Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 2
Mutations in the von Hippel-Lindau (VHL) tumor suppressor gene are responsible for the VHL hereditary cancer syndrome, and are associated with the majority of clear cell renal cell carcinomas. In this study, scanning electron microscopy of VHL-negative renal carcinoma cells was utilized to examine the effects of VHL re-expression on the morphology of these cells. Significant differences were observed between the morphology of VHL-negative control cells and those with reintroduced VHL, with VHL expression mediating an apical surface that mounded upward, as opposed to the flat surfaces seen with VHL-negative cells. In long term cultures, rounded VHL-expressing cells grew in clusters on top the monolayer, and microvilli were observed on the apical face of these cells, in a manner suggestive of proximal tubule differentiation. In contrast, VHL-negative cells remained flat and did not develop microvilli in longterm cultures. Since VHL is a key member of an ubiquitin E3 ligase complex whose best known target is hypoxiainducible factor alpha (HIF-α), we looked at the effects of HIF-α expression on cell morphology. Knockdown of HIF-2α in cells that only express this isoform had no effect on the morphology of the cells. These results indicate that VHL expression directs three dimensional morphological changes in renal cells indicative of differentiation, and while dysregulation of HIF-α may be necessary for tumorigenesis following VHL loss, it is not the major determinant of these VHL-mediated morphological changes.
<Keywords: Renal cells, Microvilli, Renal cell carcinomas
HIF: Hypoxia-Inducible Factor; Pvhl19: 19 Kda VHL Protein; Pvhl30: 30 Kda VHL Protein; SEM: Scanning electron microscopy; shRNA: short hairpin RNA; VHL: Von Hippel-Lindau
von Hippel-Lindau (VHL) disease is an autosomal dominant familial cancer syndrome caused by the inactivation of the VHL gene located on chromosome 3p25 [1]. Tumor types that are initiated following loss of the VHL gene include renal cell carcinoma, hemangioblastoma and pheochromocytoma [2]. The VHL gene codes for two protein products of apparent molecular weight of 30 kDa and 19 kDa, referred to as pVHL30 and pVHL19, respectively (collectively referred to as pVHL) [3-5]. pVHL is part of an E3 ubiquitin ligase complex, which targets hypoxia-inducible factor alpha (HIF-α) for ubiquitination and subsequent degradation under normal oxygen levels [6-9]. HIF is a heterodimeric transcriptional factor composed of a constitutively expressed ß subunit and an oxygen-regulated α subunit (reviewed in [10]). HIF plays an important role in the regulation of glucose metabolism, angiogenesis and erythropoiesis under oxygenlimited conditions [10]. Mutations in VHL that abrogate its ability to bind to and/or ubiquitinate HIF-α lead to HIF-α stabilization and the resultant up-regulation of hypoxia-inducible genes [8], which is important for the development of tumors in VHL disease [11].
Previous studies have shown that there are changes in cellular morphology due to the presence or absence of VHL [12-17]. However, there have been contrasting reports of the effects of HIF-α on cell morphology. A reduction in HIF-2α levels was shown to have no influence on the gross morphology of 786-O and A498 renal carcinoma cells [17]. In agreement, cell shape and intercellular junction formation were shown to be independent of HIF-2α levels in 786-O cells [13]. In contrast, it has been seen that intercellular junction formation, but not necessarily cell shape, responds to HIF-α levels. Lowering of both HIF-1α and HIF-2α led to restored amounts and localization of the adherens junction protein, E-cadherin [16], and lowering of HIF-2α levels in 786-O (which do not express HIF-1α) caused a slight to partial restoration of localization of the tight junction protein, ZO-1, although this did not result in obvious morphological changes [18]. Thus, a clear relationship between VHL and HIF-α with respect to cell morphology has yet to be determined. Moreover, the bulk of the findings to date relate to the shape of cells in only two dimensions.
In this study, three-dimensional cell morphological changes of two renal cell lines due to presence or absence of VHL were examined via scanning electron microscopy (SEM). To directly test the role of HIF-α in renal cell morphology, HIF-2α levels were modulated in 786-O cells. We demonstrate that VHL expression has a marked effect on the morphology of renal cells that may be coincident with their differentiation, and that down-regulation of HIF-α is not sufficient to produce these morphological changes.
