Pancreatic Disorders & Therapy
Open Access
ISSN: 2165-7092
ISSN: 2165-7092
Research Article - (2012) Volume 2, Issue 3
Abstract Objective: We evaluate the anti-tumor effect of Ethyl Pyruvate (EP) on the human pancreatic cancer xenograft in nude mice. Methods: Human pancreatic cancer cell line SW1990 was used in vivo to investigate the effect of EP on the human pancreatic cancer in mice. The mice were treated with different doses of EP (100 mg/kg, 50 mg/kg, i. p. ) initiating 1 hour (early treatment) after tumor cells injection daily for 14 days, and EP (100 mg/kg, i. p. ) was administered beginning12 days (delayed treatment) after tumor cells injection daily for 14 days. Tumor volumes were measured. ELISA was used to measure tumor necrosis factor (TNF) -α, interleukin (IL) -1β, IL-6 and high mobility group box 1 (HMGB1). Results: EP administration inhibited pancreatic tumor growth significantly. The differences in tumor volumes were statistical significant (P=0.0001) and in tumor weights (P=0.0028) between treatment and untreated groups. Treatment with the higher (100 mg/kg) dose of EP conferred better efficacy than lower (50 mg/kg) group (P=0.024). Early treatment with EP inhibited tumor significantly than delayed treatment (P=0.001). Early administration of EP inhibits TNF-α, IL-1β, IL-6 and HMGB1 release. Conclusions: EP may have potential as a therapeutic candidate for the treatment of pancreatic cancer.
<Pancreatic cancer is the fourth leading cause of cancer-related deaths in the world and has a poor prognosis; the 5-year survival rate of patients remains less than 5% and a median survival time after diagnosed is only 6 months [1,2]. Up till now, surgical resection still remains the only curative treatment for pancreatic cancer [3]. But, due to this aggressive disease, most pancreatic cancer patients have early metastasis by the time of diagnosis, and only a few cases can undergo surgical resection. Even among cases undergoing resection, the longterm outcome remains unsatisfactory because of early recurrence and metastasis [4]. Although significance advances have been made in the understanding the biology of pancreatic cancer, the molecular and cellular mechanisms involved in the metastasis and recurrence in the early stage or after resection remain poorly understood. Recent literature has highlighted an important role of inflammation in inducing and promoting cancer, chronic inflammation has been linked with the development and growth of cancer and even tumor metastasis [5-7].
Patients with ulcerative colitis, a chronic inflammatory disease of rectum and colon, have a 10-fold higher likelihood of developing colorectal cancer. Also, chronic hepatitis B or C and cirrhosis, an inflammatory condition of liver, increase the risk for the development of hepatocellular carcinoma [8]. The previous study has highlighted those innate immune cells, such as macrophages, NK cells, dendritic cells, granulocytes, drive cancer development through the production of proinflammatory cytokines such as Tumor Necrosis Factor (TNF), interleukin (IL) -1, IL-6 and High Mobility Group Box 1 (HMGB1), and these cytokines create inflammatory micro-environment of tumor growth and play an important role in inducing and promoting tumor development and metastasis [8-10]. Interactions of HMGB1 with Toll-Like Receptor (TLR) 2 and TLR 4, with recapitulation of signaling cascades originally activated by Lipopolysaccharide (LPS), could result in perpetuation of proinflammatory responses, and interventions inhibited intercellular signal associated with TLR would be effective in reducing the inflammatory actions of HMGB1 [11]. HMGB1, a nuclear DNA-binding protein, has been demonstrated to posse tumor promoting and tumor metastasis, angiogenenic activities and acting as a proinflammatory mediator [8,10]. Collectively, inhibiting HMGB1/ TLR pathways could be involved in cancer development and prolonged antitumor protection of chemotherapy and radiotherapy [5,6].
