Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2014) Volume 3, Issue 3
Studies examined the effects of different culture conditions for the brain, gonads (testis and ovary), and liver of adult male and Female Fathead Minnow (Pimephales promelas) (FHM) during a 28 day assessment of viability and functionality. Five parameters were tested including media pH, incubation temperature, media composition, media exchange rate, and substrate. Within the ranges tested, incubation temperature was the most influential on tissue viability with dramatic improvements using cooler incubation temperatures (<18°C) over the entire 28 day test period. Tissues remained equally viable in five different common culture media, although highest viability with each medium was observed with a weekly media exchange over more frequent exchanges, alginate over polystyrene, and a media pH near physiological values. Use of these combined conditions resulted in continual function of testis and ovarian tissue over the entire 28 days, as indicated by continued production of 11-ketotestosterone (11-KT) by testes and estradiol (E2) by ovaries. Liver function was maintained through seven days, although vitellogenin production in response to added E2 eventually decreased over time. Consistent tissue viability over time periods commonly used for in vivo studies will enhance the link of in vitro tissue toxicology studies to whole-fish and population level impacts.
<Keywords: Fathead minnow; Endocrine tissue; Reproductive toxicology; Endocrine disruption; Tissue culture; Pimephales promelas
In vitro testing and its link to whole animal biology are becoming increasingly important in predictive ecotoxicology [1]. Better chemical screening systems are necessary to connect adverse outcomes of in vitro studies to population level effects through a mode of action [2]. In vitro culture of a key organ system of a model organism can serve as a potential screening tool if common conditions can be established for all tissues involved.
The hypothalamus (brain)-pituitary-gonadal axis and liver (HPG-L) is a complex organ system that is critical for sexual maturation and reproduction in all vertebrates including fish [3,4]. As a result, chemicals that disrupt the HPG-L endocrine system (endocrine disrupting compounds or EDCs) can have severe impacts on population survival of fish and other animals [5]. Due to the conservation of this system among vertebrates, extrapolating effects of EDCs on fish endocrine physiology can identify potential impact of these contaminants on humans when mechanisms are known. Since thousands of chemicals exist and many more are being introduced, there is an urgent need to determine the reproductive or endocrine disrupting potential of chemicals to which humans and the environment are exposed.
In vitro testing using cell lines and organ cultures offer significant advantages over the use of whole animals in EDC screening and toxicology such as better control of conditions, more defined exposures, greater spread in analysis and less use of animals for testing. While cell lines and assays are available for fish such as gill, muscle, and swim bladder, immortalized versions of all cell lines comprising the endocrine system, including the ovary, testis, pituitary, and brain, are not available, eliminating 2-D cell culture and the possibility of 3-D tissue replication at this time [6,7]. Additionally, cell culture systems lack endocrine or paracrine feedback responses that can compensate for low levels of EDCs as has been found in the fathead minnow, (Pimephales promelas), a common fish species used for ecological toxicity testing [8,9].
Because important tissues of the HPG-L are assemblages of more specialized cells, researchers have developed explant systems to assess the effects of EDCs on sex hormone production. Systems have been developed that utilize whole tissues cultured in vitro, including tail biopsies from Rana catesbeiana used for endocrine disruption detection [10] and testes culture from rainbow trout (Oncorhynchus mykiss) for reproductive studies [11]. Explants from human cervical, vaginal, and colorectal tissue have been successfully cultured for one to two weeks with applications for viral inhibition [12-15]. Testes [11,16] and ovary [17] explants have been successfully cultured for several fish species up to 36 days in vitro such that continued germ cell development and steroidogenesis occurred and viable offspring were generated from these cultures, indicating that long term tissue cultures are feasible and practical. Tissue explants from neonatal rats had superior histology and electrophysiologies and were more similar to freshly isolated tissue as compared to 2-D cell culture [18]. Ex vivo assays for the fathead minnow have been developed where ovary tissue slices are cultured for 14hrs and production of testosterone and estradiol (E2) monitored to detect chemical inhibition of any enzyme involved in the production of testosterone and E2 [19,20]. These studies demonstrate that it is possible to maintain explant cultures and that these cultures provide more realistic physiological systems in which to examine EDCs.
While the previous studies have demonstrated that tissue explants can be useful to assess the effects of EDCs, they fall short in examining the role of endocrine signaling in responding to EDCs. Assembly of tissue explants representing the components of the HPG-L axis into spatially isolated chambers that are fluidically connected on an engineered platform would be very valuable in examining effects on specific organ interactions and allow improved understanding of the impacts of EDCs on the entire system. In order to develop a HPG-L explant system, we explored how to create a common culture condition that maintains individual tissue viability and function such that hormone production is maintained. Here, fathead minnow tissue explants from organs involved in the HPG-L axis (ovary, testis, brain, and liver) were used to develop common culture conditions where all tissues maintained ≥ 80% viability after 28 days in culture. The 80% metric has previously been used as a standard of successful culture [7]. The effects of different media, temperature, and growth substrates on viability were examined. Once an optimal culture condition was established, the effects of culture time and signaling hormone exposure on estradiol (ovary), 11-ketotestosterone (testis) and vitellogenin (liver) production were examined. In addition to cell population viability, the location of living cells was determined using live- dead staining and imaging to examine necrosis along the tissue cross-section.
Animals
Juvenile fathead minnow were purchased from Aquatic Biosystems, Inc. (Fort Collins, CO) and reared in 20-gallon tanks. Fish were housed based on Environmental Protection Agency (EPA) guidelines established by Denny et al. [21] and followed an approved IACUC protocol for the University of Illinois. We replaced 4L of water on each of 6 days/week and 8L of water once weekly with conditioned and pH adjusted water. The fish were fed New Life Spectrum® hormone-free pellets twice daily. The tanks were located in a lab with an automated cycle of 16:8 h light and dark. We raised the fish to sexual maturity (5.5 to 6 months) before use in the studies. Fish chosen for study showed characteristics indicating sexual maturity, i.e., males displayed nuptial tubercles and a thickened fatpad on their heads and females had an enlarged abdomen and pointed face.
