Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2018) Volume 7, Issue 1
search for drugs that can target cancer cell microenvironment in as much as being able to halt malignant cellular transformation, the natural dietary phytochemical curcumin was currently assessed in DMH-induced colorectal cancer rat model. The study enrolled 50 animals divided into a control group (n=10) and DMH-induced colorectal cancer control group (n=20) (20 mg/kg.-body weight for 28 weeks) versus curcumin-treated group (n=20) (160 mg/kg suspension daily oral for further 8 weeks). Treatment by curcumin succeeded to significantly decrease the percent of ACF and tended to normalize back the histological changes retrieved in adenomatous and stromal cells induced by DMH. The drug also significantly elevated GSH and significantly reduced most of the accompanying biochemical elevations (namely MDA, TNF-α, TGF-β and COX2) observed in colonic carcinomatous tissue, induced by DMH, thus succeeding to revert that of MDA, COX2 and TGF-β back to near normal as justified by being nonsignificantly altered as compared to normal controls. The only exception was PAF that was insignificantly altered by the drug. When taken together, it could be concluded that curcumin possess the potentiality to halt some of the orchestrated cross-talk between cancerous transformation and its microenvironmental niche that contributes to cancer initiation, progression and metastasis in this experimental cancer colon model. Envisioning these merits to a drug with an already known safety preferentiality, awaits final results of current ongoing clinical trials, before curcumin can be added to the new therapeutic armamentarium of anticancer therapy.
Keywords: Curcumin; Dimethyl hydralazine; Aberrant crypt foci; Malondialdehyde; Reduced glutathione; Cyclooxygenase-2; Tumour necrosis factor-alpha; Transforming growth factor-beta; Platelet activating factor
Colorectal cancer is one of the highest prevailing malignancies worldwide and ranks second to lung cancer as a cause of death among adults, despite the progress in its diagnosis and the advancement in its treatment [1].
Such therapeutic advancement built up, along the last decades, when the molecular basis of individual susceptibility to colorectal cancer, its genomic, epigenomic and other evolutionary factors promoting its initiation, driving its progression and contributing to its invasion, local spread and distant metastasis, were all unraveled [2-4]. However, the therapeutic targeting of many of these cues in colonic tumor cells, seemed still not enough to bridge the discrepancy of limited incurability and high death rate.
This prompted, endless efforts, to further probe and explore, in hope to refine our understanding. In this respect recent data support the notion that the acquisition of colonic epithelial cells to its inherent hallmarks of carcinogenesis [5], relies in part on its synergistic crosstalk and interaction with its surrounding microenvironment namely; stromal infiltrating cells [fibroblasts, immune cells (neutrophils, macrophages, lymphocytes, etc.)] and other non-stromal cellular structures as extracellular matrix and vasculature [6-8].
This reciprocal relationship, implies that host microenvironment does not stand as an innocent by standers but at very early stage, will recruit chronic inflammatory infiltrates that intend to exert antitumor actions [9]. A little latter with time, its dynamic response to the epithelial carcinogenic signals will stimulate secretion of mediators (PGs, PAF, etc.).
Cytokines [8] (TNF-α, ILs, ect) [7,8], growth factors (VEGF, TGF- β1, etc.) [10,11], and expression of receptors and enzymes (COX-2) [8,12], that will switch the ongoing reaction to favor epithelial mesenchymal transition, angiogenesis, intravasation, circulation, extravasation and colonization to local and distant tissues [7,8,13].
In light of this recent understanding, the eagerness to find therapeutic options that can also target the varied cues of microenvironment contributing to colonic carcinogenic growth, invasion, spread and metastasis, in as much as it targets tumor cell machinery directly, is a current therapeutic ambition [8,14]. This is because if such potentiality would be existing, it will hold hope to bridge the discrepancy in mortality rates, still reported in relation to the therapeutic outcome of some cancers, as colorectal cancer.
In this respect, curcumin (diferuloylmethane), the phytotherapeutic derived from rhizomes of turmeric (used as spices ever since the early history of mankind), was of current concern. Its anticancer propensity strike via a multiple edge sword mechanism to attack tumor cells (as that of colorectal cancer). This enrolls an ability to arrest cell growth via modulation of cell cycle regulatory machinery by suppressing growth factors and proto-oncogenes and modulating checkpoint regulatory genes [15-17]. It also encompasses a utility to induce apoptosis by expressing pro-apoptotic cues, repressing anti-apoptotic proteins and preventing telomere shortening. More-over curcumin is capable of halting the invasive ability of tumor cells by preventing tumor-induced loss of T-effector immunomodulation [15,16,18].
No wonder, it was of current interest, in this study to expand on a possible potentiality of curcumin to target some of the signaling molecules involved in tumor cell-microenviroment cross-talks, that are proven to be contributing to colorectal carcinogenic transformation, growth, invasion and dissemination [8].
