Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2014) Volume 5, Issue 3
We evaluated habitat models that determined relative abundance relationships among microhabitat, reach and watershed-scale factors important for predicting habitat selection. Thirty stream reaches in central Indiana were sampled to determine relationships between habitat associations and relative abundance, size, and age associations of the Northern Clearwater Crayfish, Orconectes propinquus. Females are significantly more abundant than males and the frequency of crayfish in gravel substrate was significantly higher than that of cobble substrate. The sizes of crayfish in cobble substrate were significantly larger than individuals found in gravel substrates, while females were significantly larger than males in gravel substrates. Watershed variables were not significantly related to crayfish abundance. The only reach scale variable that proved to be significant was boulder substrate. Microhabitat variables showed a significant increase between CPUE and cobble and gravel substrates. Habitat models provide valuable information on the conservation status and habitat parameters responsible for determining species preferences, life history strategies, and relative abundance.
Keywords: Habitat suitability, Crayfish, Microhabitat scale, Watershed-scale, Reach-scale
Species responses to various habitat cues provide strong selection preferences that determine aquatic species relative abundance and ecological life history strategies. Crayfish are keystone species in many stream ecosystems since they create and establish habitat use of other sympatric aquatic species. Crayfish are among the largest macroinvertebrates and limit access for other taxa to aquatic food web energy transfer [1-4]. Crayfish are ecosystem engineers that provide a vital role in the structure and function of stream ecosystems [5,6] by determining species distribution and placement within the stream benthic habitat [7,8]. Crayfish are decomposers of organic material and contribute to energy transformation between trophic levels [6,9,10]. They are sensitive indicators of habitat degradation and respond to anthropogenic effects in streams. Species composition and relative abundance reflects anthropogenic response to water quality, habitat, and land use change and stressors [6,11].
The Northern Clearwater Crayfish, Orconectes propinquus (Girard 1852) [12] is ubiquitous in North America ranging from southern Ontario and Quebec, as far south as southern Illinois, Indiana, Ohio, Pennsylvania, and New York, and as far west as Iowa and Minnesota [13,14]. It is a tertiary burrowing crayfish [13] that dig simple depressions in the sediment beneath rocks during drought conditions and spend its entire 2-3 year life history [14-16] in surface water. The species is common in midwestern headwater streams [17] and is found in both stream and lake ecosystems [15,18]. The species’ habitat is typically rocky riffle habitat in streams [14-16,19], and they prefer coarse habitat in lakes that can provide more cover from predators.
Watershed-, reach-, and microhabitat-scale land use can influence chemical and physical factors associated with a stream [20,21]. Watershed-scale land uses are known to have negative impacts on stream ecosystems [22]. For example, agricultural practices increase sediment inputs and nutrients into streams and may negatively affect water quality, habitat, and biological assemblages [20,23]. Excess sediments in streams can negatively affect macroinvertebrates, such as crayfish, by reducing food sources and filling in interstitial pore spaces in preferred habitats [23]. Urban land use has also been found to reduce stream habitat quality by the addition of chemical contaminants [21]. Alternatively, many types of land use can improve stream quality. Forest land use has been found to correlate with high quality habitat and also for bank stability and instream cover [21]. On a reach-scale, the channel morphology can be largely influenced by bank material, riparian vegetation, and the slope at which water and other inputs enter the stream [20,21]. The resulting channel morphology and substrate can determine the types of species that will likely inhabit that particular reach. On a microhabitat-scale cover and substrate particle size can influence individual strategies for feeding, reproduction, and establishment of territories. Factors known to influence crayfish distribution in stream reach scale include presence of predators, amount and stability of within stream cover, age and body size, food sources, and competition among other crayfish [10,24]. Larger crayfish are best able to defend themselves [25] and are then capable of obtaining preferred cover through competition [10].
The primary objective of this study is to determine habitat relationships between catch-per-unit of effort (CPUE) patterns with watershed-, reach-, and microhabitat-scale associations for O. propinquus. Relationships based on gender are correlated with habitat factors, including sediment particle, cover, and larger scale variables. Size, age, and gender patterns in O. propinquus habitat associations are examined in headwater streams to determine selection preferences.