Cell lines and culture
VHL-negative cell lines, RCC10 and 786-O were obtained as described previously [18]. 786-O cells with control or HIF-2α short hairpin RNA or with reintroduced pVHL19 were created as indicated previously [17]. RCC10 cells with reintroduced pVHL30 were created as indicated previously [18]. All cells were grown in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum supplemented with penicillin-streptomycin (100 U/ml and 10 μg/ ml, respectively), and maintained in 60 mm dishes in a humidified incubator at 37°C in 5% CO2.
Scanning electron microscopy
Cells were cultured on sterile 13 mm Thermanox coverslips (Electron Microscopy Sciences, Hatfield, PA) placed in 35 mm plates. After one, two, three, and four weeks of growth, cells were washed in sterile PBS and immediately fixed in 2.5% glutaraldehyde in PBS. Cells were dehydrated using a graded series of ethanol washes (30%, 50%, 70%, 90%, 95%, 100%, and 100%) for a half hour per wash. Cells were then critical point dried using liquid carbon dioxide in a Denton Critical Point Drier, DCP-1 (Denton Vacuum, Moorestown, NJ). After mounting the coverslips onto sterile SEM metal stubs, cells were sputter coated with gold and palladium in a Hummer 6.2 Hybrid Sputter Coater (Anatech, Ltd, Union City, CA) and examined using a Jeol T300 Scanning Electron Microscope (Japan Electron Optics Laboratory, Commerce, MI) at an accelerating voltage of 20kV. Pictures were taken on Polaroid Type 55 film.
Western blotting
Western blotting was performed as in [18], using rabbit polyclonal anti-HIF-2α antibody from Novus Biological (Littleton, CO), anti- VHL mAb 11E12 [3], and anti-alpha tubulin mAb from Sigma (St. Louis, MO).
VHL influences cell morphology
To test the effects of VHL on renal cell morphology, VHL-negative RCC10 cells (hereafter referred to as VHL-cells) and RCC10 cells in which pVHL30 was stably expressed [18] (hereafter referred to as VHL+ cells) were used (Figure 1A). The cells were grown for one, two, three, and four weeks consecutively on SEM coverslips. Cells were prepared for SEM analysis after each respective week, as outlined in materials and methods. Images were taken at a magnification of 500× and at a 45° tilt. VHL-RCC10 cells (Figure 1B, top panels) had an elongated, flat morphology throughout the progression of the four weeks of growth. VHL+ RCC10 cells (Figure 1B, bottom panels) were more polygonal in shape than their VHL-counterparts. At week 1, there is an appearance of a spherical mound protruding from the apical surface for some cells, and by week 3, the majority of the cells have this mounded apical appearance. At week 4, there is evidence of VHL+ cells growing in multiple layers, with the top layer of cells retaining the apical mounds.
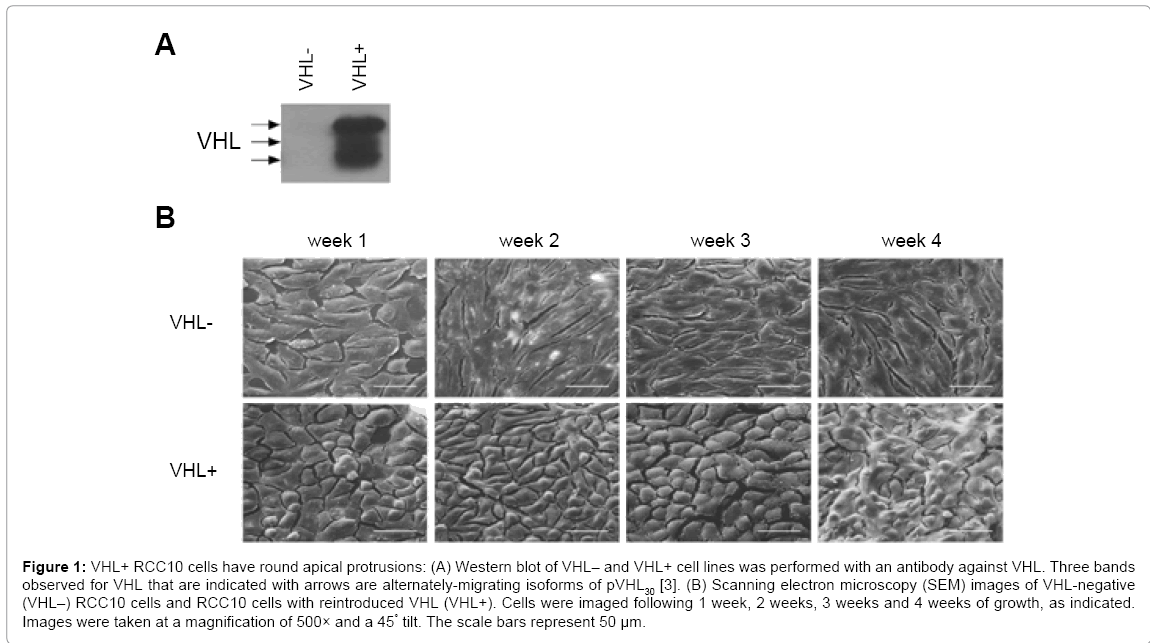
