Forest Research: Open Access
Open Access
ISSN: 2168-9776
ISSN: 2168-9776
Research Article - (2014) Volume 3, Issue 4
A detailed understanding of the laws and processes that determine the ecosystem dynamics is essential in order to develop well-adjusted forestry management plans in light of climate change. This includes a good understanding of natural regeneration processes since natural regeneration can become an important component of stand resilience. Moreover, climatic changes may reduce the success of natural regeneration and hence require adjustments to silvicultural practices. This work analyzes the effect of two different soil treatments and different stand densities on the initial Pinus nigra Arn. ssp salzmanni seedlings growth at six testing sites located in the Serranía de Cuenca (Spain). Preliminary results showed that soil preparation does not affect initial seedlings growth since differences between soil preparation techniques were always not significant (P>0.05). This work also showed that Spanish black pine initial seedlings growth tended to be higher at the most common and favourable habitats than in the marginal populations. Further studies are necessary to better understand the specific influences across sites and under changing climatic conditions.
<Keywords: preparation; Brushing; Scalping; Basal area; Spanish black pine.
The Mediterranean forests have a rich biodiversity, are attractive for tourism and recreation, and have a great potential for high value forest products. In this context, sustainable forest management must encompass the maintenance of the ecosystem’s integrity to support multiple users. In spite of the hot and dry season in the Mediterranean region, and water deficits during the growing seasons, natural forest regeneration problems can be overcome by selecting silvicultural characteristics of trees, which are best adapted to the specific ecological conditions of the Mediterranean eco-regions.
Spanish black pine (Pinus nigra Arn) is a taxonomically complex species found in the Mediterranean region (Greece/Turkey and the Iberian Peninsula), and there are various subspecies with similar adaptive capacity and low genetic differentiation [1]. Spanish black pine forests have been included in the Europe Union endangered habitats listing of natural habitats requiring specific conservation measures (as per Resolution 4/1996 in the Convention on the Conservation of European Wildlife and Natural Habitats), due to their environmental value (in particular landscaping and soil protection) and the difficulty of achieving successful natural regeneration. It is necessary to analyze the factors involved in its natural regeneration in order to advise forestry engineers and foresters.
Since the end of the 19th century, Spanish black pine stands in Spain have been managed under different systems that do not always promote forest stand persistence [2]. A shelterwood system with a 100 to 120 year rotation length and regeneration periods of 20 to 30 years regeneration period is now one of the most commonly used management methods [3,4]. Several authors agree that it is a shadetolerant species in the early stages, although others suggest that it may invade full-lighted sites. Preto [5] recommend a crown coverage level from 20 to 40% in order to achieve an adequate natural regeneration. Debazac [6] observed its capacity to establish both in abandoned crops and under 90 years old pure stands. Amorini and Gambi [7] concluded that Spanish black pine needs a high proportion of sunlight. Other authors focus the question from a side-shade point of view; thus Pardé [8] concluded that the natural regeneration is difficult in openings and requirements in side shade are needed at least. Recently, some research works has been focused on the Spanish black pine seed rain, seed germination and seedlings survival under different stand densities and soil preparation techniques [9,10]. Data from these studies showed that Spanish black pine regeneration can be avoided in different forest stand in Cuenca Mountain due to the low number of seedlings survival and that climatic conditions had a strong influence in regeneration process. However, an adequate interval of basal area in combination with soil treatments (i.e. scalping) can improve seed germination and seedlings survival in mast years (low seed production years).
Initial seedling recruitment has always been difficult to achieve due to factors including summer drought, soil compaction, masting conditions, seed predation, inadequate overstorey density, and attacks by European pine shoot moth on seedlings and damage from grazing animals, and the combination of each [3]. However, available information on management history shows that the challenges of natural regeneration of Spanish black pine stands in the Cuenca Mountains are not new. During the last century forest managers tried to enhance regeneration by using soil treatment, artificial regeneration, and the introduction of new varieties for the species. Yet, results have been inconsistent [9]. Protecting areas of natural regeneration from browsing by large mammals has also been necessary, and is a major problem for regeneration of forest stands.