Study area
All of the study sites occurred within the Interior Plateau Level III ecoregion of Indiana [26]. This ecoregion is characterized by rolling and heavily dissected, rugged terrain [26]. The underlying soil is composed of sandstone, siltstone, shale, and limestone [26]. The ecoregion consists of high hills and knobs and low and narrow valleys. The streams of this region are medium to high gradient [26]. Land use is mainly agricultural cropping and livestock pasturing, but includes several forest types. Forested areas were the most common land use surrounding study streams, composing 57% of the total land use.
A total of 30 sites were sampled in the counties of Brown, Monroe, Morgan and Lawrence in southcentral Indiana (Figure 1). Sites are located in the East Fork White River watershed, which is a primary tributary of the Wabash River drainage, Ohio River basin. The East Fork White River watershed is dominated by karst topography and limestone quarries [27].
Figure 1: Study sites (red dots) sampled within the catchment land use during an investigation of Orconectes propinquus habitat suitability preferences in southcentral Indiana, USA, headwater streams. Land use key, Gray = forest; yellow = agriculture; orange = pasture; blue = water features.
Study design
Sites were selected using a random probability study design. Sites were classified by Strahler stream order [28] and selected without replacement from the universe of wadeable first through third order streams in the four county area of the Eastern Corn Belt Plain [29]. A variety of stream conditions were included in the study to determine response to both the highest quality streams and to those of lower qualities due to poor land management practices in southcentral Indiana [30]. Habitat relationships were tested to evaluate the association between relative abundance and CPUE of Northern Clearwater Crayfish based on gender, age, and watershed-, reach-, and microhabitat-scale factors. Our study investigates the relative abundance relationships with gender bias towards microhabitat, size, and scale. We evaluated the percentage of cobble and gravel substrates at each site using a qualitative habitat procedure [31]. Within similar relative abundance categories, we evaluated if greater CPUE of O. propinquus individuals occurred in cobble compared to gravel substrates. We tested two size-related hypotheses, based on whether O. propinquus are larger in size in large coarse substrates, compared to smaller coarse substrates, and whether males are larger than females due to sexual dimorphism or intraspecific competition.
Scale and habitat association hypotheses are placed into three categories, including watershed-, reach-, and microhabitat-scales. For the watershed-scale associations, we determined does the CPUE of crayfish change with land use, while for reach scales, we evaluated whether increasing reach scale habitat heterogeneity led to greater CPUE. For microhabitat scale associations, we tested if CPUE increases with increasing size of substrate. Finally, age was evaluated to determine if ontogenetic differences existed in age class use of large coarse- compared to moderate-coarse substrates.
Field sampling
The stream reach length sampled was 15 times the wetted width [32]. Our study stream reach ranged from a minimum distance of 50 m and a maximum distance of 250 m during this study. Sampling proceeded in an upstream manner beginning at the downstream end of the stream reach, thereby reducing disturbance to upstream crayfish. The sampling events occurred between June 17, 2010 and July 18, 2010, and generally followed the method used by Simon [32]. A one-man common sense minnow seine (1 m x 1 m) with 3.1 mm standard mesh netting was used to sample crayfish by kick-seining a 1 m2 area of substrate directly upstream of the seine [33]. Quantitative collection of crayfish were sampled from 20 one square meter (m2) of habitat randomly distributed in the stream reach, which represent the coarse substrate habitat portions of each stream reach. This was defined as our catch-per-unit of effort. Captured individuals of O. propinquus and all other crayfish species were counted, sexed, and placed into live wells until the completion of sampling. All individuals were released after all site data was obtained.