Figure 1: VHL+ RCC10 cells have round apical protrusions: (A) Western blot of VHL– and VHL+ cell lines was performed with an antibody against VHL. Three bands observed for VHL that are indicated with arrows are alternately-migrating isoforms of pVHL30 [3]. (B) Scanning electron microscopy (SEM) images of VHL-negative (VHL–) RCC10 cells and RCC10 cells with reintroduced VHL (VHL+). Cells were imaged following 1 week, 2 weeks, 3 weeks and 4 weeks of growth, as indicated. Images were taken at a magnification of 500× and a 45° tilt. The scale bars represent 50 μm.
To more closely examine the apical surface of the cells, images of the cells grown for 3 weeks were taken at 1500× (Figure 2). Again, VHL-cells were seen as elongated and flat, whereas VHL+ cells smaller and polygonal, with apical mounds. Additionally, the VHL + cells showed small projections from the apical surface indicative of microvilli. Interestingly, a few VHL+ cells growing on a layer above the main monolayer of cells did not have microvilli (Figure 2, bottom right corner of right panel), which may suggest that these are new cells formed by cell division (and thus on a layer above the regular layer) that have not yet fully differentiated.
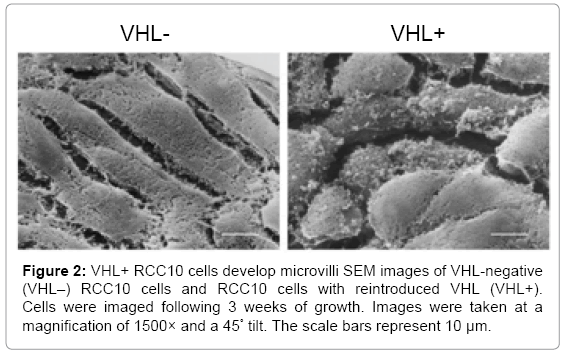
Figure 2: VHL+ RCC10 cells develop microvilli SEM images of VHL-negative (VHL–) RCC10 cells and RCC10 cells with reintroduced VHL (VHL+). Cells were imaged following 3 weeks of growth. Images were taken at a magnification of 1500× and a 45° tilt. The scale bars represent 10 μm.
Lowering levels of HIF-α in VHL-cells does not affect cell morphology
To test the effects of HIF-α levels on cell morphology, we used a set of 786-O cells that included cells in which expression of HIF-2α was knocked down via short hairpin RNA (shRNA)-mediated RNA interference [17]. The cells with lowered HIF-2α were previously shown to undergo cell cycle arrest in a manner similar to VHL replacement, indicating full functional removal of HIF-2α [17]. Also included in this set of 786-O cells were parental VHL-786-O cells, VHL+ cells (with reintroduced pVHL19), and VHL- cells with a control shRNA construct (Figure 3A). All of these cells were grown for two weeks on SEM coverslips and fixed after two weeks of growth. Cells were then prepared for SEM analysis as outlined previously (Figure 3B, top panels). Again, VHL-786-O cells were flat, but seemed to grow in layers, as compared to the RCC10 VHL- cells, which were flat and grew in one monolayer. VHL-786-O cells were not as elongated as VHL- RCC10 cells. The control shRNA cells were similar in appearance to the parental 786-O cells in terms of their flat shape and layered growth. VHL+ 786-O cells appeared to grow up and out of the plane of the monolayer and had a rounded appearance, similar to what was observed with the VHL+ RCC10 cells at 4 weeks of growth. Importantly, the cells with shRNA targeting HIF-2α were flat in shape and layered in growth, similar to the VHL-parental 786-O and control shRNA cells, indicating that lowering of HIF-2α did not restore their morphology to those of VHL+ cells.