Dissections
Fathead minnow were anesthetized using 500 mg/L Finquel® MS-222 (Tricaine methane sulfonate; Ardent Chemical Laboratories, Redmond, WA) and 1000 mg/L sodium bicarbonate. The caudal artery was cut and fish were allowed to bleed out. We removed the liver, gonads, brain, and pituitary. Each organ, except the pituitary, was sliced several times using forceps and razor blades to allow for treatment comparisons using tissues from the same animal. Liver, brain, and testis slices were typically 1-5 mg, while ovarian slices were between 5 and 15 mg. The slices for each tissue were approximately equal in size for each treatment for each fish. Tissue weight was recorded to normalize data to tissue mass. The tissue slices were washed in chilled PBS+2% (by volume) penicillin-streptomycin, amphotericin B solution to reduce risk of contamination and kept in this solution until slices were weighed (<30 min). This step is important because, unlike traditional cell culture, bacteria and fungi may be present within the fish to contaminate the culture if left untreated. The slices then were placed in individual wells of 96-well plates assigned to specific conditions. The extracted pituitary typically weighed 0.02-0.15 mg and was too small to slice with razors while retaining representative tissue structure. Each pituitary was used for only one treatment, reducing sample size for this organ. Further, in pilot experiments, it was determined that pituitary viability had high variability when normalized using mass due to its small size. Therefore, the pituitaries were not included in these results, and based on anatomic proximity, an assumption can be made for future research that the brain culture preferences are reasonable starting conditions for pituitary culture.
MTT assay
Tissue viability was measured using the MTT assay [22]. MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide is a tetrazolium salt that is reduced to formazan in active mitochondria. Tissues were incubated in 10 μL MTT per 100 μL media for 4 hours. After 4 hours, the media-MTT supernatant was removed and replaced with 200 μL methanol to solubilize the formazan overnight. Tissue slices were removed after this solubilization process. UV absorbance of the resultant methanol solution was measured at a wavelength of 570 nm. Absorbance data were normalized using the mass of each tissue piece. A control slice from each organ was immediately assayed to acquire a baseline measurement at time zero of culture. Tissue viability over time was determined by comparing experimental normalized absorbance values to the control normalized absorbance value on tissue slices from the same organ.
Media formulation
Viability of tissues was evaluated in five common cell and tissue culture media types from Gibco®(Life Technologies, Grand Island, NY) that have been used previously in fish research [11,16,17,23-29]. Media used were Basal Medium Eagle (BME), Minimum Essential Medium (MEM), Leibovitz’s L-15 Medium (L-15), Medium 199 (Earle’s salts), and Medium 199 (Hanks salts). As-purchased culture media differed in salt combinations, buffering method, and presence of L-glutamine (Table 1). Media containing Hanks salts buffer well at atmospheric conditions, while Earle’s salts buffer better in CO2 controlled environments. The salts in Hanks and Earle’s salts typically buffer the media, but other buffering agents including sodium bicarbonate and HEPES are sometimes added to modify the buffering capacity. L-glutamine is an amino acid that can benefit cells in culture and is often included in media formulations. To reduce bacterial and fungal contamination, 0.5% gentamicin and 1% penicillin-streptomycinamphotericin B were added to all media. It was predicted that the tissues cultured in a medium with Hanks salts would improve the viability of the fathead minnow tissues compared to those made with Earle’s salts because the tissues were kept in atmospheric conditions in a cooling incubator.
| Medium | Earls salts | Hanks salts | L-Glutamine | Sodium bicarbonate | HEPES |
|---|---|---|---|---|---|
| BME | + | - | - | - | - |
| MEM | + | - | + | - | - |
| L-15 | - | + | + | - | - |
| Medium 199 | + | - | + | + | - |
| Medium 199 | - | + | + | - | + |
Table 1: Comparison of five culture media types for (A) seven day viability testsusing HPG-L tissues, n=28(B) 28 day viability tests using female HPG-L tissues, n=6. Error bars represent standard error of the mean.
Culture conditions
Tissue slices were placed in individual wells of 96-well culture plates for each experimental test. Each fish represented one replicate per treatment; slices from each organ were randomized by treatment, but each fish was represented in every treatment. Incubation temperature, pH, and frequency of media change were investigated. Incubation temperature was tested at 25°C, 20°C, 18°C and 4°C. pH was adjusted and maintained at five different values within the range 6.35-7.9 using either 1M HCl or 1M NaOH. We compared the viability of tissues when using daily, alternate day, and weekly media changes. All culture condition experiments included a 7-day culture period and a final 28-day culture period, unless otherwise noted. Optimal culture conditions and parameters (vide infra) were used for testing from this point on.
Culture substrate
Tissue explants are often cultured on substrates that are more suitable surfaces for cells than the polystyrene surface of culture plates, such as elder pith and gauze [7,16,25]. HPG-L tissues were cultured on polystyrene (PS) 96-well plate bottoms, Transwell™ (Millipore, Inc., Billerica, MA) polycarbonate membrane inserts, and 1.0% and 1.5% alginate coated wells for 7 days. Viability of each tissue type was compared using the four substrate types.
Pregnant mare serum gonadotropin (PMSG) and estradiol (E2) hormone dosing
Liver, ovaries, and testes were extracted, sliced, and cultured for 48 h using the optimal media parameters established in the culture experiments described above. This two day time period served to rinse any residual hormones from the tissues prior to exposure to endogenous hormones. Ovarian and testicular tissues then were cultured in 0, 2.5, 5, or 10 IU PMSG (Pro Spec, East Brunswick, NJ)/mL medium for 24 h. PMSG has been shown previously to stimulate hormone production in fish gonads [30]. Male and female liver were cultured for 24 h in 0, 250, 1000, or 2000 pg/mL E2. Media were collected from these tissues after this treatment period and an aliquot was analyzed for vitellogenin (Vtg) (from the liver tissue), estradiol (E2) (ovary), or 11-keto-testosterone (11-KT) (testis), that was produced as a result of the PMSG dose.
28-day hormone stimulation
Optimal culture conditions were used to culture liver, ovary, and testis tissues for 28 days. The medium was supplemented with 5IU/ mL PMSG for the ovary and testis cultures beginning at time zero and exchanged weekly. The medium for the liver cultures was modified by adding 208 pg/mL E2 (Cayman Chemical Co., Ann Arbor, MI) beginning at time zero and this medium was also exchanged weekly. Media supernatant was collected after 24 h and after each 7 d interval, replaced with fresh non-dosed L-15, stored and later analyzed for E2 in the ovarian media, 11-KT in testicular media, and Vtg in liver media.