A total of twenty 1 m2 seine samples were completed at each site [34], 10 samples were randomly located in both gravel-dominated and cobble-dominated substrates (total=20). Substrate size was classified following EPA physical habitat procedures [35] and seine samples were classified as either cobble or gravel when at least 50% of the substrate comprised the dominant substrate type. At each site 10 random m2 samples in each of the two substrate types where sampled using a kick seine method [34] to collect individual crayfish. Stream width measurements at each site included wetted, active, and bank full widths [35]. The wetted width is the perpendicular measurement from shoreline to shoreline. The active width is the perpendicular stream measurement where the normal stream flows fluctuate by season and are delineated by the point where streambed vegetation ends, whereas bank full width measures the lateral extent of water during flooding.
Laboratory methods
Small individual O. propinquus, too small to sex or measure in the field, were taken to the laboratory where carapace length (CL, from tip of rostrum to the posterior border of the thoracic region, to nearest 0.1 mm), postorbital carapace length (POCL; [36]) and sex were recorded. Each individual crayfish was classified as either a male, female, or as a juvenile. A juvenile is defined as the size of the individual that prevented accurate determination of the sex based on the sexual organs not being fully developed. Age was determined based on length-frequency numerical count distributions plots of CL. Crayfish specimens were deposited in the Astacology collection at the Aquatic Research Center of the Indiana Biological Survey, Bloomington, Indiana.
Watershed-scale variables
ArcMap 10.0 was used to overlay the watershed boundary with stream hydrology and 2006 land cover to obtain site data. The stream and land cover data were obtained from IndianaMap.org [37]. The stream layer included the 2008 National Hydrology Data (NHD) streams, rivers, canals, ditches, artificial paths, coastlines, connectors, and pipelines layer. This layer was derived at 1:100,000 scale. The land cover layer included the 2006 USGS 30-meter resolution National Land Cover Data (NLCD). The percentage of each land use type was determined from the land use layer for each individual watershed.
Watershed-scale variables included 15 land cover types, i.e., open water, developed open space, developed low intensity residential, developed medium intensity residential, developed high intensity residential, deciduous forest, evergreen forest, mixed forest, shrub/scrub, grasslands/herbaceous, pasture/hay, cultivated crop, barren land, woody wetland, and emergent herbaceous wetlands [37]; and three additional variables (i.e., latitude, longitude, drainage area). The watershed boundaries were delineated using the Watershed Delineation Model [38], which uses the digital elevation associated with specific latitude and longitude. The drainage areas for each of the 30 sites sampled were obtained from US Geological Survey sources [39].
Reach-scale variables
Reach-scale variables were derived from qualitative habitat measures, i.e., Qualitative Habitat Evaluation Index (QHEI) [31]. The habitat measures include a variety of habitat qualities within the wetted stream width and the riparian area in the floodplain. The qualitative habitat measures include the following categories, i.e., substrate types, instream cover, channel morphology, riparian quality/bank erosion, pool/glide and riffle/run quality, and local stream gradient. The qualitative habitat information was collected from each stream reach. Each qualitative habitat category is ranked by a series of categories representing varying states of stream habitat condition. The total reach habitat score is the sum of each of the category scores, which provides a cumulative score for the entire stream reach. Each qualitative categorical score and the total reach habitat score was regressed against crayfish relative abundance to determine any significant relationships between the habitat category and crayfish relative abundance. Individual substrate particle size categories for each stream reach were compared to crayfish CPUE to determine if any significant relationships existed. The substrate types observed included boulder, cobble, gravel, sand, bedrock, detritus/muck, and artificial. The percentage of each specific substrate size class was determined for each site and used for the comparison. Several other physical reach-scale factors were evaluated including the total percentage of pool, run, and riffle habitat, and the wetted width, active width, and bank full width measurements for each reach. Each parameter was compared to the CPUE of crayfish at each site.
Microhabitat-scale variables
Two microhabitat-scale variables were examined and included the two primary coarse substrate types (i.e., cobble-dominated substrate and gravel-dominated substrate). A CPUE was calculated based on the 10 kick seine samples in each substrate size class and compared to evaluate associations between gender, size, and CPUE with each of the microhabitat substrate types.