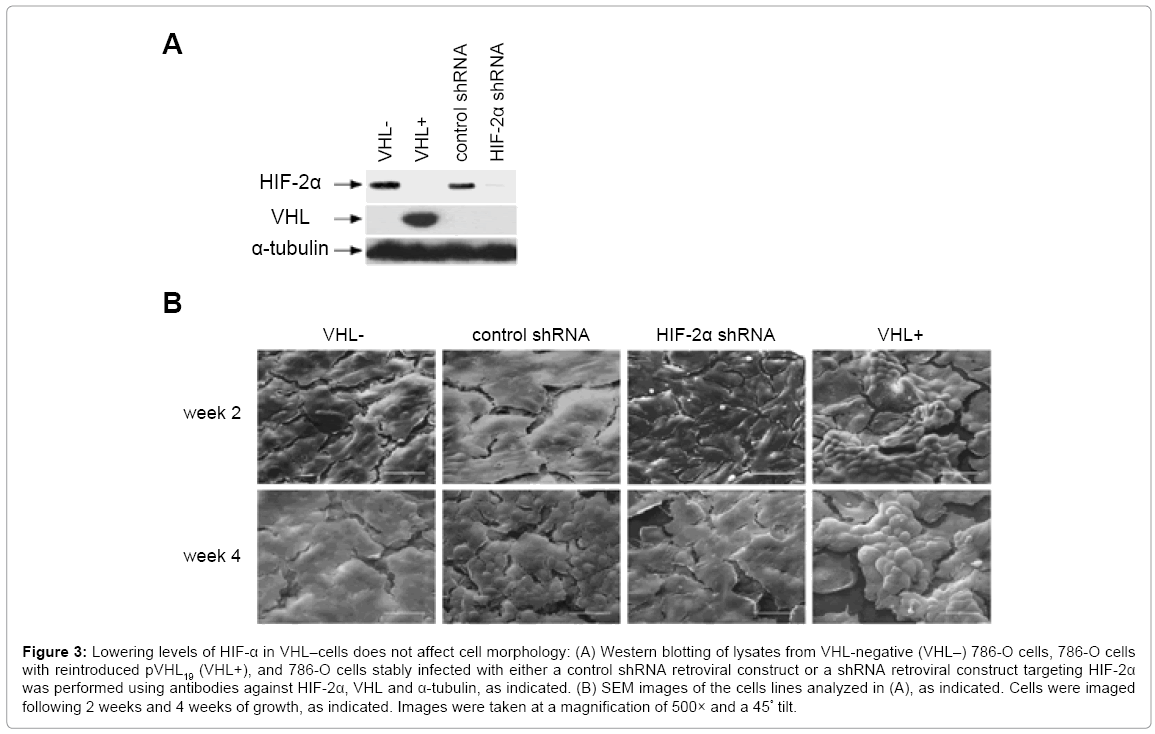
Figure 3: Lowering levels of HIF-α in VHL–cells does not affect cell morphology: (A) Western blotting of lysates from VHL-negative (VHL–) 786-O cells, 786-O cells with reintroduced pVHL19 (VHL+), and 786-O cells stably infected with either a control shRNA retroviral construct or a shRNA retroviral construct targeting HIF-2α was performed using antibodies against HIF-2α, VHL and α-tubulin, as indicated. (B) SEM images of the cells lines analyzed in (A), as indicated. Cells were imaged following 2 weeks and 4 weeks of growth, as indicated. Images were taken at a magnification of 500× and a 45° tilt.