Hormone immunoassays
Media were analyzed for E2 and 11-KT using Enzyme Immunoassay (EIA) kits and Vtg with an Enzyme-Linked Immunoassay (ELISA) kit (Cayman Chemical Co., Ann Arbor, MI). Assays were conducted according to manufacturer directions and we obtained acceptable values for intra- and interassay variations. The intra-assay variation for the E2 EIA was 7.2%, the inter-assay variation was 18.7%, and the limit of detection was 24 pg/mL. The intra-assay variation for the 11-KT EIA was 5.4%, the inter-assay variation was 14.4%, and the limit of detection was 0.27 pg/mL. The intra-assay variation of the Vtg ELISA was 7.7%, the inter-assay variation was 13.0%, and the limit of detection was 0.39 ng/mL.
Imaging
A set of tissue slices was examined using calcein AM and ethidium homodimer (Life Technologies, Grand Island, NY) live-dead stains. Tissues were incubated in 0.1 μL of 2 μM calcein AM and 0.4 μL of 4 μM ethidium homodimer in 200 μL phosphate buffered saline (PBS) for 2 hours. Images were taken of every 10 μm of tissue depth using an Olympus IX81 microscope (Olympus America Inc., Center Valley, PA) to determine if cells within the interior of the tissue remained viable throughout the 28 day period. We used a FITC 510 nm emission filter to image tissues stained with the calcein AM and a TRITC 610 nm emission filter for the tissues stained with the ethidium homodimer.
Statistical analysis
Statistical tests were run using Systat13® (Cranes Software International Ltd., 2008, San Jose, CA). Analysis of variance (ANOVA) was used to determine if there were differences in means of treatments in each experiment. Differences were considered statistically significant if p ≤ 0.05. Tukey’s test and Fisher’s Least Significant Difference (LSD) were used post hoc to determine which groups differed if a difference was found using ANOVA.
Values for hormonal immunoassays were normalized by the viability of tissues by dividing the measured hormone concentration by the % viability of the tissue at the time of the hormone sampling. The limit of detection was used for statistical purposes for any sample that was not detectable in these assays.
Fathead minnow (Pimephales promelas) is a common fish species used for ecological monitoring and toxicity testing due to its ease of use as a model species for reproduction and behavior (31,32). Our goal was to use tissue explants from all four organs involved in the HPG- L axis directly extracted from the fathead minnow as our organ models and then develop common culture conditions such that all tissues maintain ≥80% viability at 28 days in culture.
Media selection
Culture media type did not impact the viability for any of the tissue types after seven days of incubation (liver, F4, 128=1.418, p=0.232; ovary, F4,129=0.943, p=0.442; testis, F4,25=1.298, p=0.578; brain, F4,133=0.195, p=0.941) as shown in Figure 1a. Liver tissue viability exceeded 80% in all five media, but it appears to thrive in L-15 with a mean viability at least double that of the day-1 control. Brain tissues reached the 80% viability goal only in BME although the Confidence Interval (CI) included 80% for all media types except Medium 199 (Hanks).
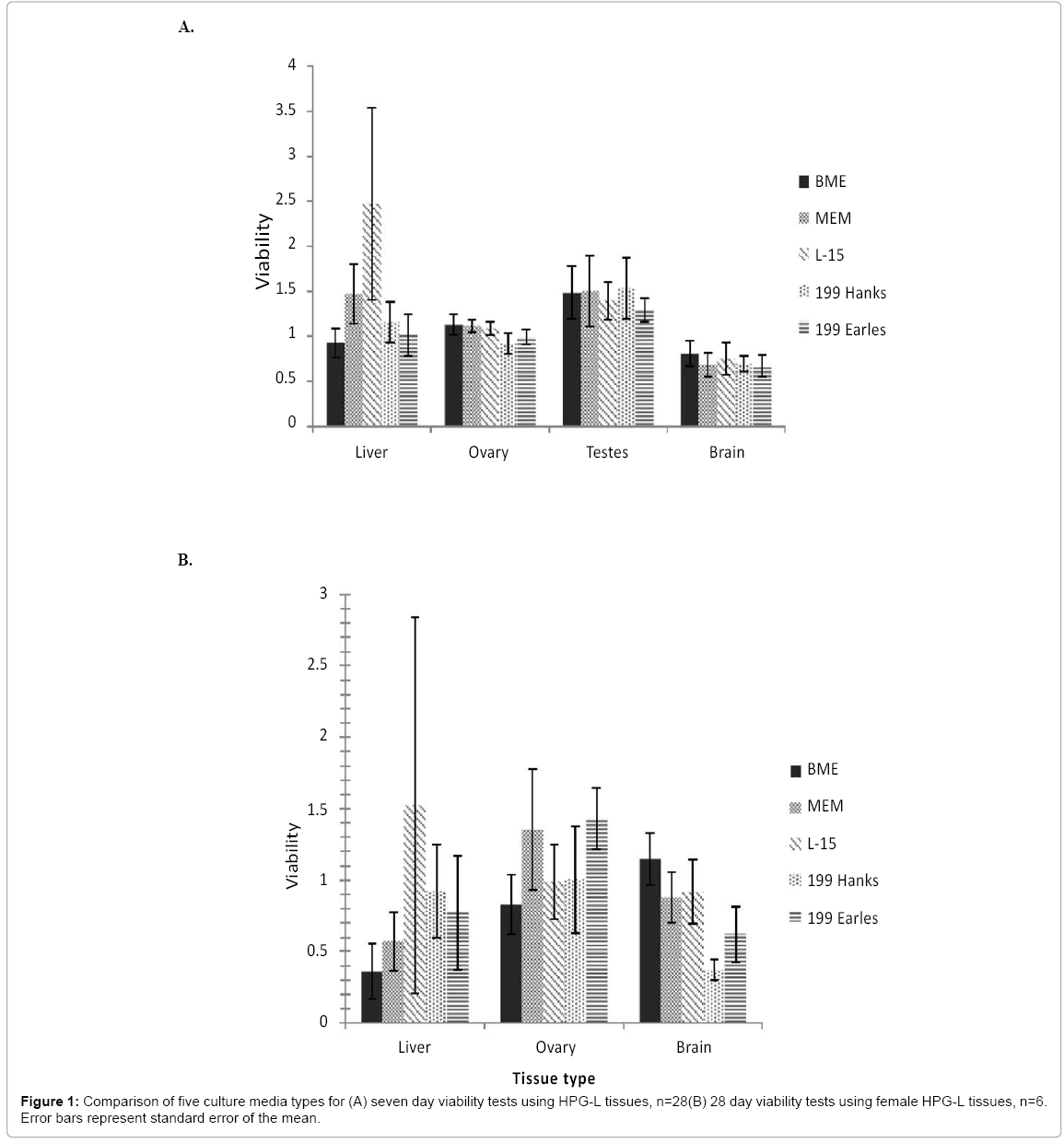
Figure 1: Comparison of five culture media types for (A) seven day viability tests using HPG-L tissues, n=28(B) 28 day viability tests using female HPG-L tissues, n=6. Error bars represent standard error of the mean.