If so is the case, then curcumin would rank among agents targeting the different facets of carcinogenicity, with a logic propensity of better improving the therapeutic outcomes of colorectal cancer. Moreover, when considering its safety preferentiality being a natural dietary phytochemical [19,20], this would tend to further contribute to an improvement in the overall mortality outcomes of a disease, like colorectal cancer [19,20].
Animals
The work was conducted on 50 adult male Wistar rats, weighing 220-250 g each. They were obtained from the Animal Care Center, College of Pharmacy, King Saud University. All animals were fed a standard rat chow and water ad libitum. They were housed in groups in a temperature-controlled environment (20-22°C) with an alternating 12 hours light-dark cycle. Animals were handled with care throughout experimentation, in concordance with the strict guidelines issued by the National Institute of Health for Experimental Care. The experimental design set and the procedures followed throughout the work were approved by Institutional Ethical Committee for Animal Care, King Saud University, Riyadh, Kingdom of Saudi Arabia.
Experimental Design:
After 2 weeks of acclimatization, animals were randomly assigned into two experimental groups;
1. A control group (10 animals); received a weekly intraperitoneal injection of 0.1 ml normal saline vehicle, throughout the period of experimentation (36 weeks).
2. A DMH-induced colorectal cancer group (40 animals); received a weekly intraperitoneal injection of 1,2 dimethyl hydralazine [DMH] (E. Merck Ltd., Bombay, India) dissolved in normal saline vehicle at a dose of 20 mg/kg.-body weight [21], for 28 weeks. After that period, those rats were further divided into 2 equal subgroups.
a. A control colorectal cancer group; received 5 ml/kg daily oral distilled water by gavage, for a further of 8 weeks.
b. A curcumin treated colorectal cancer group; received 160 mg/kg curcumin suspension in distilled water by gavage [22], for a further of 8 weeks.
At the end of experimentation (36 weeks), all rats were fasted overnight before termination day. They were anaesthetized by deep ether anesthesia before being sacrificed. Via a post-mortum laparotomy, the colon (from caecum to 1 cm above the anal sphincter) was immediately excised and very quickly cleaned with ice cool saline to remove faecal residues. It was then bathed into ice cool Krebs buffer (pH 7.5) where it was incised along its mesenteric border and gently rinsed. It was then instantaneously transferred on ice cold plate, with mucosal surface up, for macroscopic scoring and evaluation by three independent expertise that were non-cognizant of the different experimental subgroupings.
The tumors were easily discernable from the inflamed colonic segments and a descriptive notion regarding their multiplicity and incidence, their size and their carcinogenic burden were recorded within their macroscopic scoring. Accordingly, the nature of each tumor was preliminary segregated as being an adenoma (preneoplastic) or carcinoma relaying on whether invasion existed or not through the muscularis mucosa. Only lesions showing clear cut invasion were considered potential carcinoma for further verification while the rest were classified as adenomas and were excluded from the counts.
The three main axes of each potential carcinogenic lesion were then measured using a Vernier Caliper with 0-1 mm graduation and a biopsy was quickly taken from each lesion for conducting the Aberrant Crypt Foci [ACF] Assay. Accordingly, only those lesions that met the criteria given by bird [23] for carcinogenic transformation were the ones finally enrolled in this study. These lesions were instantaneously dissected out and divided on into 2 pieces; a portion for running the histopathological examination and the other portion was further divided and stored at -80°C till conducting the varied biochemical estimations [pro-oxidant/antioxidant variables (MDH and GSH), inflammatory/angiogenic variables (Cox-2, TNF-α, PAF) and epithelial-mesenchymal transition variable (TGF-β1)].
ACF assay
The colonic lesion biopsies were stained by 30% methylene blue for light microscopic examination [using a 40 magnification to transluminate the specimens] in search for lesions fulfilling Mc Lellan and Bird criteria for carcinogenic transformation. These mainly include the presence of crypts of increased size with a thicker and deeply stained epithelial lining and increased peripheral zone girth as compared to crypts with normal architecture [23].
Histopathological examination
The full thickness specimens were fixed in 10% formol saline prior to wax embedding, sectioning and staining with haematoxylin and eosin for light microscopic assessment.
Biochemical estimations in colonic tissues
Malondialdehyde (MDA) assay: This reflects the extent of lipid peroxidation induced by oxidative damage within the colonic lesions, as MDA is the end product of such oxidative inflict. The principle is based on the reaction of MDA with thiobarbituric acid (TBA) (E. Merck Ltd., Bombay, India), according to the method described by [24]. The concentration of MDA-TBA complex was being quantified, measured spectrophotometrically at 532 nm and expressed as n mol/g wet tissue.