SEM analysis was also performed on the 786-O cell lines after four weeks of growth (Figure 3B, bottom panels). The VHL-parental, control shRNA cells and the cells with shRNA-lowered HIF-2α continued to grow in flat layers, although for reasons that are unclear, the control shRNA cells did show a bit of a rounded appearance. However, the VHL+ cells had a pronounced growth up from the monolayer, apparently reaching several cell diameters high. In addition, these cells grew in clusters, instead of spreading out laterally (Figure 3B, bottom right panel and Figure 4A).
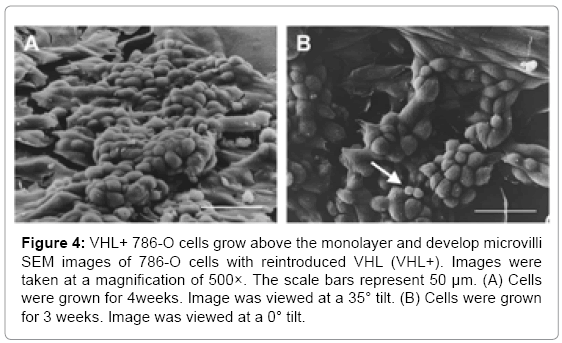
Figure 4: VHL+ 786-O cells grow above the monolayer and develop microvilli SEM images of 786-O cells with reintroduced VHL (VHL+). Images were taken at a magnification of 500×. The scale bars represent 50 μm. (A) Cells were grown for 4weeks. Image was viewed at a 35° tilt. (B) Cells were grown for 3 weeks. Image was viewed at a 0° tilt.
VHL expression leads to microvilli as renal cells differentiate
Since microvilli were observed on VHL+ RCC10 cells, long term cultures of 786-O VHL+ cells were grown to analyze whether they also demonstrated the formation of microvilli. 786-O cells were grown for 3 weeks before being treated for SEM analysis. At a magnification of 500× and viewed from directly above, VHL+ cells had a grainy appearance on the cell surface, suggesting microvilli (Figure 4B). Interestingly, some apparently newer cells, as suggested by their smaller size, had smooth surfaces (see arrow in Figure 4B), which indicates that the prospective microvilli develop as cells mature, presumably as part of cell differentiation. At a magnification of 1500×, microvilli were apparent on the VHL+ cells grown for 3 weeks, but not on similarly grown VHL-cells (Figure 5). Both the VHL- and VHL+ cells appeared to be growing in multiple layers. Notably, a hole was observed in one of the VHL+ cells, which was not observed in any VHL- cells (Figure 5). The microvilli seen on VHL+ cells at lower magnifications were also clearly evident at a magnification of 3500× (Figure 6).
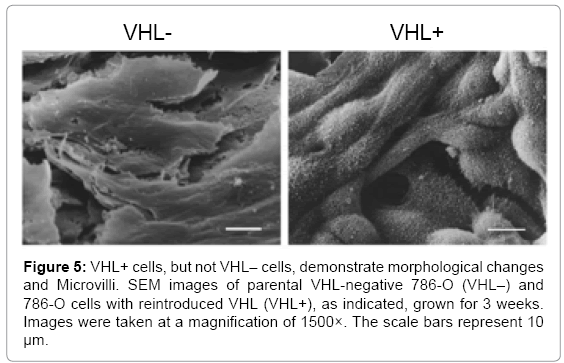
Figure 5: VHL+ cells, but not VHL– cells, demonstrate morphological changes and Microvilli. SEM images of parental VHL-negative 786-O (VHL–) and 786-O cells with reintroduced VHL (VHL+), as indicated, grown for 3 weeks. Images were taken at a magnification of 1500×. The scale bars represent 10 μm.