Ethyl Pyruvate (EP), a simple aliphatic ester derived from the endogenous metabolite pyruvate acid, has recently been reported to prevent lethality in mice with established lethal sepsis and systemic inflammation [12,13]. Other investigators reported that treatment with EP ameliorate organ injury or dysfunction in animal models of myocardial [14], intestinal [15], and severe acute pancreatitis [16]. EP inhibits activation of NF-kB in a variety of in vitro and in vivo systems and the release of inflammatory cytokines such as TNF-α, IL-1β, IL-6 and HMGB1 [12,16,17]. EP may suppress tumor development through its activity to induce cell death mode switch from tumor promoting necrotic cell death to tumor suppressive apoptotic cell death, and necrotic cell death is thought to contribute to tumor growth and aggressiveness through a tumor promoting cytokine HMGB1 [18]. Previously, our study demonstrated EP inhibited cell proliferation and induced cell apoptosis by inhibiting HMGB1 expression in pancreatic carcinoma cells [19]. Therefore, we reasoned that EP also might suppress growth of pancreatic tumor in vivo through inhibition of inflammatory cytokines release. Here, we investigated the effects of EP administration on pancreatic cancer development.
Cell lines and materials
Human pancreatic cancer cell line SW1990 was purchased from Beijing institute for Cancer Research (Beijing, China). Tumor cells were maintained in RPMI-1640 medium (Hyclone, USA) supplemented with 10% heat inactivated FBS (Gibco, USA) in a humidified incubator with an atmosphere of 5% CO2 and 95% air at 37°C. All chemicals were purchased from Sigma-Aldrich Chemical (Sigma-Aldrich, St. Louis, MO, USA). Ethyl pyruvate solution was prepared with 28 mM of EP, 130 mM of sodium, 4 mM of potassium, 2.7 mM of calcium, and 139 mM of chloride (pH 7.0).
Animals and tumor models
BALB/c- (nu/nu) nude mice, 4-5 weeks old, purchased from Beijing University Health Science Center (Beijing, China). All nude mice were bred and maintained in an air-conditioned animal house of Medical School of Shandong University under specific pathogenfree conditions, using an alternating 12-hour cycle of daylight and darkness; all nude mice were allowed free access to water and standard laboratory chow. This study protocol was conducted with the consent of the Committee for the Use of Experimental Animals of Shandong University School of Medicine.
After 1 week of adaptive feeding, BALB/c- nude mice were injected with 200 μl of suspension of 4×106 SW1990 cells into the left flank, randomly assigned to one of four groups, each consisting of five mice. Twelve days after injection, mice had borne tumors with sizes of 5×5 mm~6×8 mm, as measured by vernier caliper. Caliper measurements of tumors were recorded biweekly from this time point. Tumor volumes were estimated as length×width×width/2. Mice were anesthetized with injection of ketamine (100 mg/kg, i. p.) and xylazine (10 mg/kg, i. p.). At 33 days after injection, all experimental mice were sacrificed with their tumors removed and weighed to obtain Tumor Weight (TW). Antitumor activity was further evaluated by Tumor Growth Inhibition (TGI), calculated according to the formula: TGI= (1-TWt/TWc)×100% in treated (t) and control (c) mice (TW=tumor weight).
Experimental protocol
Firstly, we determine the effect of treatment with different doses of EP (100 mg/kg, 50 mg/kg, i.p.) initiating 1 hour (early treatment) after tumor cells injection daily for 14 days. Two groups of mice (n=5, each group) were used for this study. Secondly, we evaluate the therapeutic efficacy of EP when first treated in pancreatic carcinoma; treatment with EP (100 mg/kg, i.p.) was administered beginning 12 days (delayed treatment) after tumor cells injection daily for 14 days. Control mice were treated with the same vehicle solution. Thirdly, we observed the effects of EP treatment compared with control group. Tumor volumes were measured biweekly. The photographs of flanks with tumor were taken and tumors were removed and weighed at 33 days after tumor cells injection.