Reduced Glutathione (GSH) assay: This reflects the antioxidant potential of colonic lesions. The principle is based on the reaction of reduced GSH with 5.5-dithiobis-(2-nitrobenzoic acid (DTNB) (E. Merck Ltd., Bombay, India), according to method of Owens and Belcher [25]. The absorbance was measured by Schimadzu double beam spectrophotometer (UV200S) at 412 nm. The amount of GSH present was calculated using a standard solution of GSH containing 1 mg of GSH/ml of 3% metaphosphoric acid. The increase in the extinction at 412 nm was proportional to the amount of GSH present and was expressed as n mol/g wet tissue.
Cyclooygenase-2 (COX-2) activity measurement: This in part reflects the extent of inflammatory reaction within colonic tissue. The principle is based on measuring the peroxidase activity of COX according to the method of Kulmacz and Lands, using the COX Activity Assay Kit, following steps as per manufacturer instructions (R and D Systems, Minneapolis, USA). It was measured spectrophotometrically at 590 nm and COX-2 activity was distinguished from that of COX-1 by the isoenzyme specific inhibitors provided. It is expressed as n mol/min/g wet tissue.
Tumour necrosis Factor-alpha (TNF-α) determination: This in part reflects the extent of inflammatory reaction within colonic tissue. It is determined according to the method of Reinecker et al. [26] using an Enzyme-Linked Immunosorbent Assay Kit, following steps as per manufacturer instructions (R and D Systems, Minneapolis, USA). It is measured spectrophotometrically at 532 nm and expressed as pg/g wet tissue.
Platelet Activating Factor (PAF) activity measurement: This, in part, reflects the extent of inflammatory reaction and angiogenic triggers within colonic tissue. It is determined according to the aggregation assay method of Wallace et al. [27] which determines PAF induced aggregation to aspirin treated, differentially centrifuged prepared platelets, in presence of adenosine diphosphate scavengers, by using a Chronology Corporation Aggregometer. It is expressed as pg/g wet tissue.
Transforming Growth Factor-Beta (TGF-β) determination: This, in part, reflects the extent of epithelial-mesenchymal transition within colonic tissue. It is determined according to the method of Javelaud and Mauviel [28] using the quantitative Sandwich Enzyme-Linked Immunosorbent Assay Kit, following steps as per manufacturer instructions (R and D Systems, Minneapolis, USA). It is measured spectrophotometrically at 450 nm and expressed as pg/g wet tissue.
All data are expressed as mean+standard error of the mean(S.E.M.). Statistical analysis was performed by SPSS (version) statistical software. Unpaired Student t-test was used to compare between the different studied groups. P values <0.05 were considered significant.
Biochemical results
Table 1 and Figure 1A-1F showed that administration of curcumin at a dose of 160 mg/kg daily for 8 weeks in DMH –induced colorectal cancer (taken once weekly for 28 weeks) caused significant reduction in MDH, TNF-α, TGF-β and COX2 in colonic tissues as compared to DMH carcinogenic control group with the mean values 0.31 ± 0.010, 7.71 ± 0.42, 0.34 ± 0.031 and 0.42 ± 0.045 respectively. (p<05), While curcumin produced non-significant reduction in PAF as compared to DMH group with a mean value of 21.05 ± 3.9 (p>05).
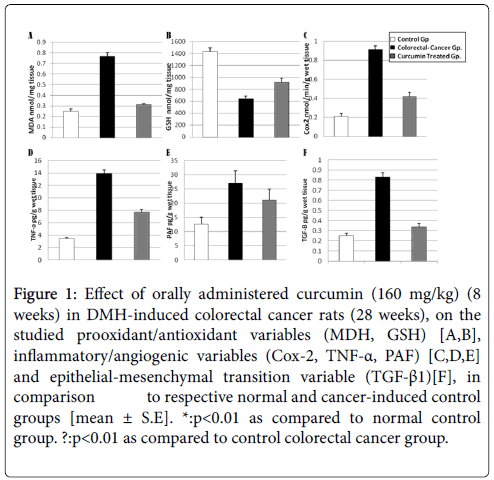
Figure 1: Effect of orally administered curcumin (160 mg/kg) (8 weeks) in DMH-induced colorectal cancer rats (28 weeks), on the studied prooxidant/antioxidant variables (MDH, GSH) [A,B], inflammatory/angiogenic variables (Cox-2, TNF-α, PAF) [C,D,E] and epithelial-mesenchymal transition variable (TGF-β1)[F], in comparison to respective normal and cancer-induced control groups [mean ± S.E]. *:p<0.01 as compared to normal control group. ?:p<0.01 as compared to control colorectal cancer group.