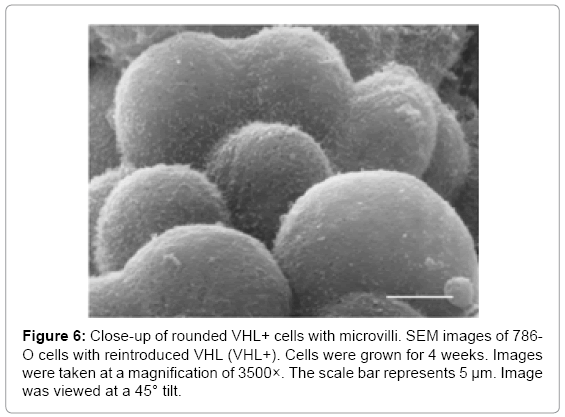
Figure 6: Close-up of rounded VHL+ cells with microvilli. SEM images of 786- O cells with reintroduced VHL (VHL+). Cells were grown for 4 weeks. Images were taken at a magnification of 3500×. The scale bar represents 5 μm. Image was viewed at a 45° tilt.
In this study, the morphology of two VHL- renal carcinoma cell lines were compared to the corresponding derivative cells with VHL stably reintroduced. In both cell lines, it was seen that VHL expression mediated a mound-like protrusion of the apical face of the cell, with growth of cells perpendicular to the plane of the culture dish and the presence of microvilli on cells cultured for some time. Previous studies have shown that expression of VHL elicits changes in cell shape in two dimensions [12-17], and alters the growth of cells grown as spheroids [19]. An increased height of VHL+ cells has been noted as well [13,14]. However, to our knowledge, this report is the first to fully describe three-dimensional differences in individual cells due to VHL expression, including a transition to a more rounded cell and the formation of clearly visible microvilli that develop with time in culture.
It is likely that the morphology observed here for VHL+ cells is a result of cell differentiation. It has been reported that in 786-O cells, VHL expression in combination with long-term culturing leads to upregulation of leucine aminopeptidase and hepatocyte nuclear factor 1α (HNF-1α), both markers for differentiation of renal proximal tubule cells [12]. RCC10 cells have also been shown to undergo biochemical changes consistent with renal differentiation [18]. The VHL-mediated morphological differentiation seen here was very similar for 786-O and RCC10 cells. While the cell of origin of these two cell lines is not totally clear, it is likely that both 786-O and RCC10 cells are derived from the proximal tubule, since clear cell renal carcinomas are believed to originate from the proximal tubule (reviewed in [20]). In agreement, both of these cells demonstrate VHL-mediated microvilli formation, which is characteristic of brush border cells found in the proximal tubule. Thus, findings here agree with previous data that VHL expression is important for the differentiation of proximal tubule cells. Of note, both pVHL19 and pVHL30 demonstrated the ability to direct morphological differentiation.
Unlike previous reports [14,15], we did not observe the formation of primary cilium as a result of VHL expression in our cultures. However, different culturing conditions were employed here that may have precluded the formation of cilia. As a result of longer times in culture, we observed VHL+ cells to grow upwards, on top of each other (Figure 4A), which may appear to contradict the tumor suppressor role of VHL. However, this growth of cells should not be considered indicative of increased cell division or a reversion to a malignant phenotype. The 786-O VHL+ cells used here, unlike the VHL- cells, have been shown to be unable to form tumors in nude mice [3], and the RCC10 VHL+ cells used have been shown to have lower levels of cyclin D1 and increased levels of p27 in long-term cultures than the corresponding VHL- cells [18]. Thus, it is likely that the upward growth observed here for VHL+ cells is a consequence of these cells attempting to organize into a tubular structure, as a result of proximal tubule cell differentiation, as has been previously suggested [19]. Note that VHL- cells also showed layered growth, but these cells remained flat and seemed to lack the cell-to-cell adhesion that was seen with VHL+ cells, again suggesting that the upward growth of VHL+ cells is a result of differentiation. Knockdown of HIF-2α in 786-O, which previously demonstrated a cell cycle arrest in these cells [17], did not seem to alter either the morphology or three-dimensional growth of the VHL-cells, suggesting that properties of VHL in addition or alternative to HIF-α regulation are necessary for the morphological differentiation that was observed for VHL+ cells. Since loss of differentiation is an early step in renal cyst formation and in renal cell carcinoma, the ability of VHL to bring about the described morphological differentiation is undoubtedly important for its tumor suppression properties.
We thank from Dr. Algernon (Jerry) Churchill (Adelphi University) for his guidance with the scanning electron microscopy. Research reported in this publication was supported by the National Cancer Institute of the National Institutes of Health [award number R01CA010234].