Cytokine levels
Blood was collected from the tail vein at 24 h after tumor cells injection, and ELISA was used to measure TNF-α, IL-1β, IL-6 and HMGB1 (Abcam, UK) levels. We also routinely measured serum levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), creatinine (Cr) using commercially available clinical assay kits before cell injection and after finishing treatment.
Statistical analysis
One-way analysis of variance with bonferroni-adjusted significance level was performed to compare the difference of means among groups for pre-test tumor size. Generalized estimating equations (GEE) model, with the distribution of dependent variable as normal, link function as identity, and within-group correlation structure as auto-regressive of order 1, was utilized to assess the effects among groups. Bonferroni-adjusted significance level was conducted for the pairwise comparisons of the effects among groups. All statistical analyses were performed with STATA version 12 (Stata Corporation, College Station, TX, USA). All reported probabilities (p-values) were two-sided, and less than 0.05 was considered statistically significant.
EP inhibits tumor growth in pancreatic cancer
To determine whether EP inhibits tumor growth in the pancreatic cancer, nude mice were treated with EP, including early treatment (administration EP initiated 1 hour after injection) and delayed treatment (12 days after injection). The results showed that EP administration inhibited pancreatic tumor growth significantly. The differences in tumor volumes between treatment and untreated groups were statistical significant (P=0.0001) (Figure 1, Table 1).Statistical differences were also observed in tumor weights between two groups (P=0.0028). Moreover, EP administration in every treated group also inhibited pancreatic tumor growth significantly, as compared with control group (early 50 mg/kg vs control, P=0.001; early 100 mg/ kg vs control, P<0.0001; delayed100 mg/kg vs control, P=0.017), as measured by pairwise comparisons (Table 2).
| Variable | β | SE | z | p | 95% CI | |
|---|---|---|---|---|---|---|
| pretest | 2.200 | .377 | 5.82 | 0.000 | 1.460 | 2.941 |
| time | -170.086 | 13.294 | -12.79 | 0.000 | -196.142 | -144.029 |
| group | ||||||
| 1 | -242.185 | 65.508 | -3.70 | 0.000 | -370.579 | -113.790 |
| 2 | -443.026 | 65.316 | -6.78 | 0.000 | -571.043 | -315.008 |
| 3 | -191.311 | 63.949 | -2.99 | 0.003 | -316.649 | -65.972 |
| Group time | ||||||
| 1 | 40.1874 | 18.801 | 2.14 | 0.033 | 3.337 | 77.036 |
| 2 | 64.60842 | 18.801 | 3.44 | 0.001 | 27.758 | 101.457 |
| 3 | 32.61127 | 18.801 | 1.73 | 0.083 | -4.238 | 69.460 |
| constant | 762.043 | 69.700 | 10.93 | 0.000 | 625.432 | 898.655 |
1 = early50 mg/kg; 2 = early100 mg/kg; 3 = delayed 100 mg/kg
Table 1: Analysis of tumor volume among groups by GEE model.
| Contrast | SE | Bonferroni | ||||
|---|---|---|---|---|---|---|
| z | p | 95% CI | ||||
| group | ||||||
| 1vs0 | -242.185 | 65.508 | -3.70 | 0.001 | -415.014 | -69.356 |
| 2vs0 | -443.026 | 65.316 | -6.78 | 0.000 | -615.347 | -270.705 |
| 3vs0 | -191.311 | 63.949 | -2.99 | 0.017 | -360.026 | -22.595 |
| 2vs1 | -200.841 | 69.687 | -2.88 | 0.024 | -384.693 | -16.988 |
| 3vs1 | 50.874 | 65.124 | 0.78 | 1.000 | -120.940 | 222.688 |
| 3vs2 | 251.715 | 65.727 | 3.83 | 0.001 | 78.310 | 425.120 |
1=early50 mg/kg; 2=early100 mg/kg; 3=delayed 100 mg/kg; 0=Control
Table 2: Pair wise comparisons of the tumor volume among groups.