| Biochemical Parameters Studied | MDA nmol/mg tissue | GSH nmol/mg tissue | COX2 nmol/min/g wet tissue | TNF-α pg/g wet tissue | PAF pg/g wet tissue | TGF-β1 pg/g wet tissue |
|---|---|---|---|---|---|---|
| Normal Control Gp. | 0.25 ± 0.021 | 1436 ± 57 | 0.21 ± 0.032 | 3.47 ± 0.12 | 12.6 ± 2.48 | 0.25 ± 0.027 |
| Control Colorectal- Cancer Gp. | 0.77 ± 0.034* | 639 ± 46* | 0.91 ± 0.041* | 13.92 ± 0.58* | 27.08 ± 4.28* | 0.83 ± 0.042* |
| Curcumin Treated Colorectal-Cancer Gp. | 0.31 ± 0.010· | 920 ± 65*· | 0.42 ± 0.045· | 7.71 ± 0.42*· | 21.05 ± 3.9 | 0.34 ± 0.031· |
Table 1: Effect of orally administered curcumin (160 mg/kg) (8 weeks) in DMH-induced colorectal cancer rats (28 weeks), on the studied prooxidant/ antioxidant variables (MDH, GSH), inflammatory/angiogenic variables (Cox-2, TNF-α, PAF) and epithelial-mesenchymal transition variable (TGF-β1)], in comparison to respective normal and cancer–induced control groups [mean ± S.E.]. n: 10 rats in normal control groups. 20 rats in control colorectal cancer groups. 20 rats in curcumin treated groups. *p<0.01 as compared to normal control group. ?p<0.01 as compared to control colorectal cancer group.
Curcumin induced significant increase in GSH level in colonic tissues as compared to DMH carcinogenic group with the mean value of 920 ± 65 (p<0.05).
Curcumin succeeded to revert only MDA, COX2 and TGF-β to near normal as justified by binding them non-significantly altered as when compared to the normal control.
Histological results
Colon of normal control group displayed normal colonic appearance with no signs of apparent abnormality (Figure 2).
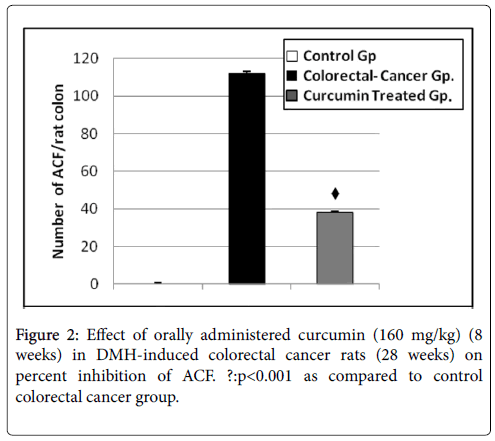
Figure 2: Effect of orally administered curcumin (160 mg/kg) (8 weeks) in DMH-induced colorectal cancer rats (28 weeks) on percent inhibition of ACF. ?:p<0.001 as compared to control colorectal cancer group.
Aberrant crypt assay
The effect of curcumin on the growth and development of ACF, induced by DMH, in rats was given in Table 1. There was a remarkable decrease in the incidence of aberrant crypts, expressed in terms of percentage, from 100% in carcinogen group to 66.07% in curcumin treatment group in. The average yield of aberrant crypts for carcinogen group was 112 ± 3.2 ACF/colon and the range was 90-115 ACF/colon. For curcumin treatment group, this was significantly reduced (P<0.001 when compared to carcinogenic group) to a mean value of 38 ± 3.1 and ranged between 15-40 ACF/colon. There were no observable foci witnessed in normal control rats receiving saline throughout the experiment (Figure 2).
In DMH-colorectal cancer group, the carcinogen produced visible macroscopic neoplastic tumors that can be easily discernable in the inflamed sections of the colon (Figure 3). In curcumin treated group no signs of neoplasia were seen (Figure 4a and 4b).
Figure 3: Colon of normal control group displayed normal colonic appearance.
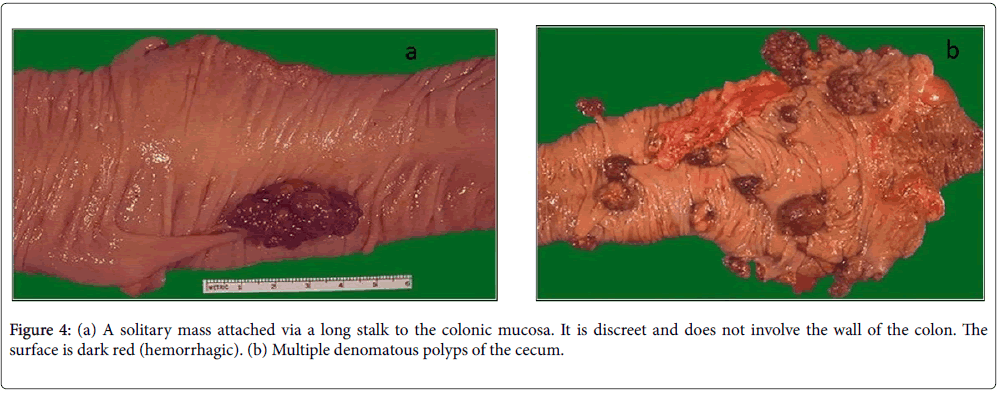
Figure 4: (a) A solitary mass attached via a long stalk to the colonic mucosa. It is discreet and does not involve the wall of the colon. The surface is dark red (hemorrhagic). (b) Multiple denomatous polyps of the cecum.
