathogens. American pokeweed plant (Phytolacca americana) manufactures pokeweed antiviral protein, a ribosome inactivating protein that disrupts protein synthesis and lowers infectivity of many plant and animal viruses. The intricate mechanism of PAP antiviral activity entails a delicate coordination and interplay of several factors, allowing the plant to battle its invaders. Here, we examine the molecular mechanism of this plant peptide, and describe a molecular model of pokeweed’s antiviral activity.
<Keywords: Pokeweed antiviral protein; Ribosome inactivating protein; Depurination
Agriculture continues to be confronted by epidemics, having devastating effects on economies and the plant sources essential for human and animal life. Plants are essential for human and animal life, and encompass natural and landscaped spaces, including forests, crops, nurseries and orchards. Multitudes of microbial pathogens invade and colonize plants, while metabolizing their tissues and disrupting a delicate balance of hormones and nutrients, and in some cases, suppressing gene activity [1-3]. Many plants have evolved to produce natural defense mechanisms that aid in the battle with foreign pathogenic invaders. Plant defense mechanisms include myriad physical and chemical defenses, which prevent pathogens from entering the plant cell, limit their availability, and/or restrict the nutrients necessary for the growth and replication of the pathogen [4-8].
Ribosome inactivating proteins (RIPs) are a group of cytotoxic proteins possessing extremely specific rRNA N-glycosidase activity, and proficient in catalytically inactivating ribosomes, inducing cellular death [9]. The biological effects credited to these protein toxins go back to early times, owing to the high toxicities of the castor bean and jequirity bean [10]. Yet other plants, such as American pokeweed (P. americana) and common soapwort, synthesize pokeweed antiviral protein (PAP) and saporin, that impose lower toxicity on intact cells while inhibiting protein production in cell-free lysates [11-13]. RIPs exert their cytotoxicity by impairing ribosomes [14]; specifically they interfere with the advanced stages of translation [15-19]. While all RIPs exert N-glycosidase activity toward ribosomes, their substrate specificity is greatly diversified. For instance, ricin exhibits vigorous activity toward mammalian and yeast ribosomes while virtually having no significant effect on bacterial or plant ribosomes [20-23]; PAP however is able to depurinate ribosomes from plants, bacteria, yeast, as well as lower and higher animals [15,24-27]. Contribution of the substrate specificity comes from both RIPs and ribosomes alike. RIPs depurinate specific adenines within the sarcin/ricin loop of large rRNA; this introduced lesion interferes with the advanced stages of translation [15,28].
In addition to ribosomal depurination, some RIPs act on naked rRNA (rRNA devoid of ribosomal proteins); some remove multiple adenines and guanines from various nucleic acid substrates including DNA, poly(A), rRNA, and even viral RNA [29-32]. Viral RNA depurination is a matter that is worthy of attention. Duggar and Armstrong [33] have observed that a protein from P. americana possessed an antiviral activity, and inhibited transmission of tobacco mosaic virus (TMV) in plants; though, not until 1978 PAP was accepted as an inhibitor of protein synthesis [25]. While the mechanism of PAP antiviral activity is somewhat unclear, recent findings, produced by the Hudak and Tumer laboratories, show that this activity is not dependent exclusively on inactivation of ribosomes [34,35]. It has been postulated that a direct interaction of PAP with viral RNA (or DNA) is an alternative antiviral mechanism in play. The pokeweed plant produces several isozymes of PAP, all exerting potent antiviral properties [11,13,36-42]. PAP isozymes evoke depurination of genomic HIV-1 RNA [43-45], TMV RNA [46], poliovirus [47], herpes simplex virus (HSV) [48], influenza virus [49], and brome mosaic virus (BMV) [50], among many others, showing a broad spectrum of antiviral activity [13]. This depurination is concentration dependent.