Statistical methods
Basic statistics using Statistica were used for all analyses [40,41]. Each statistical analysis conducted used a significance value of α=0.10 for field evaluation and a Tukey HSD post-hoc test. We selected a higher alpha to reduce beta error and reduce Type II errors in data interpretation since our study was based on a single watershed. Differences between relative abundance and CPUE of male and female O. propinquus among cobble and gravel habitats were determined using a Z-test. Differences in crayfish length were determined using an Analysis of Variance (ANOVA) single factor analysis to analyze variance between populations. A length-frequency distribution was developed to evaluate differences in age structure. To analyze each category of the habitat scale factor questions, a simple univariate linear regression was used. The regressions compared a specific watershed, reach, or microhabitat variable with the CPUE of crayfish at each site.
Relative abundance and Catch-Per-Unit Effort
A total of 2,648 O. propinquus was collected from 29 of the 30 (96.7%) sites that were sampled during this study. No other secondary or tertiary burrowing species was sympatric; however, three other primary burrowers were collected during this study including Fallicambarus fodiens (Cottle, 1863), Cambarus polychromatus Thoma, Jezerinac, and Simon, 2005, and C. diogenes Girard, 1852. Orconectes propinquus individual CPUE effort ranged from 0 to 19.1 individuals/ m2 at each stream reach. The number of males compared to females was consistent by site with males comprising 990 individuals and females 1,048 individuals.
The sex ratio was 1:1.05 males to females. A total of 610 juveniles (range: 4.4 mm to 9.8 mm CL) were captured. The number of crayfish captured in cobble-dominated substrates was 989, while 1,049 were collected from gravel-dominated substrates. Females were significantly more abundant than males (Z-statistic=-1.733, P=0.083) within the stream reaches. CPUE of crayfish was significantly (P=<0.001) different in gravel-dominated substrates (Table 1A).
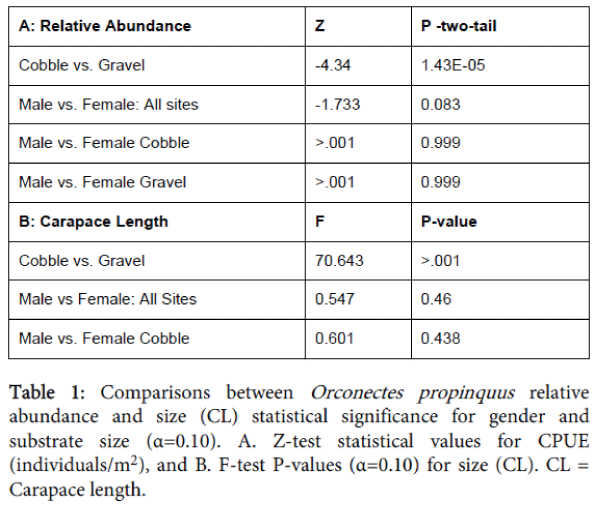
Table 1: Comparisons between Orconectes propinquus relative abundance and size (CL) statistical significance for gender and substrate size (α=0.10). A. Z-test statistical values for CPUE (individuals/m2), and B. F-test P-values (α=0.10) for size (CL). CL = Carapace length.
Length frequency distribution and age range
The mean CL for all of the 2,648 sampled Orconectes propinquus was 12.7 mm. Sampled O. propinquus ranged in CL from 4.4 mm to 39.8 mm. Mean CL was significantly larger for crayfish found in cobble-dominated substrates (P=<0.001) compared to gravel-dominated substrates. Females collected in gravel-dominated substrates had significantly larger CL than males (P=0.013; Table 1B). We observed three age classes based on our study (Table 2A). Both male and female O. propinquus individuals attained similar size at each age. Age 0 individuals were 3-18 mm CL; age I individuals were 18-33 mm CL; and age 2 indivudals were 33-42 mm CL. The length-frequency distribution showed the greatest number of individuals occurred at age 0 (cobble=973, gravel=1285; and males=1433, females=1434). The number of individuals decreased with increasing age group. Only 8 individuals were found in the 2 year age group, and no individuals reached age 3 (Table 2A and 2B). Large individuals (>18 mm CL) had a greater association with cobble substrates compared to smaller individuals (3-18 mm CL), which were associated with gravel substrates (Table 2B).