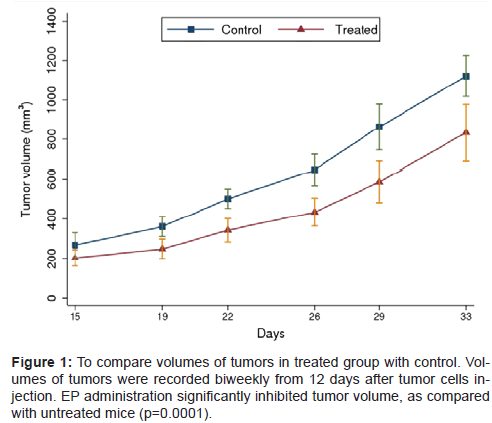
Figure 1: To compare volumes of tumors in treated group with control. Volumes of tumors were recorded biweekly from 12 days after tumor cells injection. EP administration significantly inhibited tumor volume, as compared with untreated mice (p=0.0001).
EP inhibits tumor growth in a dose-dependent manner
To assess the anti-tumor effects of different doses of EP, the mice received different doses of EP (100 mg/kg, 50 mg/kg) initiating 1 hour, daily for 14 days after injection cells. Treatment with the higher (100 mg/kg) dose of EP conferred better efficacy than lower (50 mg/kg) group. Significant difference in volumes of tumor was observed between two groups (P=0.024, Table 2, Figure 2A and 2B). The inhibition of tumor weight was boundary significance (P=0.076, Figure 2B and 2C). TGI was higher in the 100 mg/kg treatment group (32.82%) than in the 50 mg/kg treatment group (27.05%).Therefore, the anti-tumor efficacy of EP was dose-dependent fashion, higher dose of EP inhibited tumor growth significantly.
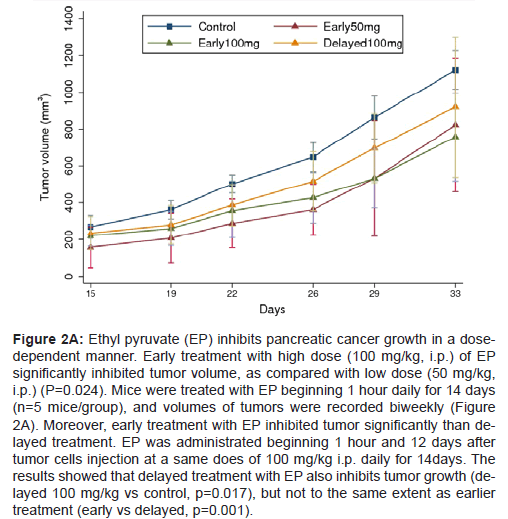
Figure 2A: Ethyl pyruvate (EP) inhibits pancreatic cancer growth in a dosedependent manner. Early treatment with high dose (100 mg/kg, i.p.) of EP significantly inhibited tumor volume, as compared with low dose (50 mg/kg, i.p.) (P=0.024). Mice were treated with EP beginning 1 hour daily for 14 days (n=5 mice/group), and volumes of tumors were recorded biweekly (Figure 2A). Moreover, early treatment with EP inhibited tumor significantly than delayed treatment. EP was administrated beginning 1 hour and 12 days after tumor cells injection at a same does of 100 mg/kg i.p. daily for 14days. The results showed that delayed treatment with EP also inhibits tumor growth (delayed 100 mg/kg vs control, p=0.017), but not to the same extent as earlier treatment (early vs delayed, p=0.001).