Recent findings have put forward an interesting mechanism for the translation inhibition by PAP [51], where PAP specifically targets and depurinates capped mRNA directly. Hudak et al. [51] generated several PAP mutations (PAPx, an active site mutant (E176V); PAPn, a mutant with a substitution (G75D) in the N-terminal sequence; PAPc, a mutant lacking the C-terminal 25 amino acid residues) and showed that these PAP mutants do not depurinate rabbit or tobacco ribosomes, yet inhibit the in vitro translation of potato virus X and BMV with no notable depurination of ribosomes. These studies showed that PAP is proficient in differentiating capped from uncapped mRNAs, since wild type (WT) PAP, and several of its mutants, prompted inhibition of only capped (but not uncapped) luciferase transcripts. The presence of cap analog decreased the ability of PAP and PAP mutants to inhibit translation of viral RNA, meaning that these toxic proteins are able to identify the cap structure at the 5' end of the mRNA. Examination of the PAP-treated luciferase transcripts reveled that only the capped RNAs were subject to cleavage by acidic aniline, and hence were depurinated in vitro. Based upon these important findings, it was concluded that PAP binding to the cap structure, with subsequent RNA depurination, was the main mechanism for the translation inhibition; depurination of capped viral RNA perhaps, is the primary mechanism for the antiviral activity of PAP [52]. Furthermore, Friedland team have examined interactions between PAP and m7GTP cap analog employing direct fluorescence titrations [53]; this led to quantitative characterization of these interactions.
Although only less than twenty percent of plant positive strand RNA viruses are similarly structured to host mRNA (with a 5'-m7G cap and poly(A) tail) [54], the majority lack one or both of these moieties. Interestingly, PAP also exerts its inhibitory effects on the replication uncapped viruses such as influenza and poliovirus [47,49]. Vivanco et al. [55] have inspected PAP activity against a set of capped and uncapped viral RNAs, demonstrating that PAP does not depurinate every capped RNA, while inhibiting translation of uncapped viral RNAs in vitro without causing measurable depurination at multiple sites. PAP did not depurinate uncapped luciferase mRNA, while depurinating TMV and BMV transcripts, demonstrating that PAP is able to discriminate between capped and uncapped RNAs. No evident depurination of capped alfalfa mosaic virus (AMV) RNA was recorded either. This indicates that the recognition of the cap feature alone is not enough to cause multiple site depurination of RNAs [55]. Furthermore, the team did not record any evident depurination of uncapped satellite panicum mosaic virus (SPMV) RNA, tomato bushy stunt virus (TBSV) RNA, nor uncapped RNA encompassing a poliovirus internal ribosome entry site (IRES); yet, in vitro translation experiments illustrated PAP inhibiting translation of the above viral RNAs [55].
Work published by Wang and Hudak [56] presents confirmation that PAP is able to bind eukaryotic translation initiation factor eIF4G and its isoform eIFiso4G. Studies show that PAP binds specifically to each form, and biochemical and genetic analyses present confirmation that the region of the protein between amino acids 511 and 624 is needed for PAP binding activity [56]. PAP binds to m7GTP-Sepharose; this binding does not reduce the binding of PAP to purified eIFiso4G, indicating that PAP simultaneously forms a complex with eIFiso4G and the cap moiety. In wheat germ lysate translational system, PAP depurinated uncapped transcripts containing a functional WT 3' translational enhancer element (3' TE), but did not depurinate messages containing a non-functional mutant 3' TE [56]. These findings supports a previously postulated hypothesis that binding of PAP to eIF4G (or eIFiso4G) may offer an alternative mechanism for PAP to access both uncapped and capped viral RNAs for depurination. Baldwin et al. [53] have demonstrated that PAP not only binds to the initiation factor eIFiso4G, but that binding of the cap analog to PAP is amplified by this macromolecular interactions, supporting previous findings. This suggests a novel mechanism: PAP interacts with eIFiso4G/eIF4G (as part of eIFiso4F/eIF4F) and interacts with the cap moiety of mRNA. Moreover, addition of eIFiso4E/eIF4E (as part of eIFiso4F/eIF4F) competitively diminishes binding affinity of PAP for the cap, since both proteins are cap-binding [53]. These PAP-eIF interactions possibly unfold the active site of PAP, allowing PAP to recognize target adenine residues for depurination (Figure 1A) [53].