Microscopic results
Tissue sections of normal control group showed normal colonic architecture with no signs of any abnormality (Figure 5).
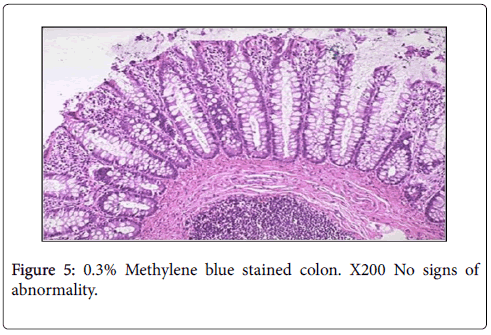
Figure 5: 0.3% Methylene blue stained colon. X200 No signs of abnormality.
In DMH-colorectal cancer group. Large numbers of aberrant crypt foci were present. In addition, DMH-induced mucosal adenocarcinomatous changes. The nuclei were enlarged, hyperchromatic with mitotic changes, tending to colocalize at its surface. Signs of invasion to surrounding stroma were seen. DMH induced also transmural necrosis, edema and submucosal polymorphic cellular infiltration in the surrounding microenvironmental area. (Figure 6a-6c).
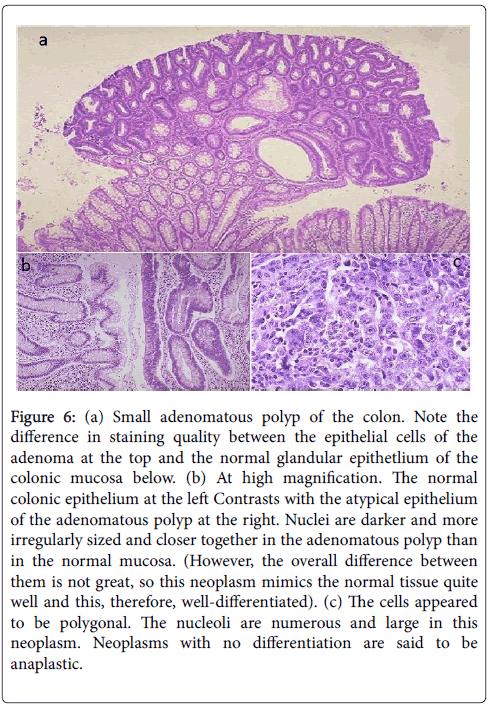
Figure 6: (a) Small adenomatous polyp of the colon. Note the difference in staining quality between the epithelial cells of the adenoma at the top and the normal glandular epithetlium of the colonic mucosa below. (b) At high magnification. The normal colonic epithelium at the left Contrasts with the atypical epithelium of the adenomatous polyp at the right. Nuclei are darker and more irregularly sized and closer together in the adenomatous polyp than in the normal mucosa. (However, the overall difference between them is not great, so this neoplasm mimics the normal tissue quite well and this, therefore, well-differentiated). (c) The cells appeared to be polygonal. The nucleoli are numerous and large in this neoplasm. Neoplasms with no differentiation are said to be anaplastic.
Treatment with curcumin for 8 weeks showed evidence of improvement, normalizing histological changes. Most areas of the colonic tissues showed no signs of neoplastic changes, with the exception of very few areas with crowding cells at the surface, no aberrant crypt foci were present, no adenomatous polyps, no edema or infiltration of polymorphonuclear leucocytes were sighted (Figure 7a and 7b).