The stages of seeds and seedlings (i.e. seed germination, seedlings survival and early seedling growth) have been recognized as the most important stages in natural regeneration governing the regeneration, structure and succession of vegetation [2]. Early natural regeneration can be viewed as the passing of a population through a series of environmental sieves that selectively filter out some individuals while allowing others to pass through to the next stage. Most Spanish black pine studies have been focused in seed germination or seedling survival stages [2,9-14] and little attention has been paid to the initial seedling growth. The objective of this study is to determinate in a general sense and under different field conditions (six experimental forests distributed across an altitudinal gradient) the influence of soil preparation on initial Spanish black pine seedling survival. The comparison among contrasting geographical ranges may contribute to an understanding of the role of environmental conditions in this natural regeneration stage and assist in forecasting Spanish black pine regeneration responses to global climate change. To accomplish this objective, initial seedlings growth were measured and modeller in different experimental sites and under two soil treatments. We hypothesized that soil preparation has a positive influence on initial seedling growth and dry weight although forest location can influence this trend.
From 2000 to 2002 six experimental sites throughout the Serranía of Cuenca (Spain) were set-up in even-age forest stand dominated by Spanish black pine, representing the climatic gradient of this area. The selected sites were Forest of Public Utility n° 131 “Cerro Candalar” (site 1), Forest of Public Utility nº 133 “Ensanche de las Majadas” (sites 2 and 3), where forest of Spanish black pine are mixed with Scots pine (Pinus sylvestris), Forest of Public Utility nº 109 “Ensanche de Buenache” (site 4), where Spanish black pine is mixed with Spanish juniper (Juniperus thurifera L.) and Forest of Public Utility nº 106 “Los Palancares y Agregados” (sites 5 and 6), close to pine stands where Spanish black pine is mixed with Maritime pine (Pinus pinaster). According to the soil classification included in Soil Survey Staff (1999), Calcic haploxerept and Lithic haploxeroll is present in the forest of Public Utility n° 106, Los Palancares y Agregados (sites 5 and 6 respectively) whereas Typic xerorthent and Lithic xerorthents can be found in the others experimental sites. Further information related to the experimental sites can be viewed in Tables 1-3.
| Site | Experimental site | n/ha | B.A. | Dm | Hm | Ho | Principal Stand Composition | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | “Cerro Candalar” | 399 | 12.59 | 25.54 | 12.41 | 13.56 | 100% Pn | |||
| 2 | “Ensanche de lasMajadas” | 265 | 11.79 | 24.19 | 13.12 | 14.89 | 75% Pn; 25% Ps | |||
| 3 | “Ensanche de lasMajadas” | 387 | 13.52 | 26.44 | 12.72 | 13.21 | 70% Pn; 30% Ps | |||
| 4 | “Ensanche de Buenache” | 373 | 13.56 | 29.71 | 15.85 | 16.45 | 70% Pn; 30% Jt | |||
| 5 | “Los Palancares y Agregados” | 307 | 11.87 | 23.16 | 11.33 | 13.01 | 100% Pn | |||
| 6 | “Los Palancares y Agregados” | 555 | 14.02 | 27.94 | 13.76 | 15.35 | 100% Pn | |||
n/ha: Number of trees per ha; B.A: Basal area (m2/ha); Dm: Average stand diametrer (cm); Hm: Average stand height (m); Ho: Dominant stand height (m).Pn: Pinus nigra Arn. ssp salzmannii,; Jt: Juniperus thurifera L.; Pp: Pinus pinaster ; Ps: Pinus sylvestris.