A 28-day study was also performed using female fish tissues and did not result in a statistically significant difference for any media for any tissue (Figure 1b). A viability value of one indicates the tissue has the same viability, that is, the same metabolic conversion of MTT, after twenty-eight days in vitro as the control tissue did immediately after removal from the fish. A higher viability (>1) indicates a greater MTT assay value than the control tissue and more metabolically active cells are present in the sample, suggesting growth and/or replication of cells. All tissue types did maintain a mean of >80% in at least one media type when incubated for 28 days. The liver had a mean viability >80% in L-15 and Medium 199 (Hanks). The ovary had >80% viability in all media types. Brain viability was >80% in BME, MEM, and L-15.
pH
Media pH in this range had a notable effect on liver and ovary, and only a muted influence on testis and brain tissue. Liver tissue average viability ranged from 0.57 to 2.46, and the ovary average viability ranged from 0.64 to 2.25 (Figure 2). Testis and brain viability, however, went from 0.82 to 1.09 and 0.52 to 0.81, respectively. All four tissues show highest viability when incubated at pH 7.4, but this was significant only in the liver (F4,25=6.478, p=0.001). Liver viability was significantly higher at 7.4 than all other pHs (6.35, p=0.001), (6.74, p=0.007), (7.15, p=0.009), (7.9, p=0.006). Although viability was not significantly different in the other three tissues (ovary, F4, 25=2.065, p=0.116; testis, F4,25=0.376, p=0.545; brain, F4,25=0.557, p=0.696), the trend was consistent for all. The average viability was low at pH 6.35 for all tissues, increased as pH increased from 6.35 to 7.15, reached a peak at 7.4, then dropped again at pH 7.9.
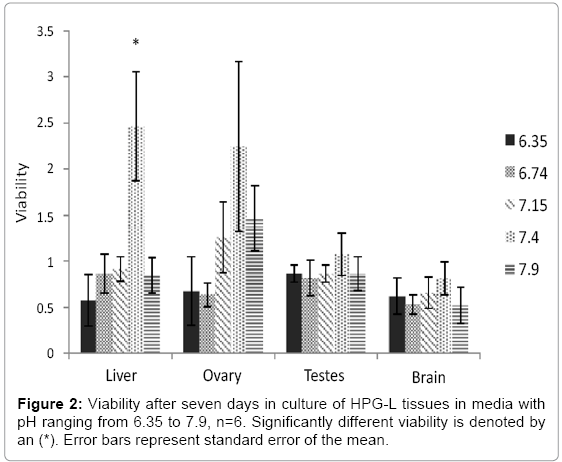
Figure 2: Viability after seven days in culture of HPG-L tissues in media with pH ranging from 6.35 to 7.9, n=6. Significantly different viability is denoted by an (*). Error bars represent standard error of the mean.
Temperature
Viability is clearly linked to incubation temperature (Figure 3). Of Viability is clearly linked to incubation temperature (Figure 3). Of the four tissue types, liver was the most sensitive tissue to incubation temperature (F4,32=4.897, p=0.003). Its highest viability was noted at the cooler temperatures 18 ºC followed by 4°C, and decreased significantly at 19.5°C and 25°C. Ovary and testis were the least sensitive to incubation temperature (ovary, F4,32=1.119, p=0.372; testis, F2,6=2.731, p=0.143). They maintained a viability >1 at all temperatures tested below 20 ºC. The brain followed a very similar trend as the liver, but appeared to be slightly less sensitive to temperature than the liver (F4,32=4.777, p=0.004). The brain had high viability (>0.8) at both 4°C and 18°C and then dropped significantly at 19.5°C (p=0.032) and again at 25°C (p=0.024). Since two of the tissues (liver and brain) had highest viability at 18°C, and the remaining two (testis and ovary) had close to the highest viability, 18°C was selected as the best common temperature condition.
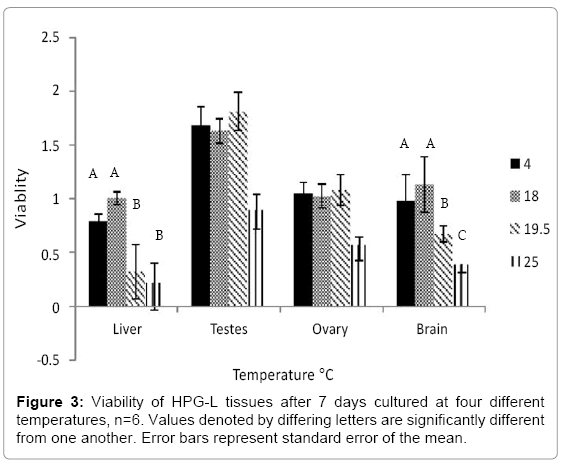
Figure 3: Viability of HPG-L tissues after 7 days cultured at four different temperatures, n=6. Values denoted by differing letters are significantly different from one another. Error bars represent standard error of the mean.
Media change frequency
Weekly media change and alternate day media change had no difference in viability compared to daily media change for liver, ovaries, and testes (liver, F2,10=0.663, p=0.537; ovary, F2,7=1.625, p=0.263; testes, F2,10=1.406, p=0.290). In a 28-day study, however, a weekly media change for the brain resulted in a statistically higher tissue viability than alternate day media changes (weekly mean=1.057, 2-day mean=0.354; F2, 10=5.613, p=.023, Tukey’s p=0.008).
Culture substrate
Tissues were cultured for seven days on four different substrates to determine the effects of materials in direct contact with the tissue, the media access to the tissue, and the ease of handling. Tissue slices were placed directly on the substrate, cultured for seven days under optimal conditions, and removed for the MTT viability assay. Ovaries began to grow into the Transwell™ membrane inserts in this time and were not removable for the MTT assay. Additionally, media changes were much less efficient using the Transwell™ inserts and exposed the tissues to nonoptimal conditions, such as increased temperature, longer than the other substrates. The inserts did not improve viability in tissues that were tested (Figure 4). For these reasons, the Transwell™ membrane inserts were not used for future experiments. The PS plate bottoms were easiest to use, as there is no additional procedure or handling. Both alginate surfaces require initial preparations to make the solutions and coat the plate bottom, but no additional handling time for the tissues is necessary.