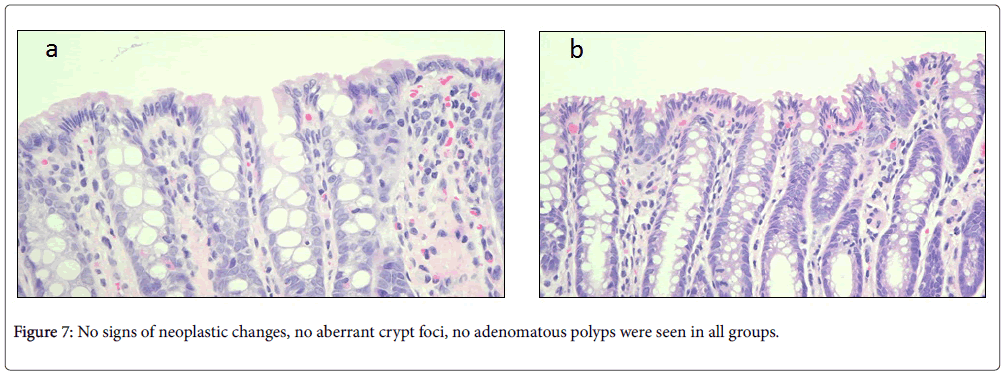
Figure 7: No signs of neoplastic changes, no aberrant crypt foci, no adenomatous polyps were seen in all groups.
The therapeutic armamentarium of anticancer therapy is recently webbing in many intersecting domains, in trial to improve by all means, the mortality outcome of such devastating threat as that of colorectal cancer [2] of such new domains, is the search for drugs that can target cancer cell microenvironment [7,8,29]. The current interest in that, resided on recent recognitions, linking the acquisition of cells to their inherent six hallmarks of carcinogenic transformation [5], to their concomitant synergistic interaction with their cellular and noncellular stromal surroundings [30]. And that only then, tumorogenesis is in progress, and that the journey becomes paved for its local and metastatic invasion and distant colonization [7,8,30].
Accordingly, the therapeutic insight to find a drug that can target tumor-stromal cross-talk, looks promising, especially if it can concomitantly halt the malignant transformation within cancer cells themselves i.e., achieving a bidirectional benefit [7,30]. Such a drug, would even be better, if it happens to simultaneously possess a safe profile on non-malignant cells as well [19,20].
That is why curcumin, the polyphenolic compound derived from dietary spices, was the candidate of interest in this study, being known for its safe non-toxic profile on normal cells in as much as its potent anticancerous potential; suppressing proliferation, inducing apoptosis, etc. within malignant cells [15,16,31]. Such has been reported, in vitro and vivo experimental and in human studies [32-34] and verified clinically in prevention and treatment of cancer as colorectal cancer [20,35-37].
Accordingly, what seemed prudent in this study, was probing in the possible ability of curcumin to target some of these cross-talk signals supporting cancerous initiation, unrestrained growth and ability to detach and disseminate. This is in hope, to expand its therapeutic utility to encompass more and more strategies to combat colorectal carcinogenesis; knowing that the prognosis and therapeutic outcome of this devastating disease, recently was reported to greatly relay on such tumor-stromal microenvironment interactions [7,8,38].
In this domain, a major contributor to such microenvironment is inflammation (and its related inflammatory mediators) that is now considered the seventh trait acquired by colonic cancer cells to survive, grow, and metastasis [7,14,39,40]. Such cancer-associated inflammation is recruited, secondary to a disturbed pro-oxidant inflict posed by the colonic carcinogenic transformation, in the face of a suppressed anti-oxidant stromal buffering capacity interactions [7,38,40]. This misbalance can be anticipated to exist in the current study whereby a significant suppression in GSH and an elevation in MDA, was observed in DMH-induced colorectal group.
This intensified present concerns on a role for curcumin to reset back such misbalance, in the current study; whereby a significant reduction in MDA and elevation in GSH was observed on comparing treated versus DMH-induced cancer groups, the MDA levels were tending to return to normal as judged by being non-significantly changed with respect to that in control group. Such antioxidant potentiality posed by this phytochemical has been a common finding in many previous works assessing its antioxidant utility whether in treatment of cancer or other immune or degenerative disorders [16,41,42]. The likely mechanism is claimed to be via its direct chain breaking antioxidant ability, that will scavenge oxygen species implicated in perioxidative damage of biomembranes [4,43,44] which helps to explain its current induced decrease in MDA. This aside claims of an indirect antioxidant potentiality that enhances GSH synthesis [4,45] which has most likely contributed to the present observed elevation in GSH, produced by curcumin. It was further explained that it is either the hydroxyl group or CH2 group of the β- diketone moiety of this phytochemical that hold responsible for exerting this unique antioxidant activity [4,44]. In this respect, a word of notion delineates that such potentiality can limit the chronic inflammatory recruitment within the microenvironment, at dose ranges within that used in this study, and that at higher doses, curcumin would exert pro-oxidant inflicts that activate caspases to induce apoptosis of colonic cancer cells [46-48].