Table 1: General characteristics of the forest stand at each experimental sites.
| Site | Geographic coordinates | A | p | E | P | Ps | S | tmm | tm | Tmm |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 40º15'40'' N, 1º56'40'' W | 1350 | 2.0 | - | 1137 | 139 | 3 | -4.5 | 9.6 | 28.3 |
| 2 | 40º14'30'' N, 1º58'10'' W | 1440 | 19.5 | North | 1137 | 139 | 3 | -4.5 | 9.6 | 28.3 |
| 3 | 40º16'10'' N, 1º58'40'' W | 1420 | 10.0 | North | 1137 | 139 | 3 | -4.5 | 9.6 | 28.3 |
| 4 | 40º06'08'' N, 1º55'10'' W | 1335 | 3.8 | - | 1031 | 111 | 3 | -1.7 | 9.8 | 27.4 |
| 5 | 40º01'50'' N, 1º59'10'' W | 1230 | 4.0 | - | 600 | 100 | 3 | -0.5 | 11.9 | 30.5 |
| 6 | 40º01'20'' N, 1º58'40'' W | 1200 | 4.0 | - | 600 | 100 | 3 | -0.5 | 11.9 | 30.5 |
A: altitude above sea level (m). p: slope (%). E: exposure. P: mean annual precipitation (mm). Ps: mean summer precipitation (mm). S: drought period (months). tmm: mean lowest temperatures of the coldest month (°C). tm: mean annual temperature (°C). Tmm: mean highest temperatures of the hottest month (°C).
Table 2: Geographical location and physiographical and climatic characteristics of each experimental site (Meteorological station of Environment Ministery, Cuenca 1950-2007).
| Site | Soil Type* | O | A | B or C | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Th | Th | R.F. | C. | S. | Sa. | Th | R.F. | C. | S. | Sa. | ||
| 1 | TypicXerorthent | 3 | 21 | 15 | 22 | 10 | 68 | 39 | 17 | 24 | 24 | 52 |
| 2 | Lithic Xerorthents | 7 | 20 | 43 | 16 | 10 | 74 | 40 | 80 | 4 | 38 | 58 |
| 3 | Lithic Xerorthents | 5 | 25 | 35 | 20 | 8 | 72 | ** | 90 | 16 | 8 | 76 |
| 4 | TypicXerorthent | 6 | 44 | 48 | 16 | 12 | 72 | >20 | 10 | 22 | 12 | 66 |
| 5 | Calcic Haploxerept | 3 | 8 | 13 | 18 | 10 | 72 | 33 | 3 | 30 | 4 | 66 |
| 6 | Lithic Haploxeroll | 5 | 18 | 42 | 4 | 6 | 90 | 28 | 50 | 6 | 4 | 90 |
*Soil Type: Soil Survey Staff (1999). Tn: soil thickness (cm). RF: rock fragments (%). C: clay (%). S: silt (%). Sa: sand (%).**Deeper than soil pit because rock fragments
Table 3: Summary of edaphic characteristics for each experimental site.
On each experimental site, arranged together in two 150 m parallel strips (north-south orientation) 30 rectangular plots were established (10 m length x 3 m width) in early February of 2000. Each plot was influenced by a similar Spanish black pine canopy cover and overstorey density, which can be observed in Table 1. With respect to soil treatments, three types of seedbed were once made in early January of 2000 before starting dissemination and randomly assigned to each plot: brushing (removal of ground vegetation), scalping (organic matter removed from up to 1–2 cm of mineral soil) and control (without treatment). Brushing and scalping were made with the straight blade of a Caterpillar D7. Removed vegetation and ground vegetation of control plot was characterize by Eryngium campFestre L., Thymus bracteatus L. and Geranium selvaticum L., for experimental sites number 1, 2, 3 and 4 and by Eryngium campestre L., Geranium selvaticum L., Centaurea paniculata and Achillea odorata for experimental sites number 5 and 6. At the end of the experiment in December-2002, all surviving seedlings had their height, crow volume (height x width x breadth) and basal diameter measured. Then and in order to obtain the seedlings dry weight, all seedlings were collected and carried out to the laboratory, where seedlings were dried using an oven (85 degrees during 48 hours) and weighted.