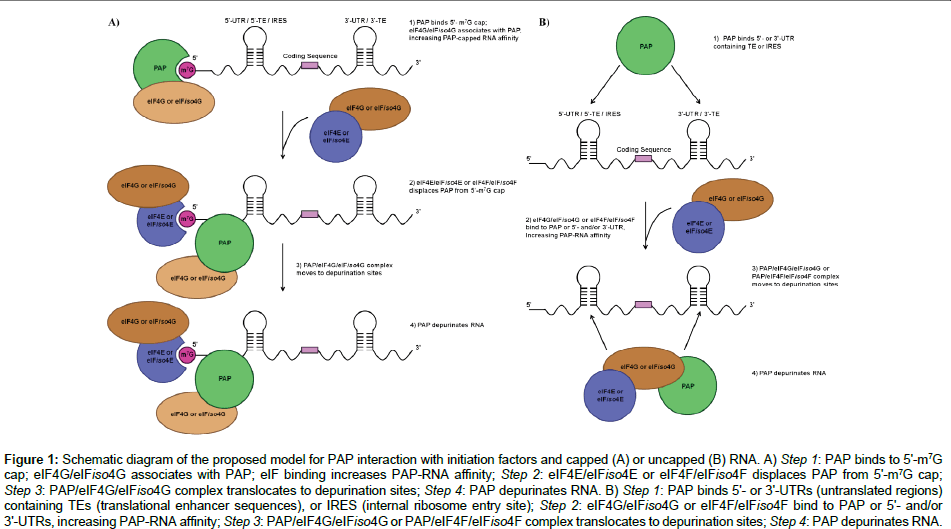
Figure 1: Schem atic diagram of the proposed model for PAP interaction with initiation factors and capped (A) or uncapped (B) RNA. A) Step 1: PAP binds to 5'-m7G cap; eIF4G/eIFiso4G associates with PAP; eIF binding increases PAP-RNA affinity; Step 2: eIF4E/eIFiso4E or eIF4F/eIFiso4F displaces PAP from 5'-m7G cap; Step 3: PAP/eIF4G/eIFiso4G complex translocates to depurination sites; Step 4: PAP depurinates RNA. B) Step 1: PAP binds 5'- or 3'-UTRs (untranslated regions) containing TEs (translational enhancer sequences), or IRES (internal ribosome entry site); Step 2: eIF4G/eIFiso4G or eIF4F/eIFiso4F bind to PAP or 5'- and/or 3'-UTRs, increasing PAP-RNA affinity; Step 3: PAP/eIF4G/eIFiso4G or PAP/eIF4F/eIFiso4F complex translocates to depurination sites; Step 4: PAP depurinates RNA..
Recent work performed in our laboratory showed that PAP binds to and depurinates the 5'-leader sequence from tobacco etch virus (TEV) RNA [57]. The TEV 5'-leader is sufficient to confer cap-independent translation, even in the absence of the 5'-terminal VPg (a viral genomelinked protein) [58,59]. The TEV 5'-leader contains an IRES element [58-60] that is notable for its small size, and represents one of the most compact viral elements capable of promoting cap-independent translation yet identified. A 5'-proximal, 45-nt RNA pseudoknotencompassing domain (PK1) within the TEV 5'-leader is essential to promote cap-independent translation [61]. Mutations disrupting the PK1 reduce cap-independent translation, including mutations to loop 3 that exhibit complementarity to a conserved region in eukaryotic 18S rRNA [61]. Furthermore, PAP bind to both full length TEV leader sequence and the PK1, yet depurinates only the full length leader element, indicating that PAP’s binding affinity is separate from its depurinating activity [62]. Employing a pull-down assay, we showed that PAP binds to eIFiso4G and eIFiso4E simultaneously, and this was confirmed by fluorescence resonance energy transfer (FRET) [63]. These findings support a model, previously proposed by Wang and Hudak [56]: the recognition and depurination of uncapped mRNA by PAP is promoted by the translation initiation factors, allowing PAP to gain access to uncapped, non-polyadenylated RNAs containing either 5'- (and/or 3'-) TEs, associating with eIF4F/eIFiso4F protein complexes (Figure 1B).