Beyond modulating the pro-oxidant-antioxidant misbalance, curcumin was also shown to exert a profound anti-inflammatory effect in the current work when comparing the changes in inflammatory markers in treated versus DMH-induced colorectal cancer groups. This therapeutic potentiality is of crucial importance knowing that cancerassociated inflammation is pivotal to colorectal carcinogenesis and that the key player laying at the cross-road of all this has been explicitly reported to be the transcription factor NF-κB [8,40,41,49,50]. This in turn further expresses downstream; COX-2, TNF-α, iNOS, interleukins, metalloproteinases, cell cycle regulatory proteins, etc that all orchestrate tumor microenvironment cross-talk characteristic to colonic tumerogenesis [8,16,41,51].
Accordingly, the retrieval of a significant elevation in COX-2 activity, in the present DMH-induced colorectal cancer group, is in common line with the so many reports declaring its over expression in the majority of colonic carcinogenesis [52]. This being further supported by experimental and observational studies correlating longterm use of NSAIDs to a 40–50% risk reduction in colorectal cancer prevalence and an improved prognosis (judged by colorectal-specific and overall deaths) in patients therapeutically treated by it, after being diagnosed [53]. And since COX-2 is essential for prostanoid synthesis, the yielded prostaglandin E2 (PGE2) was moreover found to be proneoplastic [54] interplaying to induce both tumor cell proliferation, survival, and dissemination and microenvironment supportive recruitment [7,55-57].
On the contrary, curcumin administration did currently succeed to significantly lower COX-2 activity, as compared to that found in DMH-induced colorectal cancer group, as part of its overall observed anti-inflammatory potentiality. This was not surprising, because this phytochemical has been extensively used as if being a “herbal aspirin” since more than 2000 A.D. in Asian traditional medicine [15,58] and is even now comparable to steroids, NSAIDs and coxibs [4]. Literatures have linked the methyl group of curcumin, to the mediation of such anti-inflammatory activity [16,44], via suppressing many of evoked inflammatory cytokines and chemokines. This was ascribed, to be provoked through modulation of COX-2 activity both at transcription and post-transcription levels [4,31,59]. Transcriptionally, this is being orchestrated by inhibition of MAPK, which subsidiary inhibits IκK to pertain NF-κB bound to IκBα in the cytoplasm. As a result, curcumin, will not allow this universal transcription factor to exert its tumorpromoting role; as that in colorectal cancer, thus preventing its translocation to the nucleus to express many of such downstream regulated pro-inflammatory and antiapoptotic genes nor its related protooncogenes [44,60], of which TNF-α is one of its target and was of equivocal concern, in the present work.
Thus, observing a parallel significant decrease in TNF-a, in treated versus DMH-induced cancer group, that returned back towards normal as judged by its finding insignificantly altered as compared to control group, seems promising. This relays on knowing how that parallels comparable data weighing curcumin ability to suppress such inflammatory marker whether in similar [16,48,60,61] or in other cancerous transformation [15,31,62,63]. Some of such studies, did verify that blocking NF-?B pathway and its downstream signaling, is the way, to halt TNF-a induced NF-?B reactivation, whereby blocking of such feed-back loop is central, in curcumin anticancerous propensity in curtailing the vicious circuit of inflammatory-oncogenic interplay [15,18,64].
This pivots on notions clarifying, how that during colorectal carcinomatous transformation (like in many other tumors), TNF-α is the main inducer utilized by cancer cells, to ignite NF-κB- regulated antiapoptotic cascade within them, in the face of their stromal niche, with intention of maintaining self-growth and invasion [65-67]. Vice versa, in response, the microenviroment tries to encompass such growth by creating an inflammatory milieu, that intends to hamper tumor progression. In such a setting of altered immune response, posed by so many intersecting signals, the released inflammatory mediators in reality end up by perpetuating rather than co-localizing the cancerous transformation, thus permitting progression and allowing invasion [7,8,68].
An integral paradigm in this scenario, is the parallel induction of angiogenesis, fostering tumour vascularization, to sustain more growth and to start metastasis. This is achieved via the increased production of various angiogenic molecules [as VEGF, bFGF, EGF, COX-2, TNF-α, IL-8, HIF-1, ….etc] whether by tumor cells themselves and/or by their infiltrating stromal inflammatory cells, specially mast cells and macrophages [7,8,69].
At this juncture, it is worth to point out, that PAF, the known phospholipid mediator of inflammation, is a major player in the game. It is recognized as the proximal inducer mediator which ignites NF- κB-regulated angiogenic cascade calling for the expression of all aforementioned angiogenic triggers [70,71]. Its finding significant elevated in DMH-induced cancer versus control groups in the present work could be interpreted in light of its justified role in promoting angiogenesis for cancer progression.