Hypothesized differences in initial seedling growth among experimental sites (random effect) and soil treatments (fixed effect) were tested using analysis of variance (ANOVA). Each representative forest stand can be considered as spatially independent. The post-hoc test applied was Fisher´s least significant difference (LSD) method. A significance level of α=0.05 was adopted throughout, unless otherwise stated.
Two models have also been adjusted by multiple linear regression (one with measurements taken in the field and another in the laboratory) with a view to estimating the biomass (total dry weight) of the seedlings measured. The structure of the models studied is as follows:
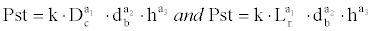
where Pst is the total dry weight (g), Dc is the diameter of the crown, Lr is the length of the whole root (mm), db is the diameter of the stem base (mm), h the total height of the aerial section (mm), and k, a1 a2 and a3 are the coefficients estimated by the LSM.. Statistical analyses were made with Statgraphics 5.0® and JMP 7.0® software.
Results showed that whereas experimental site was an important source of variation, soil preparation was not an important factor for any tested parameter related to initial seedling growth. As can be seen in Table 4, seedlings basal diameter and height were significantly influenced by experimental site but not by soil treatment. With respect to crow volume and dry weight, neither experimental site nor soil preparation were important factors (P>0.05).
| Factor | n | Basal diameter | Height | Crown volume | Dry weight |
|---|---|---|---|---|---|
| Experimental site | 182 | * | * | not significant | not significant |
| F-ratio | 2.49 | 3.03 | 1.13 | 2.23 | |
| P-value | 0.02 | 0.01 | 0.75 | 0.06 | |
| Soil preparation | 182 | not significant | not significant | not significant | not significant |
| F-ratio | 0.92 | 0.28 | 0.27 | 0.42 | |
| P-value | 0.49 | 0.75 | 0.34 | 0.65 |
Table 4: Results of analyses of variance for Spanish Black pine seedlings at the end of the experiment (December 2002). For selected factors (experimental site and soil preparation) asterisk indicates differences statistically significant(P-value<0.05).
Spanish black pine seedlings basal diameter tended to be greatest for the species located at Cerro Candalar (mean standard error: 0.96 ± 0.02 mm) and site 1 at Los Palancares y Agregados (mean standard error: 0.96 ± 0.01 mm). No differences were found for the rest of experimental sites (Ensanche de las Majadas site 1 and 2 (mean standard error): 0.87 ± 0.03 mm and 0.86 ± 0.03 mm, respectively; Ensanche de Buenache (mean standard error): 0.87 ± 0.02 mm and site 2 of Los Palancares y Agregados (mean standard error): 0.88 ± 0.03 mm). Soil preparation techniques did not have a significant impact in seedlings basal diameter although seedlings collected in soil preparation plots showed higher basal diameter (mean standard error: 0.93 ± 0.17 mm, 0.94 ± 0.15 mm and 0.84 ± 0.19 mm for scalping, brushing and control, respectively). In relation to seedling height, it was again highest in Cerro Candalar (mean standard error: 46.06 ± 1.03 mm). No differences were found between the rest of experimental areas (mean standard error: 39.01 ± 3.07 mm, 42.08 ± 1.90 mm, 43.24 ± 1.23 mm, 41.23 ± 1.67 mm, 38.72 ± 4.17 mm for Ensanche de Las Majadas site 1 and 2, Ensanche de Buenache and Los Palancares y Agregados site 1 and 2, respectively). Soil preparation generated higher seedlings height although differences were statistically not significant (mean standard error: 42.46 ± 2.73 mm, 42.47 ± 1.95 mm and 40.84 ± 2.09 mm for scalping, brushing and control, respectively). Crow volume factor was not influenced neither experimental site (mean standard error: 15.40 ± 3.17 mm, 14.40 ± 1.96 mm, 16.58 ± 2.24 mm, 17.96 ± 3.01 mm, 15.90 ± 2.67 mm, 16.41 ± 2.14 mm for Cerro Candalar, Ensanche de Las Majadas site 1 and 2, Ensanche de Buenache and Los Palancares y Agregados site 1 and 2, respectively) nor soil preparation (mean standard error: 15.75 ± 1.83 mm, 16.52 ± 2.78 mm and 16.13 ± 2.19 mm for scalping, brushing and control, respectively) (Figures 1 and 2).