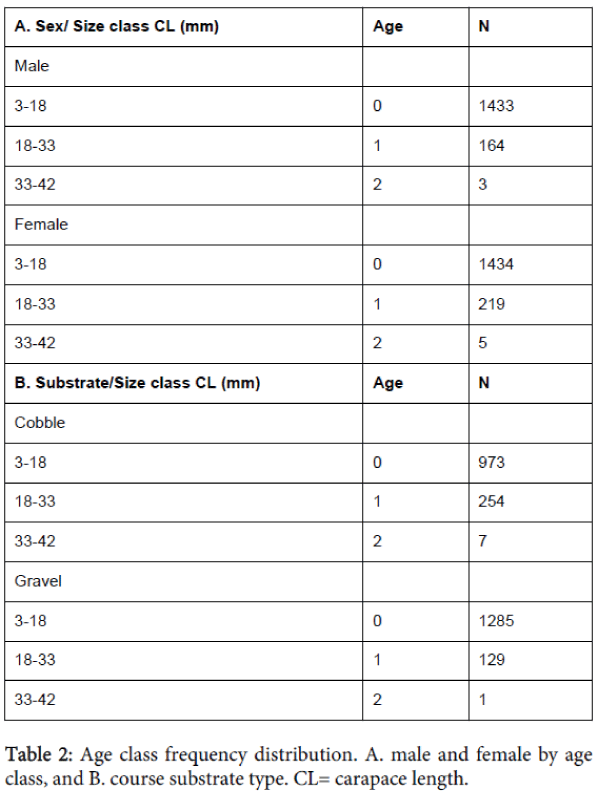
Table 2: Age class frequency distribution. A. male and female by age class, and B. course substrate type. CL= carapace length.
Habitat scale factors
None of the 18 watershed-scale variables showed a significant relationship with O. propinquus CPUE (Table 3). Only a single reach-scale variable, i.e., boulder substrate, was significantly correlated with crayfish CPUE (Table 4). An increasing amount of boulder habitat was associated with a decrease in crayfish relative abundance. Both microhabitat-scale factors, cobble- (R2=0.103, F=3.232, P=0.083) and gravel dominated (R2=0.094, F=2.904, P=0.099) substrates, were positively associated with O. propinquus CPUE.
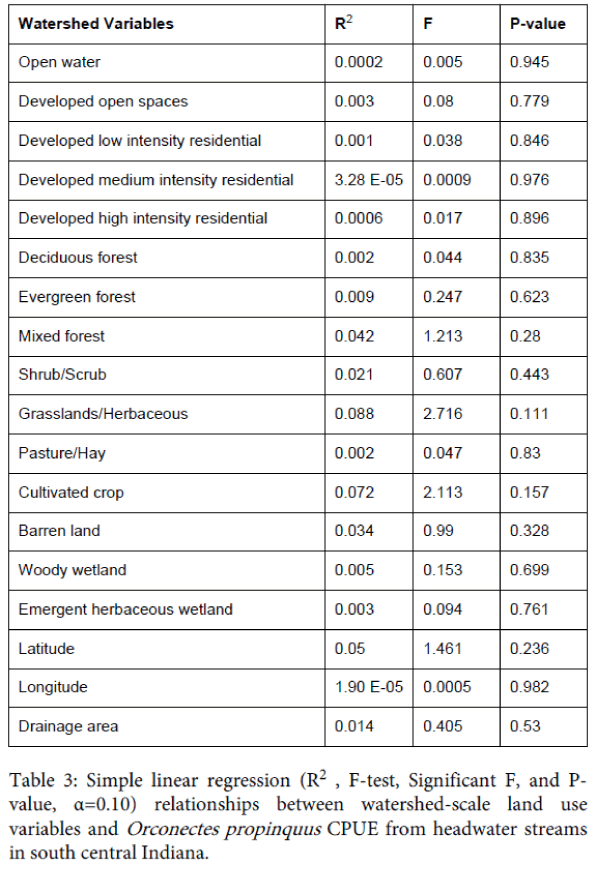
Table 3: Simple linear regression (R2 , F-test, Significant F, and Pvalue, α=0.10) relationships between watershed-scale land use variables and Orconectes propinquus CPUE from headwater streams in south central Indiana.
