Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2013) Volume 1, Issue 2
The sublittoral seafloor of the Humboldt Eastern Boundary Current ecosystem (HEBCE) is characterized by sediments rich in sulfur compounds and extensive and sometimes massive benthic bacterial mats. This sulfur-based sublittoral ecosystem is maintained by the very high input of organic matter from the productive surface waters and the concurrence of subsurface hypoxic to anoxic waters of the Peru-Chile Undercurrent (PCU). Generically here named the “Humboldt Sulfuretum” (HS), this benthic habitat extends latitudinally along the continental margins off Peru and Chile for over 3,500 km potentially covering around 100,000 km2 thus making it one of the largest reduced soft sediment regions in the world. Since its first recognition in 1962 the HS has consistently been found the home of massive bacterial mats which can attain standing-stocks in the order of 1,000 gm-2 (wet weight). Although hosting a complex and diverse community of prokaryotes, microbial eukaryotes, and meio- and occasional macrobenthic fauna, the dominant components, both in weight and volume, are large vacuolated species of megabacteria of the Beggiatoaceae family (Gammaproteobacteria). These mat-forming autotrophic/mixotrophic species singularly link the C, S, and N cycles (possibly also the P cycle) and constitute one of the largest macroscopic bacterial populations living in today’s naturally reduced world oceans, i.e., eastern boundary current systems, in fact the HS with its microbial communities could be considered modern analogs of Proterozoic benthic systems. This work intends to highlight the extensive geographic distribution, the huge, although variable, massiveness, and thus the potential biogeochemical significance of the giant mat-forming prokaryotes and their accompanying microbial community, as a contribution to a better understanding HEBCE’s functioning one of the most productive eastern boundary current systems of the world.
<Keywords: Benthic bacteria; Bacterial mat; Microbial eukaryotes; Sulfuretum; Standing stock; Thioploca; Candidatus Marithioploca; Oxygen minimum zone; Eastern boundary current system
A recent work on the biogeochemistry of continental margin ecosystems has identified that one of the major enigmas in the HEBCE is the question of the role played by the oxygen-deficient water masses (Oxygen Minimum Zone-OMZ) and that of its inhabitant’s on the whole eastern boundary current system [1]. This work is a contribution from benthic ecology to help solve the above mentioned enigma. With productivities in the order of 1,000 gm-2y-1 the HEBCE is one of the most productive eastern boundary current systems and consequently its benthic habitat is exposed to one of the strongest organic matter inputs and oxygen stress in the world [2]. By the same token the benthic realm is also rich in reduced sulfur compounds, the ideal environment for the flourishing of ancient prokaryotes [3,4].
Large extant filamentous multicellular photosynthetic megabacteria of the phylum Cyanobacteria have been known from freshwater sediments since the XIX century. However, phenotypically similar marine, non-photosynthetic megabacteria were first discovered in 1962 on the continental margin off northern Chile conforming massive mats in the sulfide-rich sediments under an Oxygen Deficient Layer (OMZ) (Table 1 and Figures 1a-1d) [5-8]. After an initial confusion with “cyanophyceae”, the main bacterial component was identified as belonging to the family Beggiatoaceae, genus Thioploca, a genus at the time only known from freshwater [9].
| Station | Date | S Lat. | W Long. | Depth (m) | Sediment type |
|---|---|---|---|---|---|
| 1 | 07/12 | -18°29’ | -70°22’ | 50 | Putrid mud |
| 3 | 07/12 | -18°29’ | -70°28’ | 114 | Greenish-gray mud, putrid smell |
| 19 | 07/15 | -19°02’ | -70°22’ | 142 | Greenish-gray mud, putrid smell |
| 20 | 07/15 | -19°02’ | -70°20’ | 102 | Greenish-gray mud, putrid smell |
| 45 | 07/25 | -20°48’ | -70°21’ | 282 | Compact clay |
Table 1: Localities where Candidatus Marithioploca mats were found in the northern Chile benthic survey conducted during the “Mar-Chile II Expedition” (1962). Data adapted from Gallardo [5]. Note: at the time the findings were reported as “cyanophyceae”.