Figure 1: Mean basal diameter, total height and crow volume for seedlings collected at each experimental site. CC: Cerro Candalar, EM1: Ensanche de las Majadas (zone 1), EM2: Ensanche de las Majadas (zone 2), Ensanche de Buenache, PA1: Los Palancares y Agregados (zone 1), PA2: Los Palancares y Agregados (zone 2).

Figure 2: Mean basal diameter, total height and crow volume for seedlings collected at each soil preparation
In the case of seedlings dry weight (Figures 3 and 4) forest location and soil preparation did not affect significantly this parameter (Table 4). However, seedling dry weight tended to be higher in Cerro Candalar (mean standard error: 0.28 ± 0.09 g) and in site 1 at Los Palancares y Agregados (mean standard error: 0.27 ± 0.08 g) if we compare with the other experimental sites (Ensanche de las Majadas site 1 and 2 (mean standard error): 0.19 ± 0.05 mm and 0.17 ± 0.05 mm, respectively; Ensanche de Buenache (mean standard error): 0.20 ± 0.06 mm and site 2 of Los Palancares y Agregados (mean standard error): 0.22 ± 0.03 mm). Seedlings dry weight was higher but not statistically significant in scalped and brushed plots (mean standard error: 0.23 ± 0.04 g and 0.24 ± 0.03 g, respectively) than in control plots (mean standard error: 0.20 ± 0.02 g)

Figure 3: Dry weight for seedlings collected at each experimental site. CC: Cerro Candalar, EM1: Ensanche de las Majadas (zone 1), EM2: Ensanche de las Majadas (zone 2), Ensanche de Buenache, PA1: Los Palancares y Agregados (zone 1), PA2: Los Palancares y Agregados (zone 2).

Figure 4: Dry weight for seedlings collected at each soil preparation.
The model which best represents the total dry weight data was the following:
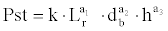
where Pst is the total dry weight (g), Lr is the total root length (mm), db the diameter at the stem base (mm), h is the total height of the aerial section (mm), and k, a1 a2 and a3 are the coefficients estimated by the LSM. The previous model has been adjusted after logarithmic data transformation as we go on to show:
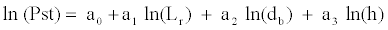
This model accounts for 56.56% of the dry weight variability according to the multiple correlation coefficient corrected in terms of explicative variables, of which Lr accounts for 23.85%, h for 20.05% and db• for 12.66%. The inflation of variance factor takes the values of 1.16, 1.19 and 1.25, respectively, thus no significant multicolinearility exists. According to the inclusion order in the model however, an explained variability of 27.31% was detected when comparing the variability explained by each variable collectively. Thus, the model that estimates the dry weights of those seedlings sampled in the laboratory would be as follows:
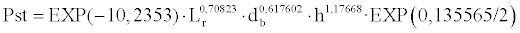
With regard to the seedlings measured in the field, which represent the real population, data collected from the aerial section variables were used exclusively. The model that best represents the total dry weight data and which exclusively takes these variables into account was:
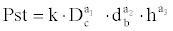
This model was adjusted by means of logarithmic transformation:
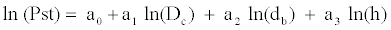
This model accounts for 44.39% of the total variability, of which the variables Dc, db and h account for 6.52, 16.83 and 21.04%, respectively. Although the inflation of variance factor is low (1.178, 1.204 and 1.256, respectively), 22.36% of collectively explained variability has been detected. The model to estimate the dry weights of the seedlings sampled in the field would be the following:
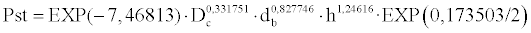
Management strategies must be focused on ecosystem persistence and a sufficient degree of ecosystem stability. A detailed understanding of the laws and processes, which determine ecosystem dynamics, is an essential basis for developing well-adjusted management plans. This understanding must include the natural regeneration process because a comprehension of natural regeneration is fundamental for planning the stand regeneration in order to get the stand persistence. The plant dynamics must be the base upon which any forest management decision must be made. Water deficits adversely affect several physiological processes, resulting in loss of vegetative growth as well as in injury and mortality seedlings. Moreover, our previous research [9-11] indicates that site preparation, which exposes the mineral soil, enhances conditions for Spanish black seed germination and seedlings survival. Light to moderate partial cut would also favour establishment because the seedbed conditions created would be less extreme than those encountered in more open, exposed seedbeds. Authors concluded on these previous research works that the effect of the overstorey density in combination with soil preparation can be summarised as being positive one on the Spanish black pine seed germination and seedlings survival in Mediterranean forest areas [9-11].