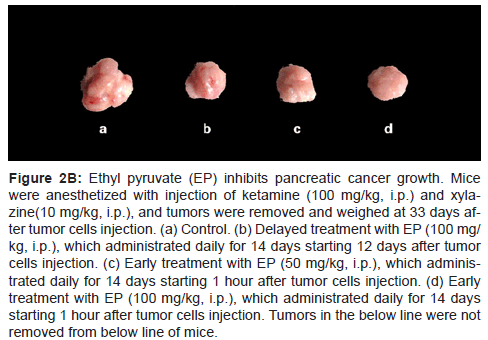
Figure 2B: Ethyl pyruvate (EP) inhibits pancreatic cancer growth. Mice were anesthetized with injection of ketamine (100 mg/kg, i.p.) and xylazine( 10 mg/kg, i.p.), and tumors were removed and weighed at 33 days after tumor cells injection. (a) Control. (b) Delayed treatment with EP (100 mg/ kg, i.p.), which administrated daily for 14 days starting 12 days after tumor cells injection. (c) Early treatment with EP (50 mg/kg, i.p.), which administrated daily for 14 days starting 1 hour after tumor cells injection. (d) Early treatment with EP (100 mg/kg, i.p.), which administrated daily for 14 days starting 1 hour after tumor cells injection. Tumors in the below line were not removed from below line of mice.
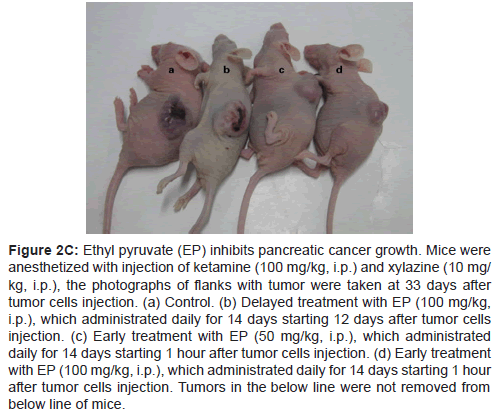
Figure 2C: Ethyl pyruvate (EP) inhibits pancreatic cancer growth. Mice were anesthetized with injection of ketamine (100 mg/kg, i.p.) and xylazine (10 mg/ kg, i.p.), the photographs of flanks with tumor were taken at 33 days after tumor cells injection. (a) Control. (b) Delayed treatment with EP (100 mg/kg, i.p.), which administrated daily for 14 days starting 12 days after tumor cells injection. (c) Early treatment with EP (50 mg/kg, i.p.), which administrated daily for 14 days starting 1 hour after tumor cells injection. (d) Early treatment with EP (100 mg/kg, i.p.), which administrated daily for 14 days starting 1 hour after tumor cells injection. Tumors in the below line were not removed from below line of mice.
Early treatment with EP inhibited tumor significantly than delayed treatment
To evaluate the anti-tumor efficacy of EP when first treated in mice, EP was administrated beginning 1 hour and 12 days after tumor cells injection daily for 14 days at a same does of 100 mg/kg. The results showed that delayed treatment with EP also inhibits tumor growth, but not to the same extent as earlier treatment. Statistical significance in the inhibition of tumor volumes (P=0.001, Table 2, Figure 2A) and boundary difference in tumor weights (P=0.066) between two groups (Figure 2B and 2C). TGI was higher in the early treatment group (32.82%) than in the 50 mg/kg treatment group (18.40%). These data suggested that suppression of early events important for tumor implantation and development is critical to maximize the tumor inhibitory effects of EP.
Early administration of EP cytokines release
The significant anti-tumor effects conveyed by EP suggested that it might target mediators and their receptors of systemic inflammation in promoting cancer. Treatment with EP beginning 1 hour after tumor cells injection significantly attenuated the systemic release of TNF-α, IL-1β, IL-6 and HMGB1 levels measured at 24 hours after cells injection (Table 3).
| Serum measurement | Early EP 50 mg/kg |
Early EP100 mg/kg |
Control |
|---|---|---|---|
| TNF-α (pg/mL) | 608 ± 53* | 388 ± 51*& | 930 ± 119 |
| IL-1β (pg/mL) | 127 ± 21* | 97 ± 15* | 261 ± 12 |
| IL-6 (pg/mL) | 75 ± 16* | 66 ± 11* | 158 ± 9 |
| HMGB1(ng/mL) | 48 ± 10# | 28 ± 6*& | 71 ± 9 |
*P<0.0001vs Control group; #P<0.01vs Control group; &P<0.05 vs earlyEP50 mg/kg
Table 3: Effect of EP on Serum Concentration of TNF-a, IL-1β,IL-6, and HMGB1 in mice with pancreatic cancer.