Figure 1: A: Collage with photos of the Candidatus Marithioploca system: Portion of a large dredge sample from the Candidatus Marithioploca-dominated microbial mat (CM-DMM) taken at -65 m depth off the Bay of Concepcion, central Chile (~-36°S), during the austral summer of 1975 (similarly rich samples were obtained also with benthic dredges off northern Chile during the 1962 Mar-Chile Expedition. Larger whitish multi-trichomic filaments are about 1 mm in diameter. B: Filaments of Candidatus Marithioploca spp. on a 1 mm2 mesh sieve taken with a quantitative bottom grab at -65 m depth off the Bay of Concepcion, central Chile (~-36°S) during the austral summer of 1975. C: View from above of a box-corer sample showing a dense CM mat. White trichomes are 3-4 cm long. It is this kind of density that standing stock can reach ca. 1 kg m-2. D: Section of a multiple trichome-containing filament of Candidatus Marithioploca araucae where the sheath is a minor component.
The new genus Candidatus Marithioploca (CM) is being now proposed to include the earlier described EBCE’s giant beggiatoaceans Thioploca araucae (cell diameter 30-43 μm and cell length 11-26 μm) and Thioploca chileae (cell diameter 10-20 μm and 7-26 μm cell length) [10] with filament lengths reaching as much as 7 cm [11]. Likewise, less abundant forms earlier identified as Beggiatoa are now renamed Candidatus Maribeggiatoa [12].
The following discussion will center on the above mentioned dominant CMs. These forms are unique in that they constitute giant multicellular threads (trichomes) that can be found free-living or longitudinally disposed within mucous sheaths forming quite visible filaments that in the usual silt or clay bottoms are in general vertically oriented (Figure 8 in Schulz, et al. [13]). Depending on environmental conditions sheaths may contain a few or hundreds of multicellular trichomes involving thousands of cells. Single filaments (sheaths plus trichomes), or even single trichomes, can be clearly seen with the naked eye, and, since CMs seem never to occur as single cells, these megabacteria are amongst the world’s largest [9,14].
As new marine Beggiatoacean megabacteria findings show, their cell volume is occupied by a large internal nitrate-containing vacuole which in the case of the HEBCE’s CMs, it occupies ca. 80% of the cell volume [15], limiting the cytoplasm with its population of sulfur globules to a thin peripheral layer. Interestingly, the sheath may contain a “garden” of endobionts [16], and the surrounding mud, a host of smaller filamentous, multicellular “macrobacteria”, to distinguish them from the “megabacteria” which are at least one order of magnitude larger [3,4] (Figure 2).
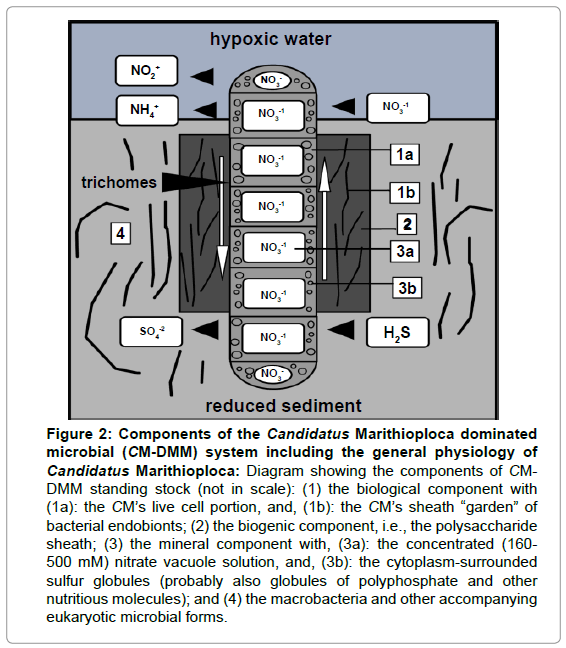
Figure 2: Components of the Candidatus Marithioploca dominated microbial (CM-DMM) system including the general physiology of Candidatus Marithioploca: Diagram showing the components of CMDMM standing stock (not in scale): (1) the biological component with (1a): the CM’s live cell portion, and, (1b): the CM’s sheath “garden” of bacterial endobionts; (2) the biogenic component, i.e., the polysaccharide sheath; (3) the mineral component with, (3a): the concentrated (160- 500 mM) nitrate vacuole solution, and, (3b): the cytoplasm-surrounded sulfur globules (probably also globules of polyphosphate and other nutritious molecules); and (4) the macrobacteria and other accompanying eukaryotic microbial forms.