Meanwhile retrieving PAF reduced, though non-significant when comparing curcumin treated versus non-treated groups, warrants questioning in the present study. This is because previous literature did confirm that the drug possesses many antiangiogenic potentialities, via repression of angiogenic growth factors, inflammatory mediators, matrix proteinases, plasminogen activators, adhesion molecules or by expression of antiangiogenic factors like TIMP or through disruption of cytoskeletal remodeling and mechanotransduction pathways [72-74]. However, its role on PAF-induced NF-kB-regulated angiogenic cascade was scarce. This gives pace to raise a probability that the used dose of curcumin in this study might not have been enough to suppress that pathway, in as much as it did in halting NF-kB anti-inflammatory cascade. This being speculated in light of the concomitant significant reduction in COX-2 and TNF-α but not in PAF by the drug, currently observed. This postulate needs further justification.
A third-dimension integral to carcinomatous transformation, is their phenotypic changes during the process of epithelial-mesenchymal transition. In this context, TGF-β [produced by both cancer cells (as colorectal cancer cells themselves) along with tumor-associated macrophages and fibroblasts] was reported to be the key initiator that induces and maintains such transition (may be through some autocrine and paracrine pathways) [7,8,13,75,76]. This will invite detachment of cancer cells, allowing for their invasion and metastasis. Some have proposed that this is associated with cytoskeletal remodeling and increased cell motility [77]. Others stated that it was triggered secondary to the concomitant increase in TNF-α which can induce expression and DNA binding of AP-1 to result in increased transcription of the TGF-β1 gene [78,79]. This permits it to persist over expressed even during colonization in local and distant tissues working in concert with the simultaneous released angiogenic triggers [7,80,81]. This theme, is relevant to the current findings, where by TGF-β1 was observed to be significantly elevated when comparing its levels in DMH-induced cancer versus control groups, mirrored by a similar change in TNF-α retrieved in the current work.
It is prudent to emphasis, that this should not be admixed with statements highlighting the paradoxical role of TGF-β in colorectal cancer predisposition versus progression. This has been elegantly explained by demonstrating how TGF-β signaling pathways act initially as tumor suppressors to colonic epithelium inducing apoptosis early during cancerous transformation. At a later stage, the changes posed by the surrounding microenvironment switch it to become a tumor promoter, acting as an oncogene to enhance epithelialmesenchymal phenotypic changes leading to tumor survival, invasion and metastasis [2,80,82-84].
In reverse, curcumin administration succeeded presently to significantly reduce TGF-β, when compared to its levels in DMHinduced cancer group and set it back towards that found in normal controls. Reports previously confirmed that the drug can inhibit key steps in TGF-β signaling pathway by being an AP-1 inhibitor [48,85,86]. While others declared that this key transcription factor down-regulates TGF-β in cancer cells, in the context of tumor to host immune interplay, to sustain the host immune surveillance, but they did not link that, to its potentiality in driving epithelial-mesenchymal transition [15,87]. A different party proved that curcumin can lower TGF-β levels by working upstream to suppress TNF-α, that will in turn halt its downstream AP-1 expression, but this was in a setting other than carcinogenesis [79,88]. Likewise, proposing that the suppression of that interrelated cascade of TNF-α/AP-1/ TGF-β1 activation, could best explain the encountered findings in TGF-β1 currently observed by curcumin in this study, needs further verifications before a final statement is declared.
When taken together, the aforementioned etiopathogenic scenario in link to each of the studied pro-oxidant/anti-oxidant, inflammatory/ angiogenic and epithelial-mesechymal transition variables, matches and logically explains the simultaneous encountered yield in ACF and histopathological cancerous and stromal alterations detected in the studied cancer group. This was not surprising, as DMH (the colonic carcinogen) currently used to induce the adenocarcinomatous changes in rats, was comparably reported before, to evoke similar pathological alterations in relevance to most of the raised biochemical variables currently studied [61,89].
Vice versa, the improvement in pathological findings currently encountered after curcumin treatment would be the logic outcome of the tackled multiple anticancer propensities hereby raised or more overly justified in the plethora of literature addressing the therapeutic potentialities of this phytochemical [35,47,48,89].
Thus, the current finding of a 66.7% reduction in ACF yield and a significant reduction in retrieving histopathological cancerous and stromal alterations in the treated group of this study seems promising and mirrors equivocal pathological findings in previous elsewhere experimental and clinical reports probing in curcumin anti-cancerous propensities [4,31,73,90].
In conclusion, finding a safe therapeutic utility, that can target equivocally the orchestrated cross-talk between cancerous transformation and its microenvironmental niche contributing to cancer initiation, progression and metastasis is a paramount need in anticancer therapeutic advancement. Curcumin possesses such potentiality but envisioning these merits into an eventual clinical outcome is awaiting the final say of its current ongoing clinical trials [15,16,19,20,37,48].
The authors extend their appreciation to the CMRC and the Deanship of Scientific Research at King Saud University for funding the work.