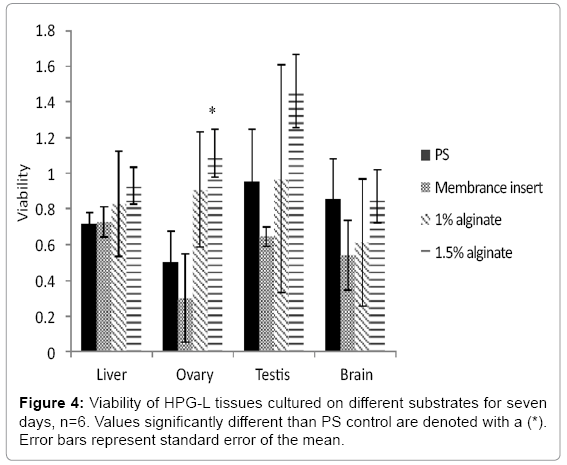
Figure 4: Viability of HPG-L tissues cultured on different substrates for seven days, n=6. Values significantly different than PS control are denoted with a (*). Error bars represent standard error of the mean.
The liver was the most robust tissue type in that the substrate material had little effect on viability. The brain tissue preferred the PS alone or the 1.5 % alginate. The liver and brain viability were not different between male and female fish and therefore data from both sexes were combined. Neither liver (F3, 10=0.034, p= 0.602) nor brain (F3, 10=0.707, p=0.569) viability changed with the use of the alginate substrate. Viability in the testes was the most variable but the differences were not statistically different (F3, 10=1.427, p=0.292). The data indicate a preference trend for 1.5% alginate for the testis. Use of 1.5% alginate as a substrate on the bottom surface of the wells also increased tissue viability in the ovaries (F3, 30=2.96 p=0.024).
Dose response of gonad tissue to PMSG
Gonads were stimulated with periodic doses of PMSG to stimulate production of sex steroids. Ovaries responded to increasing PMSG dose concentration with an increase of E2 production (Figure 5a; F3, 8=3.039, p=0.093). The average E2 produced in the 24 h exposure increased from a baseline production of 250 pg/mL to 700 pg/mL as PMSG increased from 0 to10 IU/mL. The in vitro ovary tissue continued to produce E2 over the course of 28 d (Figure 5b; black bars) although the production rate decreased over time. The ability of the ovary to respond to the endogenous PMSG is maintained over the entire 28 d time course as evidenced by the increased E2 production over baseline (Figure 5b; grey bars). Although testes did produce more 11- KT over basal levels when exposed to PMSG, it was not in a dose dependent manner over this concentration range (Figure 6a; F3, 8=0.436, p=0.733) after 24 h of exposure. Testicular tissue did increase 11-KT production over basal levels when stimulated by PMSG for 24 h on a weekly basis throughout 28 days (Figure 6b).
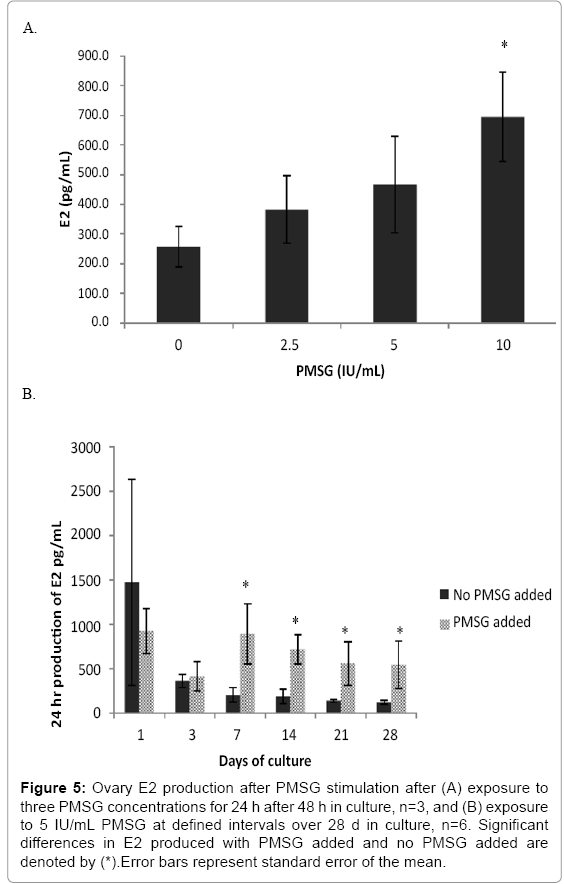
Figure 5: Ovary E2 production after PMSG stimulation after (A) exposure to three PMSG concentrations for 24 h after 48 h in culture, n=3, and (B) exposure to 5 IU/mL PMSG at defined intervals over 28 d in culture, n=6. Significant differences in E2 produced with PMSG added and no PMSG added are denoted by (*).Error bars represent standard error of the mean.
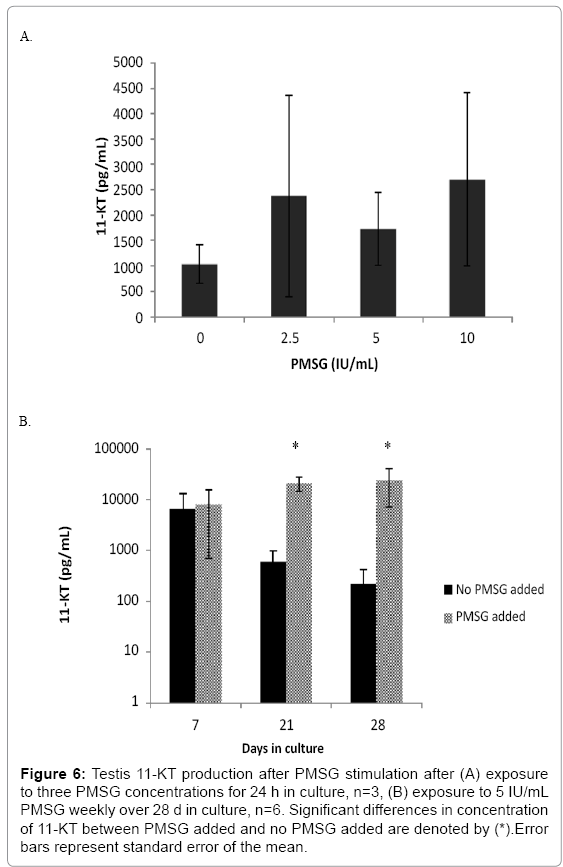
Figure 6: Testis 11-KT production after PMSG stimulation after (A) exposure to three PMSG concentrations for 24 h in culture, n=3, (B) exposure to 5 IU/mL PMSG weekly over 28 d in culture, n=6. Significant differences in concentration of 11-KT between PMSG added and no PMSG added are denoted by (*).Error bars represent standard error of the mean.