The biogeochemical importance of CM-DMMs resides in their temporal variabilities and the unusual physiology of the dominant CMs species which have been found to link the biogeochemical cycles of C, S and N [17,18] (Figure 2) and even perhaps also of P [8,19,20]. Within the sulfuretum, it is particularly important CMs nitrate oxidative function which, through the production of ammonium, might play an important role in the productivity of the HEBCE [21]. This is a feature that needs to be acknowledged for the HS and probably for other such systems under oxygen minimum zones [22]. Their potential to determine an important portion of the primary productivity of the HEBCE through the conservation of N by reducing nitrate to ammonium which is readily brought up to the photic zone through upwelling, has already been pointed out by Zopfi, et al. [21,23].
From the bio-geographic stand point, it has consistently been observed that extensive areas of the bottoms under the OMZ off the Chilean and Peruvian continental margins are occupied by CMdominated microbial mat (CM-DMM). During the US JOINT II expeditions to off Peru (October 1976, March and April 1977, and April 1978), the CM-DMMs were found densest in the top three centimeters of sediment, and, in April 1977, it was present down to 20 cm within the sediment. It was further estimated that, since in the thickest parts it occupied about 5% of the sediment volume and had a density of about 1.2 g/cm-3 of filaments, its standing-stock would have amounted to 1,530 gm-2 wet weight or about 90 gcm-2 [24]. Similar high standingstocks were observed off central Chile in December 1975, i.e., 1,060 gm-2 [7] and 800 gm-2 in March 1994 [13].
The two former records spurred a survey off Peru disclosing CMDMM occurrences almost throughout the Peruvian shelf, although with low biomasses [25]. As an explanation Zafra, et al. [26] stated that along the Peruvian shelf, during normal cold, non-El Niño years (1981, 1984 and 1985), bacterial mats were more widespread than during warm El Niño years (1976, 1982/1983) [27]. During non-El Niño regimes, either neutral or cold La Niña regimes, the CM bundles usually constitute larger standing stocks than the co-occurring uni- and multicellular eukaryotic biomass while during El Niño events, the warm phases of the ENSO (El Niño Southern Oscillation) with the intrusion of oxygenrich waters, the situation is reversed [24,27,28].
Besides the interannual variability, seasonal variability is the rule in the southern reaches of the oxygen deficient Perú-Chile Undercurrent, i.e., central Chile. During a benthic megafauna survey off central Chile (1991-1992) [29] mats were found throughout the study area at the deeper stations (-70 and -100 m) but only during summer and early autumn at the shallower station (-50 m) indicating a seasonal deepening of the oxygen-deficient PCU. Seasonality off central Chile is clearly seen in the database used for the present work extrapolations (Supplementary Figure 1).
Thus, seasonal and interannual variabilities in the occurrence of CM-DMMs, the latter related to the EL Niño Southern Oscillation (ENSO) cycle, have been well established and are the response to the prevailing conditions in the pelagic zone-higher (non-El Niño regimes), or lesser productivity (El Niño regimes)-which in turn determine whether a sulfuretum can or not exist (Supplementary Figure 2 and Tables 1 and 2).
| °S Lat. | km2 |
|---|---|
| -6° to -20° | 77,440 |
| -20° to -37° | 22,468 |
| Total | 99,908 |
Table 2: Sea-floor surface area bathed by Peru-Chile Undercurrent (PCU) containing ≤ 0.5 mL L-1 of dissolved oxygen (modified from Helly & Levin [36]).
In recent years, particularly during the development of the Global Program Census of Marine Life (2000-2010) (www.coml.org), the HEBCE region was the subject of new regional observations under the aegis of the International Census Of Marine Microbes (ICoMM) [30]. These efforts included new sampling strategies, i.e., more intensive (time-series) and opportunistic sampling using a variety of tools, i.e. besides the hand-held photography and video, sediment corers (single gravity corers, multicorers and box-corers) and robotic devices (AUV’s, ROVs) equipped with sampling and video capabilities. Biodiversity was addressed with new molecular analytical tools [31].
Most relevant to the present work is, however, the CM-DMM video footages made years before off Peru by scientists from the Harbor Branch “Johnson Sea-Link” manned submarine during the southern spring of 1992. Harbor Branch researchers reported being impressed by seeing a “sediment surface commonly covered” with a “thick, white, Thioploca mat” between -150 to -350 m depth [32]. Likewise, several video recordings have been made of the CM-DMM off central Chile (see Chile video in Supplementary Material). [https://dl.dropboxusercontent.com/u/92999543/Video/Candidatus%20Marithioploca-Dominated%20Microbial%20Mats.mpg]
Naturally gaps of information still prevail on the actual distributions of CM-DMMs in the huge Humboldt Eastern Boundary Current Ecosystem region. What is known, however, is the distribution of hypoxic waters (<0.5 mL L-1 dissolved oxygen) along the coasts of Peru and Chile, an estimated impacted seafloor area of ca. 100,000 km2 between -6°S and about -37°S latitude, creating a favorable habitat for the CM-DMMs (Table 2 and Figure 3).