In this study, we used human pancreatic cancer cell line SW1990 vitro and BALB/c- (nu/nu) nude mice in vivo to study the effect EP administration on pancreatic cancer. Cell line SW-1990 derived from a human pancreatic ductal adenocarcinoma of a 56-year-old Caucasian male was established in tissue culture and the nude mouse. Cell line SW-1990 shows morphological, biological, and biochemical characteristics similar to that of the original tumor. Injection s. c. of 1×106 cultured neoplastic cells into nude mice resulted in tumor formation histologically closely resembling the original neoplasm. These characteristics make tumor line SW-1990 a valuable tool in studying various aspects of the biology of human pancreatic cancer [20]. Nude mice were suitable animal models that permit directing testing of human pancreatic cancer including cancer growth patterns and metastatic behavior, production of various biological markers, and response to treatment [20].
TNF-α, IL-1β, IL-6 are released early (within 6 h after LPS) in the development of a systemic inflammatory response and HMGB1 released as late cytokine (within 24 h after LPS) [12]. In this study, we designed to use early EP administration beginning 1 h and blood was collected from the tail vein at 24 h after tumor cells injection. The tumors can be measured 12 days after cells injection, delayed treatment were designed at this time point.
Pancreatic inflammation may play a key role in the development of pancreatic malignancy [21]. The response of pancreatic inflammation was mediated by cytokines, reactive oxygen species, and upregulated pro-inflammatory pathways [21,22]. EP treatment has been investigated in a variety of animal models of acute and chronic inflammatory disorders [12-17]. Our results in this study indicated that EP inhibited the growth of pancreatic cancer by reduction of release of early cytokines TNF-α, IL-1β, IL-6 and late proinflammatory HMGB1. The liver and renal toxicities of EP were not found between treated and control group. The functions of liver (ALT, AST) and renal (Cr) had no difference between control and treated group (data were not showed). That we used EP intraperitoneally maybe contribute to no or less toxicity of EP. One study has recently reported that EP has antitumor efficacy in the liver metastasis model and suggested that EP inhibition of tumor growth was mediated by tumor and host (decreased anti-inflammation) effects [22]. EP attenuates the inflammatory process significantly through inhibiting TNF-α, IL- 1, IL-6 and HMGB1 release as well as NF-kB activation [12,16]. It is estimated that infections and chronic inflammatory responses are involved in the pathogenesis of approximately 15% to 20% of human tumors [22-24]. Other causes of chronic inflammation, including mechanical, physical, and chemical injury as well as immune response to injury, also predispose to cancers [25,26]. Abnormal persistence of these molecules in chronic inflammation and in tumor microenvironments underlies carcinogenesis and tumor progression indicate that damage-associated molecular pattern (DAMP) molecule and their receptors could provide novel targets for cancer therapy [5,22,26]. HMGB1 was identified a “danger signal” from dying tumor cells that triggers this protective immune response through activation of TLR 4, and TLR 4 is associated with an increased frequency of metastasis in breast cancer patients after conventional chemotherapy [6].
In summary, these results reveal that EP may have therapeutic potential for pancreatic cancer and the mechanism of EP efficacy is effective inhibitor of early cytokines TNF-α, IL-1β, IL-6 and late proinflammatory HMGB1. EP, as a low-cost, anti-inflammatory agent may represent a novel therapeutic adjunct in the treatment for pancreatic cancer.
This work was supported in part by Natural Science Foundation of Shandong Province. No: ZR2009CM084