Dose response of liver tissue to E2
Female liver tissues cultured for 2 days responded to increasing doses of E2 from 0pg/mL to 2000 pg/mL by increasing Vtg production from 30.9 (±1.6) ng/mL to 10300 (±8.2) ng/mL (Figure 7; F3,8=34.785, p≤0.001). Male liver did not produce detectable amounts of Vtg (< 0.39 ng/ml) after 24 h of incubation without E2 stimulus in the 28 day study. It also did not increase production of Vtg in response to any E2 concentration in this range (Figure 7; F3,8=0.603, p=0.631) over 24 h after only 2 days in culture. However, after stimulation with 208pg/mL E2 for a longer 7 day time period, male livers produced low amounts of Vtg (38 ng/mL; Figure 8a). These levels, which were at least 100 times the basal level production, continued through day 28 and remained between 60-100% of the production seen on day 7. Female liver cultures produced very high amounts of Vtg during the 24 h relative to male livers (Figure 8b).
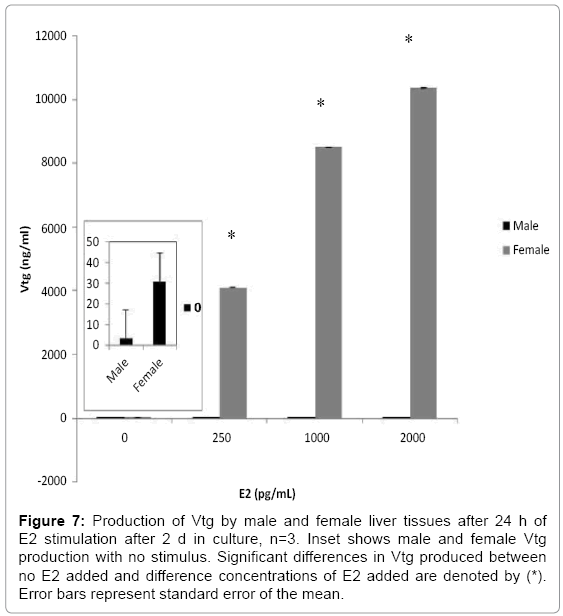
Figure 7: Production of Vtg by male and female liver tissues after 24 h of E2 stimulation after 2 d in culture, n=3. Inset shows male and female Vtg production with no stimulus. Significant differences in Vtg produced between no E2 added and difference concentrations of E2 added are denoted by (*). Error bars represent standard error of the mean.
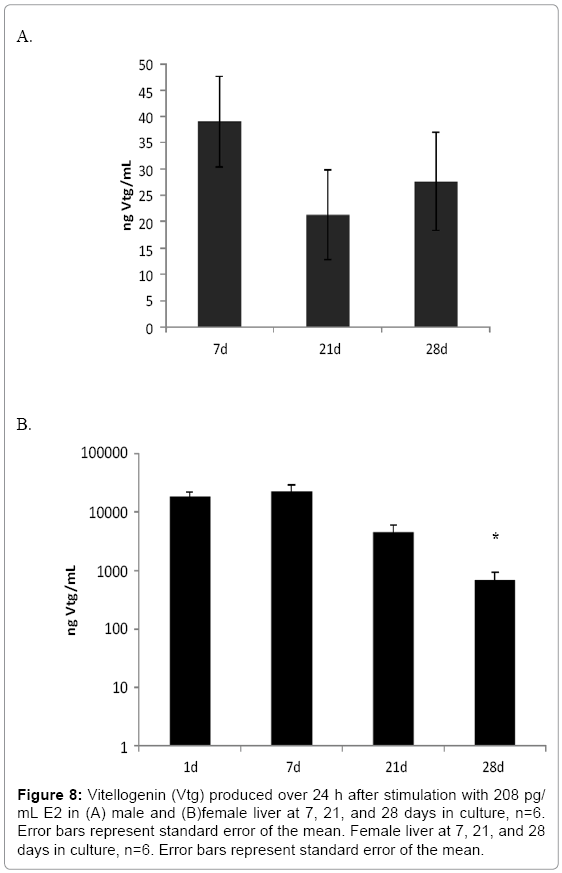
Figure 8: Vitellogenin (Vtg) produced over 24 h after stimulation with 208 pg/ mL E2 in (A) male and (B)female liver at 7, 21, and 28 days in culture, n=6. Error bars represent standard error of the mean. Female liver at 7, 21, and 28 days in culture, n=6. Error bars represent standard error of the mean.
Mean female liver Vtg production was 18,000ng/mL during the first 24 h without rinsing residual hormones from the tissue. The amount produced in a 24 h period in culture after rinsing increased to 22,000 ng/mL in response to 208 pg/mL E2 on day 7 demonstrating that female liver function is maintained in culture over this time period. By day 21, production of Vtg was only 34% of that on day 1 and by day 28 it was only 10% of day 1 production but still above that of the male.
Tissue integrity: gross appearance and viability staining
Brain, liver, ovaries, and testes maintained their 3D structure throughout the 28 d culture. Cellular outgrowth appeared at the edges of these tissues over time. The live-dead staining (Figure 9) indicates that living cells persisted throughout the entire tissue volume after the 28 d culture time and only punctate cell necrosis was observed in the tissue interior, rather than the presence of any extensive dead zones. At 14 d, most cells still alive. By 28 d, however, there was a mixture of live and dead cells throughout the tissues. While tissue samples were readily generated for imaging for the shorter time periods, we used all tissues at the longer time period (28 d) for MTT assays for statistical purposes since imaging was not our primary endpoint. Therefore, images for liver, ovarian and testis were not available at day 28.