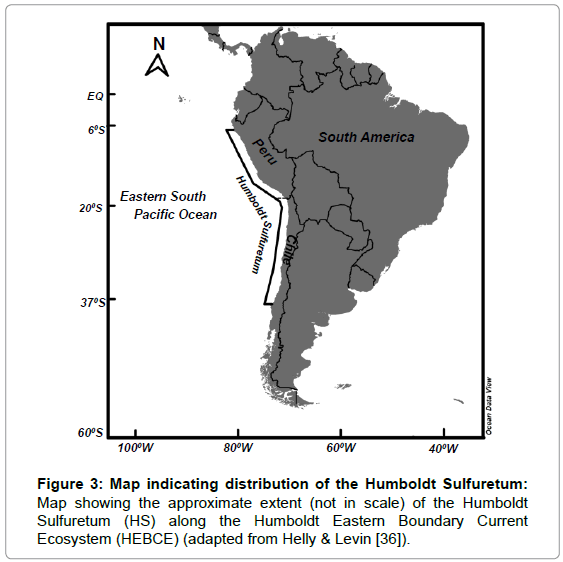
Figure 3: Map indicating distribution of the Humboldt Sulfuretum: Map showing the approximate extent (not in scale) of the Humboldt Sulfuretum (HS) along the Humboldt Eastern Boundary Current Ecosystem (HEBCE) (adapted from Helly & Levin [36]).
Figure 3 in fact depicts the potential distribution of what it is here named the Humboldt Sulfuretum, a sulfur-based system, home of the CM-DMMs. Although the potential for a significant regional standingstock of giant bacteria in the HEBCE region has often been suggested [13], this is the first attempt to extrapolate observed measurements to provide preliminary suggestive information that could spur further research on the CM-DMMs potential role on the functioning and productivity of this highly productive system.
To arrive at a useful view of the potential biogeochemical and ecological significance of the HEBCE’s CM-DMM, we suggest that its standing-stock (grams wet weight m-2, gravimetrically determined) is a composite system as follows: (1) a biological component which in turn consists of: (a) the live cell part of the CM bacterium, and (b) the sheath-dwelling, syntrophic or symbiotic (endobionts) bacteria [16,34]; (2) a biogenic component, i.e., CM’s inert gelatinous polysaccharide sheath [33]; (3) an inorganic (mineral) component, which in turn consist of: (a) the concentrated nitrate solution occupying the large cell vacuole [17,18], and (b) elemental sulfur, polyphosphate and other energetic compounds contained in intra-cytoplasmic globules [19,20,35]; and (4) the macrobacteria, a diverse assemblage of mat-associated smaller filamentous multicellular bacteria [3,4].
This component, however, due to the sediment sieving is mostly lost but preliminary direct observations however indicate that their standing-stock would amount to ca. 10% of the total CMM-DM’s standing stock (unpublished observations). For obvious reasons in the present work only components (1a), (2), and (3a) are explicitly considered (Figure 2).
To estimate the potential regional standing-stock of the CM-DMM in the entire HS, measurements of the actual CM-DMM’s wet weight expressed in gm-2 were multiplied by the estimated sea-floor surface area between -6° and -37°S latitude, affected by PCU/Equatorial Sub-surface waters with dissolved oxygen concentrations ≤ 0.5 mL O2 L-1 [36] (Table 2). The CM-DMM data was obtained from observations at two timeseries stations, one located off Callao in central Peru, covering 1992- 2005 [37], and another from off Concepción, central Chile (Table S1, S2 and S3), covering 2004-2006. Moreover, two extrapolated versions were considered: Version A, based on the estimated proportions of the standing-stock components suggested by Schulz, et al. [13], with emphasis in the biogenic standing-stock component (2) above, and, Version B, our proposition emphasizing standing-stock component (3), the vacuole-contained mineral nitrate solution (Figure 4).