However and contrary to Spanish black pine seed germination and seedling survival percentages previously recorded in the same forest areas [11], soil preparation does not affect initial seedlings growth since differences between soil preparation techniques were always not significant (P>0.05). However, higher seedlings basal diameter, seedlings height and seedling dry weigh were always found in disturbed soils indicating that may be with longer period of study (more than one year) differences could be higher (statistically significant) and soil preparation could became an important factor in relation to initial seedling growth. Contrary to this, once Spanish black pine seedlings root can extend into the deeper soil to avoid competition from ground vegetation and to attain sufficient water and resources, herbal layer of control plots can have a beneficial component, generating lower soil temperatures and higher soil moisture [12]. Thus, soil preparation may negatively affect initial seedling growth. On this context, longer research period would be desirable and needed.
This work also showed that Spanish black pine initial seedlings growth tended to be higher at the most common and favourable habitats than in the marginal populations. As can be seen in Table 1, Spanish black pine populations dominated Cerro Candalar and Los Palancares y Agregados forest stand composition, indicating that these forest areas would be more appropriate and suitable for Spanish black pine initial seedling recruitment. Spanish black pine forests are mixed with Scots pine in Ensanche de las Majadas (sites 2 and 3) and with Spanish juniper in Ensanche de Buenache (site 4) indicating that some shifts in natural conditions occurs and others species and pines growth better, compete or even displace Spanish black pine. Thus, initial seedling growth appears to be site specific and further research would be desirable in order to obtain better conclusion on this topic.
In relation to seedling biomass models, the low proportion of variation explained by site and soil treatment with regard to seedlings biomass production, and the fact that root length is the best biomass predictor, suggests that biomass production at this age depends mostly on microsite conditions favorable for root development. Moreover, Good root development may allow maximum production at reduced fertility levels, and larger root systems may contribute to increased nutrient use efficiency [13], thus favoring initial seedling recruitment.
Higher overstorey density can promote higher seedlings height and basal diameter and negative effects of water deficit and drought period can be corrected (seedling protection by surroundings trees). On places where summer drought is not a problem with respect to initial seedling recruitment, higher sunlight reaching soil surface can be promoted with lower overstorey densities and thus, higher seedlings growth can be generated. At the present work, overstorey density effect over initial seedling growth was not tested (all experimental forest showed similar overstorey density). It would be desirable to evaluate it on the near future.
Knowledge of the processes and factors influencing nature regeneration is essential for its successful application. On this case, the optimal forest management plan should be a compromise between undisturbed forest floor for maximal early seedlings growth and soil preparation for promoting the number of germinated seeds and seedling survival (this last condition demonstrated by the author in other work developed in Cuenca Mountain). The nature of the relationship between natural regeneration of Spanish black pine and all factors involved appears to be site specific and further research work is needed to determine whether the relationships described above apply to other Spanish black pine stand across Spain. It is impossible to fully comprehend the dynamics of seedlings banks on the basis of short-tern studies. Therefore, long-term demographics studies on tree seedlings are necessary for understanding the processes of natural Spanish black pine regeneration, particularly on the new context of climatic change.