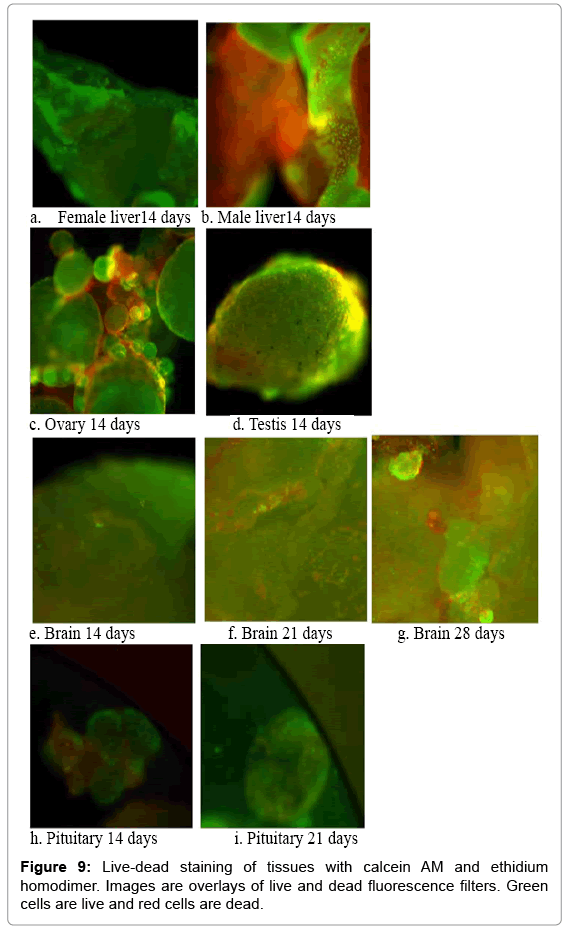
Figure 9: Live-dead staining of tissues with calcein AM and ethidium homodimer. Images are overlays of live and dead fluorescence filters. Green cells are live and red cells are dead.
Based on the results described above, the culture conditions that are suitable for each tissue type as well as the best conditions for all the tissues simultaneously are listed in Table 2. The ovary and testis responded similarly to all media types, maintaining a mean viability close to the control. In addition, the standard deviation in ovary viability was small indicating that this organ is extremely reproducible and robust. Further, this small standard deviation in the ovary serves as an indicator that our tissue handling procedures an unlikely cause of variation in viability for other tissue types. The brain was similar in that regard.
| Tissue | Media | pH | Temperature | Media change frequency | Substrate |
|---|---|---|---|---|---|
| Brain | BME, MEM, or L-15 | 7.4 | 4-18°C* | Weekly* | Equally viable |
| Liver | L-15 | 7.4* | 18°C* | Equally viable | Equally viable |
| Ovary | Inconclusive | 7.4 | <20°C | Equally viable | 1.5% alginate |
| Testis | Inconclusive | 7.4 | <20°C | Equally viable | Equally viable |
| Combined | BME, MEM, or L-15 | 7.4 | 18°C | Equally viable | 1.5% alginate |
Table 2: Optimized culture conditions for individual tissues and tissues combined in culture. * indicates statistical significance.
Results indicated that all media types can support tissue viability for 28 d and can be used for fathead minnow HPG-L co-culture. The five media selected for this research have been used successfully in previous single cell type and whole tissue culture for mammals and fish. Tissue viability was not statistically affected by culture media selection, indicating that researchers could use any of these media based on individual preference or availability. A viability ratio above 1 suggests that the media promotes tissue growth over the culture incubation period. In this regard, it appears that L-15 outperforms all other media for the liver cultures. However, it was not significant due to high variation. Although we increased sample size in the 28-day test to reduce the variation, the liver cells proliferated, resulting in some cultures that had as high as 300% viability above the controls after 28 days. Therefore, variability remained high despite a higher sample size than other tests. The viability of the ovary, testis and brain tissue was statistically equal in response to all types of media with no outliers in performance. The brain also appears to be the most delicate of the tissue types with none of the media promoting tissue growth over seven days as compared to the control since none of the viability ratios exceed 1. Our prediction that media containing Hanks salts would support tissues better than media containing Earle’s salts was not correct as there was no trend in tissue viability based on this component of the media despite the lack of CO2 control over the 28 days. L-15 is adequate to support all tissues simultaneously and is commonly used in fish tissue culture. It was therefore used as the base medium in the remainder of the experiments.
Tissue viability was most dependent upon incubation temperature and pH. Lower temperatures improved viability, which was expected due to reduced bacterial and fungal growth and lower cellular metabolism. All temperatures included in the study except 4°C were within normal fathead minnow spawning temperature range of 15°C to 30°C [33]. Incubation within the normal range would maintain relevant cellular function compared to low or high temperatures. Further, keeping tissues within the cooler end of the normal range appeared to reduce contamination and increased viability compared to higher temperatures in that range. Reinbold and Pescitelli [34] found that cold temperatures increase sensitivity to toxicants, so incubating tissue at the lower end the natural biological range would result in a tissue more responsive to EDC while remaining physiologically relevant. Additionally, 4°C did not improve tissue viability compared to 18°C. We were surprised that a mere 1.5°C temperature increase had such a great impact on brain and liver viability, whereas the gonads are quite resilient with this increase. We suspect that the reproductive tissues may be adapted to ambient temperatures and have lower blood flows, whereas core organs, particularly the brain, with high blood flow, may not be. Understanding the mechanisms behind the improved viability at even small decreases in temperature merits investigation but is beyond the scope of this study. All tissues thrived at 18°C and suffered reduced viability above that temperature. We recommend an incubation temperature of 15 to 18°C, at the lower end of the normal spawning temperature range.
Tissues showed a clear improvement in viability when incubated in media at pH 7.4 as compared to the other pH levels tested. This suggests that the common culture media should be buffered to 7.4 to maximize tissue viability. This result was not surprising here since the fathead minnow plasma immediately after dissection in this study measured 7.4. Although previous research that suggests higher pH benefits teleost tissue culture [35], the optimal culture pH may differ among teleost species. It should be noted, however, that the plasma pH for FHM reported here was not measured in a CO2 controlled atmosphere, and therefore is relevant to the pH conditions of the tissue cultures.
Brain was maintained as living tissue in BME, MEM, and L-15 after 28 days in culture. It was equally viable at all pH levels tested, but also trended toward increased viability at pH 7.4. The brain also demonstrated the best viability at 18°C.