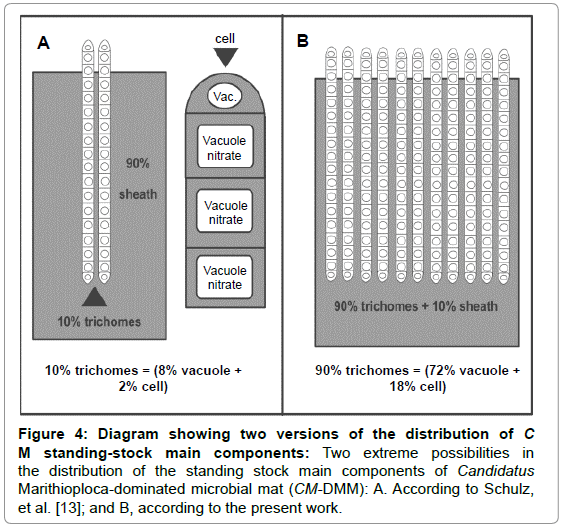
Figure 4: Diagram showing two versions of the distribution of C M standing-stock main components: Two extreme possibilities in the distribution of the standing stock main components of Candidatus Marithioploca-dominated microbial mat (CM-DMM): A. According to Schulz, et al. [13]; and B, according to the present work.
Time-series estimations of the CM-DMM’s standing-stocks measurements during a period typical non-El Niño regime (2004- 2006) rendered similar averages for both the Peruvian (66 gm-2) and the Chilean (86 gm-2) sulfuretums (Table 3). However, because of the size of the bottom surface areas potentially favorable for the microbial mat development, significant standing stock differences between two regions are obtained, i.e., 5.1 million tons for the Peruvian section, 2.7 million tons for the Chilean section, and a total of 7.8 million tons for the whole region (Figure 5).
| Locality | °S Lat. | °W Long. | Depth (m) | Average standing-stock (g m-2) |
|---|---|---|---|---|
| 1. Off Callao, central Peru, 1992-2005 [2]. | -12°02.8’ | -77°17.1’ | 94 | 66.0 |
| 2. Off Concepcion, central Chile; 2004-2007) (data in Table 1-SOM). | -36°30.8’ | -73°07.7’ | 88 | 83.7 |
Table 3: Localities, coordinates, depth, maximum and average standing-stock data for the Candidatus Marithioploca microbial-dominated mat (CMM-DM) from the Peruvian and Chilean time-series stations.
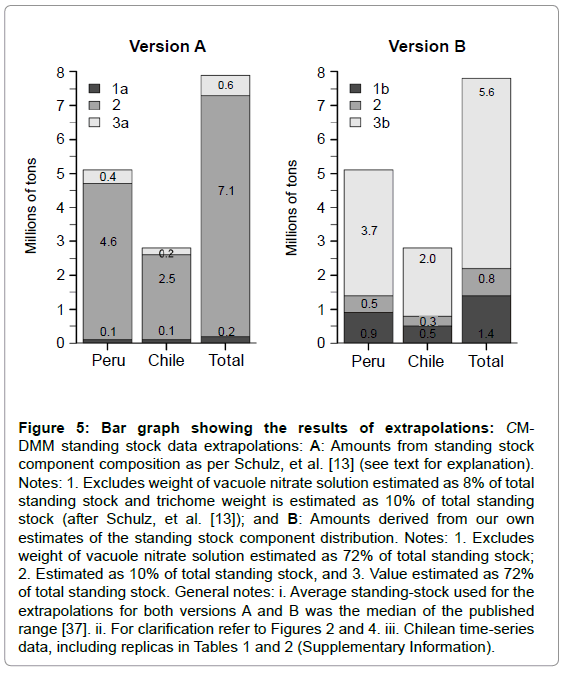
Figure 5: Bar graph showing the results of extrapolations: CMDMM standing stock data extrapolations: A: Amounts from standing stock component composition as per Schulz, et al. [13] (see text for explanation). Notes: 1. Excludes weight of vacuole nitrate solution estimated as 8% of total standing stock and trichome weight is estimated as 10% of total standing stock (after Schulz, et al. [13]); and B: Amounts derived from our own estimates of the standing stock component distribution. Notes: 1. Excludes weight of vacuole nitrate solution estimated as 72% of total standing stock; 2. Estimated as 10% of total standing stock, and 3. Value estimated as 72% of total standing stock. General notes: i. Average standing-stock used for the extrapolations for both versions A and B was the median of the published range [37]. ii. For clarification refer to Figures 2 and 4. iii. Chilean time-series data, including replicas in Tables 1 and 2 (Supplementary Information).