The mechanism of PAP antiviral activity, specifically PAP recognition of its substrate viral RNAs, is complex and entails the interplay of a set of different factors. The initial recognition may occur through PAP binding to the cap structure found at the 5'-end of the capped viral genomes, thereby enabling PAP to access its substrates for depurination (Kd for PAP-m7GTP 43.3 ± 0.1 nM at 25°C) [53]. In the instances where the 5'-cap is absent, PAP directly binds to either the 5'- or 3'-UTRs (untranslated regions), containing either translational enhancer sequences or an internal ribosome entry site (Kd for PAPuncapped full length TEV 5'-leader RNA is 28.5 ± 3.7 nM; Kd for PAP-m7GpppG-capped full length TEV 5'-leader RNA is 87.5 ± 4.8 nM) [62]; binding of the eIFs increases PAP-RNA affinity, promoting depurination of RNA (presence of eIFiso4G increases 2.4-fold PAP-cap interactions) [53]. Additionally, PAP isoforms selectivity for different ribosomes and RNAs varies (e.g., PAP-I, found in spring leaves of the pokeweed plant, exhibits RC50 of 1.5 nM towards rat liver ribosomes and 4.7 nM towards E. coli ribosomes [38]; PAP-S1, an isoform found in seeds of the plant, exhibits IC50 of 3.2 nM towards rat liver ribosomes and 280 nM towards E. coli ribosomes [41,42]; whereas α-PAP, expressed in all organs of the plant, exhibits IC50 of 1.3 nM towards rat liver ribosomes and 25 nM towards E. coli ribosomes [13,42]. In recent years, a viral protein (VPg), linked to genome of turnip mosaic virus (TuMV) was shown to inhibit PAP activity in vitro [13,57]. This viral peptide serves as an analog of the 5'-m7G cap of viral RNA, and has been shown to play an important role in mRNA translation since it interacts with the cap-binding proteins (e.g., eIF4E, eIFiso4E, eIF4F, eIFiso4F) [64,65]. Our laboratory has shown that PAP interacts with VPg (Kd is 29.5 ± 1.8 nM) and inhibits PAP’s activity in a dose-dependent manner [57], perhaps providing an evolutionary advantage for the virus to overcome this plant defense mechanism. A valid question to ask would be: If PAP depurinates both ribosomal and viral RNA, how does the pokeweed plant prevent its own death? Recent study shows that PAP is able to form a homodimer complex in the cytosol of pokeweed plant, while its monomeric form is predominantly found outside the cell, the apoplast [66]. The PAP homodimer was shown to be much less active on rRNA in comparison to the monomeric PAP. Hudak et al. have shown that PAP dimerization involves an active site Tyr123; mutations of this aromatic residue prevented dimerization of PAP in vivo, supporting the biological role of homodimerization as a mechanism to limit toxicity to cells synthesizing PAP [66].
In memory of Dr. Diana E. Friendland, who was a very special and integral faculty member at the Department of Sciences at John Jay College, CUNY. Dr. Friendland was actively involved in the development of the Program for Research Initiatives for Science and Mathematics (PRISM), which allows undergraduate students to gain valuable research experience. We also thank Jason A. Domashevskiy for the critical review of this manuscript.