The ovary and testis had viability ratios close to or greater than 1 for all media types at both 7 days and the ovary at 28 days, indicating that they are insensitive to these media types. The viability ratio is the result of the initial population of tissue cells, cellular replication, and cell death. A value greater than 1 is likely due to favorable conditions for tissue growth from replication. The gonads appeared to be the least affected by culture conditions, showing the most tolerance for nonphysiological pH and temperature. Therefore, it may be practical to use conditions optimized for liver and brain since ovary and testis appear to thrive in all media tested. The ovarian viability improved with an alginate coating on the well bottoms. This is not surprising due to the benefits of substrates that have similar structure to the extracellular matrix [11].
A weekly media change improved tissue viability during 28 d in culture compared to media change daily or every alternate day. This could be due to the reduced amount of handling and exposure to the ambient environment. Decreasing the frequency of media exchange from daily to weekly appeared to improve the health of the tissue over four weeks, probably due to decreasing the physical handling and disruption of the tissue. While media changes were performed under an aseptic hood, a chance for contamination exists each time the culture plates are opened. In addition, media change causes movement of tissues in culture, with the attendant risk of suction into or contact with the pipette tip, which may lead to damage and reduced viability. Our results are consistent with successful testes culture by Miura et al. [25], in which the media was also changed on a weekly basis.
Imaging techniques, including gross 3-D structure observations and live-dead staining, also provide evidence that these culture techniques maintain and promote physiological structure and viability. Ineffective culture conditions would result in flattened appearances of the tissues in the gross images, while live-dead images would have shown substantial dead areas especially in the interior regions of the tissues. Examination of the images did not illustrate either of these conditions. The alginate underlayer appeared to absorb the ethidium homodimer and at times, interfered with interpretation (e.g., the male liver, Figure 9b). The images otherwise showed predominantly green fluorescence indicative of living cells through 14 d. Imaging through the tissue depth via confocal microscopy showed that the center was not necrotic. After 28 d, however, fluorescence from red stained cells indicated that the amount of dead tissue was increasing over time.
PMSG and E2 were expected to stimulate healthy tissue so that the ovaries would produce E2, the testes would produce 11-KT, and the liver would produce Vtg. The production of E2, 11- KT, and Vtg in response to external endogenous hormone stimulation was an excellent indicator of not only viability but also functionality over time. Some decrease in production was predicted due to cell and tissue death but also due to the possibility that tissues preferentially replicate nonhormone producing cells. For instance, while Vtg production with E2 stimulation in the liver tended to increase at 7 d in culture in both males and females, the amount of Vtg produced from E2 stimulation decreased at later times. Vtg production at the 7 d culture time point was highly elevated for the female liver in response to E2, which could be due to the lack of a feedback loop when using only the liver. Both gonad tissues, however, retained the capacity for sex steroid production when exposed to PMSG throughout the entire 28 d culture period. This difference between continued liver and gonad hormone production can be explained with two possibilities.
The cell turnover in the liver may be higher than that in the gonads. If the newly produced cells are not Vtg-producing cells, then the viability of the tissue may remain high, while function decreases as the ratio of Vtg-producing cells to other cell types decreases. In the testis and ovary, however, if the cell turnover is lower, the ratio of hormoneproducing cells and other cell types remains more consistent over time. A second explanation is that the amount of E2 added to the liver samples in these tests (208pg/mL) was significantly less than the plasma concentration of E2 in females in vivo, previously reported as 6ng/mL [36]. Therefore, as culture time proceeded, any E2 that remained in the liver prior to tissue extraction was depleted. The dose-response of the liver to E2 over 24 h showed a response to E2 at concentrations of 1000 and 2000pg/mL. The reduced production of Vtg may then simply be due to non-physiological levels of E2 added.
Using the culture conditions that we arrived at in this study, viability was maintained for 28 days based on results from the MTT assay. Function continued throughout the 28 days as demonstrated by the ability to stimulate tissue to respond hormonally to exogenous hormones, although Vtg production decreased over time in female liver. This shows that gonads and liver maintained the ability to produce the hormones in vitro. Because the live-dead staining showed a mix of live and dead cells after 14 days and due to the decrease in Vtg production after 21 days, we currently consider this a robust culture system for studies lasting 14 days or less. Previous studies have cultured fish gonads for greater than 28 days and maintained function, so continued hormone production in culture is a realistic goal [11,16,25]. Based on our current results, this culture system is useful for in vitro toxicity culture studies in fathead minnow lasting up to two weeks and has the potential to be extended to even longer-term studies.
In this work, our goal was to define common culture conditions that satisfy the needs of all HPG-L tissue types simultaneously. This is a necessary step toward replicating the endocrine system in a fashion that allows an optimal and flexible study of individual organ components and combines brain, pituitary, gonad, and liver in an artificial, yet biofidelic responsive system. We have identified a set of co-culture parameters that serve to preserve and extend FHM tissue functionality for 14 days and viability for 28 days after excision from the fish. The critical parameters are the timing for the media exchanges, the culture temperature, and the pH, while the tissues are surprisingly indifferent to the media type. These culture parameters may even extend tissue longevity further, if needed. Established culture conditions for the HPG organs of the fathead minnow indicator species improves the ability of researchers to work with these tissues ex vivo to better understand the processes involved in endocrine disruption for each organ type. Use of in vitro tissue cultures allows careful reconstruction of the toxicity pathways with ultimate control over chemical and fluidic communications. Using tissue explants is simpler than having to rebuild the tissue architecture, while maintaining the variety of cell types included in each tissue. Researchers concerned with individual HPG-L tissues can determine optimal conditions for each tissue individually based on the data presented.
Tissue-based culture systems are emerging as platforms for toxicology research that improve upon both individual cell culture and whole organism studies through cost effectiveness, magnification of the system of interest, and increased flexibility. A tissue based endocrine system would use fewer live animals, have shorter test times, focus on the primary target of steroidogenesis, and form the basis for more specialized platforms for finer analysis of subtle effects as a tool for assessing the estrogenicity of environmental samples, chemicals and complex mixtures. Testing of this system is now underway using both endogenous and exogenous hormone additions and will be reported in a future manuscript. While additional parameters and conditions can be investigated and optimized in the future, such as glucose levels, O2 and CO2 concentrations, having a standard culture method allows us to compare mechanisms of action and endpoints more easily among numerous samples and temporally separated experiments. Discoveries of the impact of EDCs on HPG tissues will increase knowledge of the biological system function and lead to regulations and public information that can improve human and aquatic health.
This project was supported by funding from the U.S. Army Corps of Engineers, Basic Research Program. We would like to thank Drs. Jody Flaws and Megan Mahoney for their valuable technical support. Research was performed using Protocol # 12175 at the University of Illinois, Urbana-Champaign.