It is also evident that the extrapolations based on the disaggregation of the standing stocks into their main components render two qualitatively different standing stock versions, i.e., Version A with 4.6 million tons for the Peruvian area, 2.5 million ton for the Chilean area and a total of 7.1 million tons of biogenic (sheath) material for the whole region, and, Version B with 3.7 million ton for the Peruvian area, 2 million ton for the Chilean area and a total of 5.6 million tons for the whole region of the mineral (nitrate) component.
The varying oceanographic conditions in the HS could probably determine many possible combinations at the food web and the biogeochemical pathway. At present there is more information on the biogeochemical related issues for off central Peru [38] and for off central Chile [23], than on the former condition. To assess the potential ecological significance for the Humboldt marine ecosystem of the extrapolated CM-DMM’s standing-stock figures consideration must also be taken of the possible number of replications of the CM-DMM standing stock during a year cycle.
The present state of knowledge makes it possible to predict that under normal oceanographic conditions (non-El Niño regimes) these processes will in part depend on the latitude since seasonality decreases somewhat from south to north, probably, at least in part, explaining the larger average standing stock for off central Peru (Table 3).
In fact this average is considered an underestimation because the time series used to calculate the average standing stock includes depressed data from the 1997-98 El Niño. As an indication for example, Levin, et al. [36] reports standing stocks of 3-9 gm-2 during the El Niño off Peru while an average standing stock of 15.1 gm-2 was obtained for off central Chile using data obtained between August 1997 and May 1998. Chilean data used for the extrapolations obviates this effect by using information obtained in a period distant from the large 1997-98 El Niño, i.e. 2004-2007.
The complete set of data (1997-2007) is shown in (Table S1). Under experimental conditions [18] arrived at chemotrophic doubling-times ranging from 69-139 days which could be reduced under mixotrophic (acetate) conditions to 26-52 days. These authors considered the latter figures as underestimations but noted their data was similar to that of Schulz et al. [39] who suggested a 21 day doubling time. For the region off central Chile, with a pronounced seasonality, under normal non-El Niño conditions, about four doubling-times of 52 days would be possible between the beginning of the austral spring and the end of the austral autumn (October through April) a lapse that is considered favorable for the development of the CM-DMM. Off Peru with a somewhat milder seasonality [38], probably a few more replications could occur. These major scenarios are of course completely disturbed during El Niño events when higher temperature and oxygen, and nutrient-poor water enter the region profoundly affecting the Humboldt Sulfuretum ecology [2,38,40,41].
Besides trying to elucidate the potential role in the productivity of the Humboldt Eastern Boundary Current Ecosystem, the study of the giant, vacuolated, filamentous, multicellular marine Beggiatoaceans (CMM-DM) and their accompanying prokaryotic flora, i.e., macrobacteria [3,4], is interesting in that they are the alternative forms that could explain known fossils that until now have been assigned to putative Cyanobacteria. Table S3) includes a selection of the largest and broadest filamentous, multicellular Archean fossil morphotypes extracted from Table 2 in Schopf [42].
These findings range in age from ca. 3,496 million years to 2,560 million years (Archean Eon). The importance of this re-definition is that it does not require free oxygen to appear so early in the geological history of the planet. If this proposition is accepted then it would follow that the Humboldt Sulfuretum could perhaps be considered a present day analog of the Archean primeval marine sulfuretum, a notion that could stimulate more attention from many kinds of researchers on present day sulfuretums than it receives today
Given the results of this assay we posit that the main potential fundamental functions of the CM-DMM in the HEBCE are: (1) a contribution to the C budget through chemosynthesis; (2) a contribution to the N budget in the form of ammonium, which in turn could (3) subsidize pelagic C fixation, and (4) through the utilization of the toxic sulfide serve as mediator for eukaryotic colonization of the benthic realm a course of action that could have occurred in a grand scale during the late Proterozoic Eon [43,44].
We hope that our results will be of interest to specialists in several disciplines from microbiology, through benthic ecology to micropaleontology and astrobiology. This research would also help to view that the so-called “dead zones” are far from being devoid of life and also that the increment of the areas considered “dead zones” may not be too bad after all. Sulfuretums are proving to be genomic-rich realms with potentially important genetic resources [45].
We thank the program Census of Marine Life and FONDECYT Projects 150100007, 1070552 and 1110786.