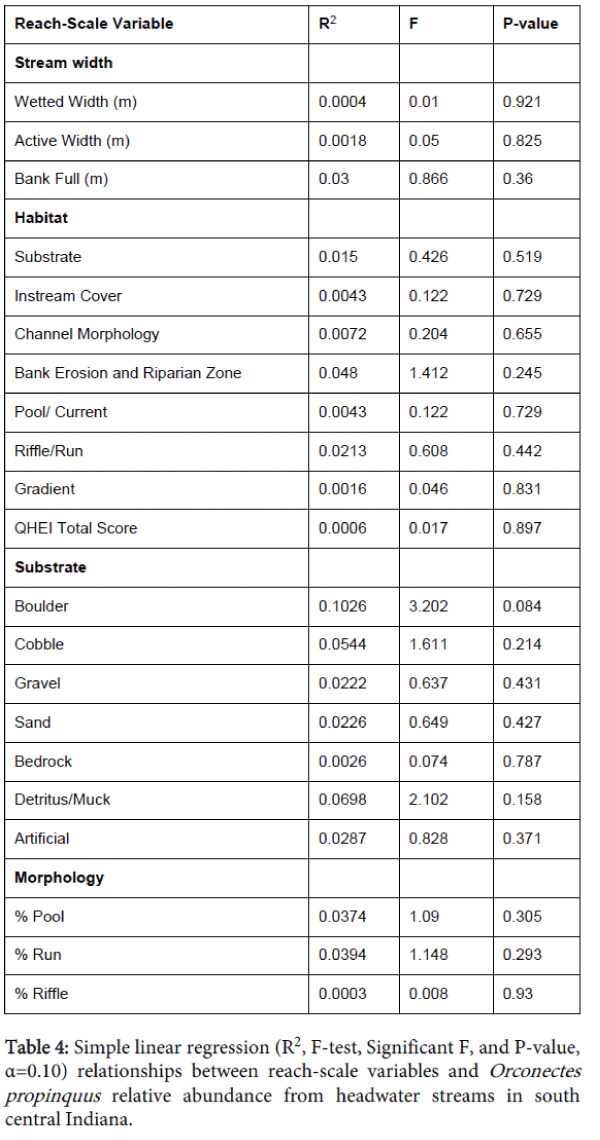
Table 4: Simple linear regression (R2, F-test, Significant F, and P-value, α=0.10) relationships between reach-scale variables and Orconectes propinquus relative abundance from headwater streams in south central Indiana.
The relative abundance of crayfish is dependent on available stream substrate types [10,22,24]. The five lowest crayfish CPUE, i.e., 0, 2, 5 10, and 15 individuals/m2, occurred at sites with reduced reach scale habitat. Bean Blossom Creek (n=2 individuals/m2) was a stagnant stream with muck substrate, while Griffey Creek (n=5 individuals/m2) was heavily impounded with an embedded substrate. These substrate factors are considered responsible for declining crayfish relative abundance due to reduced amounts of preferred substrate and instream cover. Linear regression models showed little significance between scale variables tested at watershed and reach-scales. This was a similar result observed by Burskey and Simon [22] and Stewart et al. [10]. All study area watersheds comprised relatively small drainage area size (range: 9.1 to 49,166 acres). We selected headwater streams to isolate potential impacts and increase the percentage of catchment forested land use. Forested areas provided a large amount of coarse particulate organic matter (CPOM), which are positive factors for stream ecosystems [42]. Forests provide large amounts of organic material and detritus, which are important for crayfish survival [43]. The high percentage of forested areas (mean: 57% for all sites) in these watersheds represent a least impacted condition for crayfish populations. A large amount of forested area is considered to be the most important factor for explaining low significance in watershed scale analyses.
Watershed land cover effects were not found to effect crayfish populations, while other studies linked various land use types to low crayfish abundance [10,11,22,44]. Row-crop agriculture, urban, and developed areas have been shown to negatively impact aquatic habitats and fish and macroinvertebrate communities [11]; however, agricultural land use was not a predominant component in the study streams. Reach scale stream variables scores showed increasing habitat condition levels in the study area (cumulative habitat score range: 37.5 to 91.0, mean=72.9). These relatively high reach scale habitat values show that streams represented relatively high overall ecological integrity. The only variable correlated with crayfish abundance included reach scale habitat substrate boulder proportion. Boulder presence showed a negative correlation with individual crayfish CPUE. This result seems contradictory; however, boulder substrates provide large interstitial spaces affording cover and habitat for predators. The univariate microhabitat-scale regression models showed a significant relationship between CPUE and both cobble and gravel substrates. This suggests that increasing amounts of coarse substrates correlates with increases in O. propinquus CPUE and may be differentially important for various life stages. Crayfish substrate preference is typically associated with the most overall cover and protection from predators [24,45]. Larger crayfish prefer larger substrate sizes since the larger substrates will provide more overall interstitial spaces. These interstitial spaces provide more areal coverage for protection from predators [45]. Crayfish size (CL) was significantly correlated with large substrate sizes compared to small substrates; however, since the study area was not glaciated during the latest Wisconsin glaciation event the dominant particle size is cobble in 321 the study area. Orconectes propinquuss individuals associated with cobble substrates exhibited the highest CPUE (Table 2B).
Large individuals were associated with large substrate particle size and when mature adults are present, smaller individual crayfish typically were associated with small, gravel substrates [10,24]. Orconectes propinquus individuals were significantly more abundant in gravel substrates than in cobble substrates; however, this was based on the association between CPUE and high number of age 0 individuals. Overall, Age-0 crayfish comprised the largest proportion of individual crayfish at all sites (n=2258; 85.3%). A niche shift from small substrates to large substrate occurs at lengths greater than 18 mm CL. This niche shift demonstrates that individual crayfish select increasing substrate particle size proportional to increasing body size. Likewise, small age-1 individuals showed similar response as age-0 individuals with significantly increasing CPUE in the hypothesized less preferred gravel substrates.
Studies have shown that the dominance of many freshwater crayfish is based on size [10,24,46]. The study area male to female sex ratio is 1:1.05 (χ2 (1, N=29), p >0.10), which is not statistically significant. We predicted that male CPUE would be significantly greater than female; however, this assumption was based upon the premise that males would be significantly larger than females. Females were larger than males, but not statistically significant (P=0.460); however, females were significantly larger than males in gravel substrates. This suggests that females could have a slight numerical dominance over males during the early stages of their lives or be forced into smaller substrate particle sizes due to dominance and territoriality. This would explain females being significantly more abundant than males in the study streams sampled; however, females would be exposed to increased predation pressure affecting females CPUE with increasing age class.
Primary habitat drivers effecting O. propinquus relative abundance, niche shift patterns, and age structure included microhabitat- and reach scale factors, but not watershed scale variables. Boulder presence negatively correlated with relative abundance, while cobble was selected instead of gravel substrates. Other cover types including various substrate particle sizes, woody debris, vegetation, and riparian channel factors did not correlate with relative abundance, gender, or size. No watershed scale variables effected O. propinquus relative abundance; however, our study was conducted in headwaters streams and only in a single watershed. Further study in multiple watersheds, larger order streams, or lakes may reveal differing life history strategies.
This study provides important understanding of life history strategies utilized by O. propinquus, which may be useful for management of other related imperiled crayfish species in need of conservation management. Conservation of the habitat heterogeneity and natural stream corridors in forested landscapes provide a unique opportunity to evaluate restoration goals that would promote stable relative abundance. Additional studies of other more restricted species would be necessary to confirm our results that the species is not responding to larger scale stressors.
We thank the Indiana Biological Survey Aquatic Research Center for professional courtesies, use of vehicles, and equipment. Special thanks to Wade Kimmon, Richard Barendt, and Alex Jackson for field assistance.